Šelmy
Šelmy (Carnivora) jsou řád placentálních savců, jehož zástupci jsou převážně predátoři živící se masem jiných obratlovců. I když jsou šelmy adaptovány k lovu a konzumaci masa, jejich jídelníček je často pestřejší a jenom málo druhů představuje striktní masožravce. Některé druhy se dokonce živí býložravě. Šelmy, které získávají maso aktivním lovem, využívají rozličných loveckých strategií a – v závislosti na druhu – zabíjejí širokou škálu živočichů. Predací šelmy přispívají k dynamické rovnováze ekosystémů, pomáhají snižovat množství kořisti (ať už kopytníků, nebo hlodavců), a udržují tak populace kořisti zdravější.
Stratigrafický výskyt: svrchní eocén – recent | |
|---|---|
 Typičtí zástupci šelem | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Podkmen | obratlovci (Vertebrata) |
| Třída | savci (Mammalia) |
| (nekategorizováno) | Carnivoramorpha |
| Řád | šelmy (Carnivora) Bowdich, 1821 |
| Čeledi | |
| |
| Sesterská skupina | |
| luskouni (Pholidota) | |
| Některá data mohou pocházet z datové položky. | |
Řád zahrnuje více než 270 žijících druhů, které spadají do 16 čeledí. Řád šelem byl tradičně dělen do dvou podřádů: suchozemské šelmy (Fissipedia) a ploutvonožci (Pinnipedia). Tento systém se však ukázal býti nepřirozeným, takže moderní systematika uznává spíše dělení šelem na podřády kočkotvární (Feliformia) a psotvární (Caniformia); ploutvonožci jsou v takovém případě zařazeni mezi psotvárné.
V rámci širší systematiky spadá řád šelem do širšího kladu Carnivoramorpha, který zahrnuje nejstarší známé předchůdce korunové skupiny šelem – viverravidy a miacidy. Tyto dvě skupiny se v období před asi 60 až 50 miliony lety, tj. během paleocénu a eocénu, rozšířily na kontinentech severní polokoule a vymizely ve svrchním eocénu před asi 37–34 miliony lety. Na konci eocénu se pak ve fosilních záznamech začínají objevovat první pozůstatky moderních čeledí šelem.
Šelmy jsou zřejmě nejrozmanitější řád mezi savci. Jejich nejmenší zástupce, lasice kolčava (Mustela nivalis), váží asi 50 g, zatímco rypouš sloní (Mirounga leonina) dosahuje hmotnosti až 5 000 kg. Lebka šelem se vyznačuje dobře vyvinutou zaoblenou mozkovnou a zkostnatělými bubínkovými výdutěmi, které obklopují sluchové kůstky středního ucha. Čelisti a zuby bývají přizpůsobeny zabíjení a konzumaci masa. Dolní čelist se pohybuje pouze ve svislé rovině, špičáky jsou adaptovány k zabití kořisti. Typickým znakem šelem je trhákový komplex, jenž sestává z modifikovaného posledního třenového zubu horní čelisti a první stoličky dolní čelisti. Ten jako nůžky pomáhá odkrajování kusů masa z mrtvých zvířat. Trháky ztratili všichni recentní ploutvonožci. Postkraniální skelet šelem je spíše zobecněn, ač i zde lze pozorovat některé společné znaky, jako je fúze některých zápěstních kůstek, oddělené kosti předloktí a holeně a chybějící, anebo pouze zbytková klíční kost. Pohyb je prstochodecký, anebo ploskochodecký, končetiny jsou vybaveny drápy, u některých skupin zatahovacími. Významné anatomické změny lze pozorovat u ploutvonožců, adaptovaných životu ve vodě. Smysly šelem jsou velmi dobře vyvinuty.
Šelmy osídlily téměř celý svět a jsou adaptovány prakticky všem biotopům. Lze u nich pozorovat rozmanité formy sociální struktury. Většina druhů žije samotářsky, ale některé mohou tvořit sociální skupiny. Rozmnožovací systém je většinou polygynní, kdy se samci páří s větším množstvím samic, monogamie je výrazně vzácnější. Mláďata suchozemských šelem se rodí slepá a s řídkým osrstěním, někteří ploutvonožci však produkují vysoce prekociální (vyvinutá) mláďata. Značně vyvinuté jsou u šelem komunikační dovednosti, častá je především komunikace na bázi čichových podnětů (moč, výkaly, aromatické sekrety).
V lidské kultuře mají šelmy dvojí roli. Na jedné straně vystupují jako vážená zvířata, někdy i božstva, pes nebo kočka byly domestikovány; na straně druhé jsou však konkurenty člověka. Šelmy byly a jsou pronásledovány, protože mají tendenci zabíjet lidmi chovanou drůbež a dobytek, zároveň jejich přirozené prostředí mizí. Další hrozbou pro populace divokých šelem jsou lov pro části jejich těl (spíše historicky i pro kožešinu), šíření nemocí, které jsou často přenášeny domestikovanými formami, izolované populace pak ohrožují faktory jako inbreeding nebo ztráta genetické rozmanitosti, např. vlivem genetického driftu. K roku 2018 bylo více než 70 druhů šelem nějakým způsobem ohroženo, čtyři druhy pak již zcela vyhynuly.
Systematika
Vnitřní systematika
Šelmy v rámci zoologického systému klasifikoval již švédský přírodovědec Carl Linné ve svém díle Systema naturae z roku 1758. Linné vytvořil skupinu Ferae, v rámci které sdružoval šest rodů jemu známých šelem: Phoca (tuleni), Canis (psi, vlci, kojoti, šakalové a hyeny), Felis (kočkovité šelmy včetně velkých kočkovitých šelem, jako lev, tygr či levhart, jež dnes spadají do rodu Panthera), Viverra (asijské druhy cibetek), Mustela (lasicovité šelmy) a Ursus (medvěd hnědý, rosomák sibiřský, jezevec lesní a mýval severní).[1][2] Označení Carnivora pro šelmy poprvé použil anglický cestovatel a přírodovědec Thomas Edward Bowdich roku 1821, přičemž toto pojmenování je od této doby platné.[3] Samotné pojmenování „Carnivora“ vychází z latinského „carō“ („maso“) a „vorāre“ („žrát“).[4][5]
Recentní šelmy byly tradičně děleny do dvou podřádů: suchozemské šelmy (Fissipedia), zahrnující všechny čistě pozemní druhy šelem, a ploutvonožci (Pinnipedia), zahrnující současné čeledi lachtanovití (Otaridae), mrožovití (Odobenidae) a tuleňovití (Phocidae). Rozdělení do těchto dvou podřádů zveřejnil již americký paleontolog George Gaylord Simpson ve velké taxonomické revizi savců zpracované v roce 1945.[6] Tento tradiční taxonomický systém se nicméně ukázal býti parafyletickým, tedy nepřirozeným; navzdory odlišnému způsobu života – a s tím spojenými anatomickými danostmi – analýzy krevního séra potvrdily blízkou příbuznost ploutvonožců s medvědovitými. Na základě těchto zjištění moderní systematika uznává spíše dělení šelem na podřády kočkotvární (Feliformia) a psotvární (Caniformia).[7] Kočkotvární demonstrují menší ekologickou a morfologickou různorodost ve srovnání s psotvárnými a při započítání fosilních taxonů psotvárné šelmy počtem rodů několikanásobně převyšují šelmy kočkotvárné.[8]
Ohledně vnitřní systematiky šelem stále nepanuje ve vědeckých kruzích jednoznačná shoda. Jedno z úskalí klasifikace představuje vysoká úroveň konvergence v rámci nepříbuzných skupin, kvůli níž je obtížné zařadit do systému především fosilní taxony, z jejichž pozůstatků nelze získat DNA, a tím vyřešit jejich postavení na fylogenetickém stromu pomocí biochemických analýz.[9]
Níže uvedený kladogram představuje vztahy mezi 16 uznávanými čeleděmi recentních kočkotvárných a psotvárných šelem.
| Vnitřní systematika recentních zástupců šelem na úrovni čeledí[9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
.jpg.webp)
.png.webp)
.jpg.webp)
Ploutvonožci jsou při uznávané klasifikaci na psotvárné/kočkotvárné zařazeni mezi psotvárné šelmy,[7][10] přičemž samotná skupina ploutvonožců, zastoupená třemi recentními čeleděmi, je považována za monofyletickou, a jako její nejbližší příbuzní jsou hodnoceni buďto medvědovití, anebo musteloidi.[11] Samotný taxon Pinnipedia bývá většinou označován za „nezařazenou taxonomickou jednotku“.[12]
Nadčeleď Musteloidea tvoří 4 recentní čeledi. Řadí se sem pandy malé (Ailuridae), jež tvoří samostatnou čeleď, a nejsou tak ani zástupci medvídkovitých (Procyonidae), ani zástupci medvědovitých (Ursidae), jako je panda velká.[13] V průběhu 20. století byly prakticky všechny menší psotvárné šelmy vyjma medvídkovitých, tj. lasice, kuny, medojedové, vydry, rosomáci, skunci a jezevci, zahrnuty v rámci několika podčeledí do čeledi lasicovití (Mustelidae). Teprve od konce 80. let 20. století vyšlo na základě molekulárních studií postupně najevo, že skunkovití – nyní samostatná čeleď – jsou vůči ostatním lasicovitým pravděpodobně vzdáleněji příbuzní než medvídkovití a zřejmě i pandy malé, i přestože se minimálně některými kraniálními charakteristikami podobají vydrám.[14][15]
Podobný „wastebasket taxon“ představovala i čeleď cibetkovití (Viverridae), do níž byly na základě morfologických charakteristik v minulosti řazeny i šelmy, které však podle molekulárních studií tvoří samostatné čeledi asijští linsangové (Prionodontidae), šelmy madagaskarské (Eupleridae) a nandinie (Nandiniidae).[16][17] Nandinie s jediným recentním druhem nandinie znamenaná (Nandinia binotata) jsou nyní považovány za bazální taxon vůči všem ostatním kočkotvárným.[13] Mezi cibetkovité bývali vyjma těchto skupin často řazeni i promykovití (Herpestidae), někdy od 70. let 20. století však začali být osamostatňováni a nyní je rovněž všeobecně přijímáno, že představují samostatnou čeleď.[16] Revidovaná čeleď cibetkovitých zahrnuje pouze podčeledi pucholové asijští (Hemigalinae), oviječi (Paradoxurinae), Genettinae (tj. africké ženetky + afričtí linsangové Poiana sp.) a cibetky (Viverrinae).[18]
Ještě problematičtější byla systematika madagaskarských šelem, tj. fosy, pucholů, fanaloky a galidií. Jejich taxonomie byla rozličně měněna, jednotlivé madagaskarské šelmy byly v průběhu 20. století řazeny mezi cibetkovité, jak bylo výše zmíněno, ale také do osamostatněné čeledi promykovitých, anebo do vlastních monotypických čeledí.[17] Molekulární studie z přelomu milénia nicméně prokázaly, že všichni tito zástupci jsou si navzájem blízce příbuzní a představují jednu monofyletickou linii, jež je nejblíže spřízněna s promykovitými; tvoří tedy jedinou samostatnou čeleď.[19][20][21]
Přehled recentních čeledí šelem
| řád šelmy (Carnivora) Bowdich, 1821 | |||
| podřád kočkotvární (Feliformia) Kretzoi, 1945 | |||
| Čeleď | Podčeledi | Výskyt | Příklad druhu |
|---|---|---|---|
| nandinie (Nandiniidae) Pocock, 1929 | jediný žijící druh obývá lesnatá stanoviště střední Afriky[22] |  nandinie znamenaná (Nandinia binotata) | |
| cibetkovití (Viverridae) Gray, 1821 |
|
tropická a subtropická stanoviště Eurasie, Středního východu a Afriky[22] |  cibetka africká (Civettictis civetta) |
| hyenovití (Hyaenidae) Gray, 1821 |
|
travnaté pláně, křovinná stanoviště, lesy a pouště Afriky, Indie a Blízkého východu[22] |  hyena skvrnitá (Crocuta crocuta) |
| šelmy madagaskarské (Eupleridae) Chenu, 1850 |
|
lesnatá stanoviště ostrova Madagaskar[22] |  fosa (Cryptoprocta ferox) |
| promykovití (Herpestidae) Bonaparte, 1845 | travnatá, savanová i lesní stanoviště Afriky, Asie a Středního východu[22] | -_is_it-_at_Hyderabad%252C_AP_W_106.jpg.webp) promyka mungo (Herpestes edwardsii) | |
| linsangové asijští (Prionodontidae) Horsfield, 1822 | zahrnují dva druhy žijící v jihovýchodní Asii |  linsang skvrnitý (Prionodon pardicolor) | |
| kočkovití (Felidae) Fischer de Waldheim, 1817 |
|
přirozený výskyt celosvětový vyjma Austrálie, Nového Zélandu, polárních oblastí, Madagaskaru, Japonska a většiny oceánských ostrovů[22] |  kočka divoká (Felis silvestris) |
| podřád psotvární (Caniformia) Kretzoi, 1938 | |||
| Čeleď | Podčeledi | Výskyt | Příklad druhu |
| psovití (Canidae) Fischer de Waldheim, 1817 | původní výskyt zahrnuje všechny kontinenty mimo Antarktidy, psovití se vyskytují v pouštích, pastvinách, savanách, lesích i v arktické oblasti[22] |  vlk (Canis lupus) | |
| medvědovití (Ursidae) Fischer de Waldheim, 1817 |
|
původní výskyt zahrnuje Severní Ameriku a Eurasii, Andy a pohoří Atlas od lesnatých stanovišť po arktickou oblast[22] |  medvěd hnědý (Ursus arctos) |
| mrožovití (Ursidae) Allen, 1880 | jediný žijící druh; cirkumpolární distribuce v mělkých arktických vodách[22] | .jpg.webp) mrož lední (Odobenus rosmarus) | |
| lachtanovití (Otariidae) Gray, 1825 | [P. 2] | subpolární, mírné nebo pobřežní vody Ameriky, Asie, jižní Austrálie, Nového Zélandu a různých oceánských ostrovů[22] |  lachtan kalifornský (Zalophus californianus) |
| tuleňovití (Phocidae) Gray, 1821 |
|
především polární, subpolární a mírné vody celého světa, vzácněji i tropické oblasti; některé druhy žijí i v jezerech a ústích řek[22] |
|
| skunkovití (Mephitidae) Bonaparte, 1845 | skunkové rodu Mydaus žijí v Indonésii a na Filipínách, ostatní druhy žijí na západní polokouli v Severní a Jižní Americe[22] | _DSC_0030.jpg.webp) skunk pruhovaný (Mephitis mephitis) | |
| pandy malé (Ailuridae) Gray, 1843 | vysokohorské bambusové lesy himálajské oblasti[22] |  panda červená (Ailurus fulgens) | |
| medvídkovití (Procyonidae) Gray, 1825 | mírné a tropické zalesněné oblasti Ameriky[22] | .jpg.webp) mýval severní (Procyon lotor) | |
| lasicovití (Mustelidae) Fischer de Waldheim, 1817 | šelmy pozemní, stromové i vodní, původně se vyskytují prakticky celosvětově s výjimkou Austrálie, Antarktidy, Madagaskaru, Sulawesi a některých dalších oceánských ostrovů[22] |  lasice kolčava (Mustela nivalis) | |
Modře zvýrazněna monofyletická skupina ploutvonožců. | |||

Postavení v rámci třídy savců
Šelmy (Carnivora) jsou řazeny do vývojové linie placentálních savců Laurasiatheria, její nadřazenou skupinou jest pak Boreoeutheria, sesterskou Euarchontoglires. Původ linie Laurasiatheria je pravděpodobně holarktický, vyjma šelem zahrnuje z recentních savčích řádů i hmyzožravce (Eulipotyphla), letouny (Chiroptera), lichokopytníky (Perissodactyla), monofyletické sudokopytníky (Artiodactyla) – tj. včetně kytovců – a luskouny (Pholidota).[26] Umístění těchto řádů na fylogenetickém stromu je předmětem dlouhodobých výzkumů a nejasností. Šelmy bývají konsensuálně většinou považovány za sesterský taxon luskounů, dohromady pak tvoří klad Ferae. Podle mnohých molekulárních studií Ferae společně se sudokopytníky (monofyletickými) a lichokopytníky náleží ke kladu Fereuungulata.[27] Jednu možnou klasifikaci představuje kladogram níže.[28][29]
| Laurasiatheria |
| ||||||||||||||||||||||||||||||
Pokud jsou do systematiky zařazeny i fosilní skupiny, korunová skupina šelem bývá řazena do širšího kladu Carnivoramorpha. Ten vyjma skupiny Carnivora (tj. Caniformia + Feliformia) zahrnuje parafyletickou čeleď Miacidae a čeleď Viverravidae.[30] Tyto dvě čeledi byly tradičně řazeny do nadčeledi Miacoidea,[31] nicméně novější studie se již snaží opouštět nepřirozenou systematiku, namísto toho zástupce miacidů + zástupce skupiny Carnivora hodnotí v rámci samostatného kladu Carnivoraformes, jenž je součástí kladu Carnivoramorpha. Viverravidae jsou vůči Carnivoraformes zřejmě sesterským taxonem, přičemž sami představují nejspíše skupinou monofyletickou.[32][33]
Prašelmy (Creodonta), tercierní skupina masožravých savců, pak mohou představovat sesterský taxon vůči Carnivoramorpha.[34] Podobně jako v případě šelem se i prašelmám vyvinuly trháky, nicméně v jejich případě se trháky nikdy neodvozují z premolárů, jako je tomu u pravých šelem.[35] Systematika kreodontů je obecně kontroverzní, není ani jasné, jestli je tato skupina monofyletická.[36]
Evoluce
Původ

V průběhu raného paleogénu se masožravost u placentálních savců objevila několikrát.[37] Největšími masožravci té doby byli zástupci skupiny Mesonychia, pocházející z Asie a následně kolonizující i Evropu a Severní Ameriku. Objevili se v paleocénu a vymizeli v průběhu oligocénu. Postavení těchto „archaických kopytníků“ v rámci skupiny Laurasiatheria je nejasné.[36][38][39]
V průběhu paleocénu se objevila také skupina kreodontů, kteří během paleogénu představovali nejpočetnější masožravé savce, přičemž někteří z nich se stali i hypermasožravci. V rámci kreodontů bývají tradičně hodnoceny dvě čeledi, Hyaenodontidae a Oxyaenidae (které však nemusí být sesterskými taxony), jež se vyskytovaly především v Severní Americe a Eurasii. Kreodonti jsou obecně považováni za blízké příbuzné pravých šelem, ale nejsou jejich přímými předchůdci.[36][38] Kreodonti dlouhou dobu koexistovali s pravými šelmami a nakonec jimi byli nahrazeni, přesné důvody vymizeni kreodontů jsou ale nejasné.[40] Je možné, že šelmy se svým méně specializovaným chrupem byly lépe adaptovány na změny přirozeného prostředí v důsledku klimatických změn a byly schopny konzumovat rozmanitější potravu.[7] Čeleď Oxyaenidae vyhynula ve svrchním eocénu, Hyaenodontidae přežívali až do miocénu.[41]
Nejstarší známí předchůdci korunové skupiny šelem bývají řazeni do dvou čeledí: Viverravidae a Miacidae.[42] Zástupci obou těchto čeledí se vyznačovali vesměs malou velikostí těla, dosahovali asi rozměrů dnešních lasicovitých. Viverravidé zřejmě žili na zemi a živili se hmyzem, miacidé představovali suchozemské a stromové predátory. Na základě tradičního pohledu byli viverravidé evoluční předchůdci kočkotvárných, zatímco miacidé evoluční předchůdci psotvárných, tento pohled je ale již překonaný. Viverravidé nejspíše představují bazální zástupce celého kladu Carnivoramorpha, miacidé – považováni nyní za parafyletickou skupinu – pak přechodné formy mezi viverravidy a korunovou skupinou šelem. Před asi 61–55 miliony lety se viverravidé a miacidé vyskytovali v Severní Americe a Asii, na začátku eocénu, před asi 55–49 miliony lety, pak pronikli i do Evropy. Ve svrchním eocénu, tedy asi před 37–34 miliony lety, kdy se ve fosilních záznamech začnou objevovat první pozůstatky moderních čeledí šelem, viverravidé i miacidé mizí.[32][43]
Kočkotvární

První kočkotvárné šelmy se ve fosilních záznamech objevují na konci eocénu a ve spodním oligocénu.[44] Za bazální kočkotvárné šelmy je pokládána čeleď Nimravidae, jejíž zástupci jsou někdy nazýváni též jako „nepravé šavlozubé kočky“. Nimravidé dosahovali velikosti od moderního rysa po moderního lva, a třebaže se anatomií lebky obecně podobali pravým kočkovitým šelmám, jejich sluchová oblast byla stavěna odlišně.[45][46] Podobnost s moderními kočkovitými šelmami je dána spíše konvergentním vývojem.[47] Alternativně jsou nimravidé považováni i za kmenové zástupce šelem nebo za kmenové psotvárné. Některé rody nimravidů, jako Dinictis, rozvinuly šavlovité špičáky, které se později objevily u některých zástupců kočkovitých. Nimravidé byli ve svrchním eocénu rozšířeni skrze Asii a Severní Ameriku, v průběhu oligocénu pronikli i do oblasti Evropy, vyhynuli až v průběhu svrchního oligocénu. Do čeledi nimravidů bývali řazeni i barbourofelidé, miocenní skupina kočkotvárných šelem, nicméně modernější studie určily jejich bližší spřízněnost spíše s pravými kočkovitými a nyní tvoří spíše samostatnou čeleď.[45][46] Barbourofelidé se vyvinuli ve Starém světě a později expandovali i do Ameriky. I v jejich případě lze pozorovat mohutné druhy se šavlovitými tesáky, přičemž vývoj čeledi vyvrcholil americkým druhem Barbourofelis fricki, jenž dosahoval kohoutkové výšky 90 cm a hmotnosti moderního lva. Barbourofelidé vyhynuli na konci miocénu, pravděpodobně v důsledku konkurence s modernějšími šelmami.[48][49]

V rámci kočkotvárných je nejrozmanitější skupinou – při započítání všech fosilních rodů – klad kočkovitých (Felidae).[50] Za nejstarší rod pravé kočkovité šelmy je pokládán Proailurus, jenž žil především na území Evropy od oligocénu až do spodního miocénu. Zvíře bylo velké asi jako ocelot, jeho lebka dosahovala velikosti asi 15 cm a ve srovnání s ostatními kočkovitými šelmami mělo vyšší počet zubů, zatímco modernější rody již rozvinuly odvozenější redukovaný chrup.[51][52] Před asi 20 miliony lety dal rod Proailurus – nebo některé příbuzné rody – vzniknout rodu Pseudaelurus, jehož zástupci jsou pokládáni za poslední společné předky moderních kočkovitých a machairodů (slavných „šavlozubých tygrů“, jejichž zástupci přežívali až do konce pleistocénu). Gracilnější druhy, jako P. transitorius a P. lorteti, se podobají recentním kočkovitým, zatímco P. quadridentatus vykazuje znaky machairodů.[pozn. 1][54][53] Ve svrchním miocénu kočkovití prodělali největší diverzifikaci a z Eurasie se postupně rozšířili i do Afriky a Severní Ameriky, kde rychle vytlačili původní nimravidy, resp. barbourofelidy.[50][52] Do období středního a svrchního miocénu také spadá období rozkolu malých a velkých kočkovitých šelem.[55] Ve svrchním pliocénu po vytvoření Panamské šíje pronikli kočkovití v rámci tzv. velké americké výměny i do Jižní Ameriky, čímž kolonizovali veškeré kontinenty s výjimkou Antarktidy a Austrálie.[50][52]
Nandinie (Nandiniidae), které bývají uváděny jakožto bazální taxon recentních kočkotvárných, nejsou známy z fosilních nálezů.[56]

Cibetkovití (Viverridae) se objevili v Eurasii v průběhu oligocénu před asi 25–35 miliony lety. Nejstarší rody řazené k cibetkovitým, jako je Semigenetta a Herpestides, byly datovány do období před asi 23 miliony lety. Asijští pucholové a oviječi se podle molekulárních dat odštěpili před asi 20–29 miliony lety. Do Afriky cibetkovití expandovali v průběhu miocénu.[57] Promykovití (Herpestidae) jsou rovněž čistě starosvětskou skupinou, nejstarší fosilní pozůstatky pocházejí z raného miocénu Afriky a Evropy. Ve svrchním miocénu promykovití pronikli i do Asie. Fosilní nálezy promykovitých jsou nicméně vcelku chudé. Blízce příbuzní promykovitým jsou madagaskarské šelmy (Eupleridae), jež pocházejí z afrického předka, který kolonizoval ostrov před asi 18–24 miliony let, tj. ve svrchním oligocénu až spodním miocénu. Fosilní nálezy madagaskarských šelem však zcela scházejí.[58][56]
Hyenovití (Hyenidae) se od posledního společného předka oddělili v období středního oligocénu před asi 29 miliony lety. Ve spodním miocénu se hyenovití vyskytovali v Evropě a Africe, a poměrně rychle, již v průběhu středního miocénu, kolonizovali i Asii. Během svrchního miocénu se u hyen objevila adaptace k drcení kostí, tj. vlastnost, jež je známá u většiny recentních druhů. Morfologie jiných zástupců hyen se vlivem konvergentního vývoje podobala morfologii psovitých šelem, přičemž v miocénu a pliocénu Eurasie a Afriky zaujímaly tyto druhy podobnou ekologickou niku jako psovití.[59][60] Zatímco kočkovití pronikli i do Severní Ameriky, hyenovití zůstali primárně starosvětskou skupinou. Jedinou známou hyenou, která překonala pevninský most v oblasti Beringovy úžiny, je plio-pleistocenní Chasmaporthetes.[52][61] Hyenky představují specializovanou hmyzožravou skupinu hyen s jedinečně adaptovaným chrupem. Doba jejich divergence od ostatních moderních druhů představuje otázku, původ skupiny je zhruba odhadován do období od středního do svrchního miocénu.[59]
Psotvární
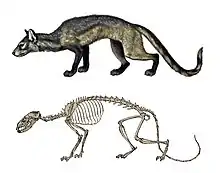
Čeleď psovitých (Canidae) se objevila v průběhu středního eocénu v Severní Americe, přičemž za její nejprimitivnější skupinu jsou pokládáni zástupci podčeledi Hesperocyoninae. Jejich nejranější formy (rod Hesperocyon, jenž se objevil před asi 37 miliony lety[62]) byly malé až středně velké šelmy představující evoluční mezičlánky mezi plantigrádními stromovými miacidními předky a pokročilejšími digitigrádními psovitými šelmami.[63][64] Později daly vzniknout předkům dalších skupin psovitých i pokročilejších linií hesperocyoninů.[62] Podčeleď Hesperocyoninae dosáhla největší diverzity ve svrchním oligocénu a ze Severní Ameriky vymizela během středního miocénu. Další velkou skupinou psovitých šelem byla pokročilejší podčeleď Borophaginae, rovněž endemitní pro Severní Ameriku, jež se objevila ve spodním oligocénu, největší rozmanitosti dosáhla během miocénu a přežívala až do pliocénu. Podčeleď Caninae, jež zahrnuje všechny recentní rody šelem, se objevila v Severní Americe ve spodním miocénu a následně pronikla i do Evropy, čímž představuje jedinou podčeleď psovitých, která opustila severoamerický kontinent. Rozmanitost podčeledi Caninae byla nicméně nízká až do první větší diverzifikace na konci miocénu, jež nastala pravděpodobně v důsledku úbytku hyenovitých v Evropě a zástupců podčeledi Borophaginae v Americe. Druhá velká vlna diverzifikace proběhla v průběhu pliocénu, kdy zástupci podčeledi Caninae kolonizovali Afriku a Asii, po vytvoření panamského mostu v plio-pleistocénu pak i Jižní Ameriku.[63][64][65]

Za nejstaršího zástupce čeledi medvědovitých (Ursidae) je považován severoamerický rod Parictis, jenž se poprvé objevil ve svrchním eocénu Severní Ameriky. Šlo o vcelku drobné zvíře, jeho lebka dosahovala délky pouhých 7 cm a celková hmotnost činila asi 2 kg. Rod Parictis bývá řazen společně s příbuznými rody do podčeledi Amphicynodontinae, jejíž zástupci přežívali až do spodního oligocénu, přičemž vyjma Severní Ameriky žili i v oblasti Eurasie (jako rod Amphicynodon z francouzských nalezišť). Postavení této skupiny – zřejmě parafyletické – v rámci psotvárných šelem je nicméně kontroverzní; samotný rod Parictis bývá považován i za kmenového ploutvonožce. Při vynechání podčeledi Amphicynodontinae prodělali medvědovití větší diverzifikaci až v průběhu miocénu. Úspěšnou skupinou byla podčeleď Hemicyoninae, jejíž nejstarší zástupci (eurasijský rod Cephalogale) žili již v oligocénu. Vyvinuli se z nich větší predátoři, nejspíš zdatní běžci, s dobře vyvinutými trháky. Během miocénu pronikli i do Severní Ameriky.[65][64][66] Od předků podobných rodu Cephalogale se pravděpodobně vydělili medvědi rodu Ursavus. Tento asijský rod je považován za předka všech moderních skupin medvědovitých. V rámci korunové skupiny medvědovitých se jako první odštěpily pandy (Ailuropodinae), podle dat z molekulárních hodin během miocénu (resp. před asi 12 miliony lety). Americká podčeleď Tremarctinae se vydělila před přibližně 6 miliony lety. Její zástupci kolonizovali i Jižní Ameriku, kde v současnosti přežívá poslední žijící druh, medvěd brýlatý. Ostatní skupiny moderních medvědovitých prodělaly radiaci během přelomu miocénu a pliocénu.[66][67]
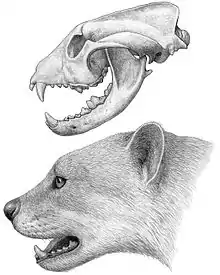
Problematickou skupinou psotvárných je čeleď Amphicyonidae,[pozn. 2] jež se objevila během středního eocénu v Severní Americe, později expandovala i do Starého světa. Šlo o úspěšnou tercierní skupinu, přičemž její pozdní zástupci dosahovali hmotnosti i 200 kg. Svou morfologií se tyto šelmy podobají jak psovitým, tak medvědovitým šelmám, některé studie je zařadily i jako bazální psotvárné.[64]
Skunkovití (Mephitidae) jsou známí ze spodního miocénu Evropy a svrchního miocénu Severní Ameriky. Do Jižní Ameriky, na rozdíl od medvídkovitých, pronikli jako většina šelem až během velké americké výměny. Fosilní skunkovitý rod Promephitis žil i na asijském kontinentu.[69]
Nejranějším rodem čeledi pandy malé (Ailuridae) byl Amphictis. Šlo o čistě evropský rod, jehož poslední zástupci přežívali ještě ve středním miocénu na jihovýchodě kontinentu. V průběhu miocénu se objevila podčeleď Simocyoninae, jejíž původ je asi rovněž evropský, nicméně zástupci simocyoninů kolonizovali i Asii a Severní Ameriku. Jedním ze zástupců skupiny byl masožravý rod Simocyon, dosahující velikosti pumy. Celá podčeleď Simocyoninae vyhynula ve svrchním miocénu, poslední zástupci možná přežívali až do spodního pliocénu Asie. Podčeleď Ailurinae je ve fosilních záznamech poprvé zaznamenána ve středním miocénu Evropy, možná i Číny. Během suchého svrchního miocénu se pandy malé pravděpodobně stáhly do vlhkých oblastí jižní Asie, a teprve během spodního pliocénu se rozšířily zpět do Evropy, a dokonce i do Severní Ameriky. Od svrchního pliocénu pandy malé opět přežívají pouze v Asii, přičemž moderní rod Ailurus je poprvé znám ze středního pleistocénu Číny.[70][71][72]
Původ lasicovitých (Mustelidae) je kladen rovněž do Eurasie, jejich nejstarší předkové se objevují ve svrchním eocénu, přičemž již ve spodním oligocénu jsou lasicovití zastoupeni širokou škálou fosilních taxonů. Ve svrchním oligocénu či spodním miocénu lasicovití kolonizovali i Severní Ameriku a Afriku, zatímco do Jižní Ameriky pronikli až s vytvořením Panamské šíje. Korunová skupina lasicovitých se objevila ve středním miocénu.[73][74] Medvídkovití (Procyonidae), nejbližší příbuzní lasicovitých, se poprvé nesporně[pozn. 3] objevují v průběhu raného miocénu v Evropě, krátce poté i v Severní Americe. Medvídkovití se stali jednou z mála skupin, která kolonizovala Jižní Ameriku ještě před velkou americkou výměnou, a sice ve svrchním miocénu. Od konce miocénu pak představují čistě novosvětskou skupinu šelem.[73]
Specializovanou skupinu v rámci psotvárných šelem představují ploutvonožci, kteří se pravděpodobně vyvinuli v průběhu svrchního oligocénu a raného miocénu, a to zřejmě v oblasti dnešní Arktidy.[75] Významnou přechodnou fosilií představuje Puijila z oblasti kanadské Arktidy, jež povrchně připomíná vydru. Puijila ještě nepřeměnila končetiny v ploutve, nohy však byly vybaveny plovacími blánami. Lebka vykazuje některé znaky shodné s ploutvonožci. Významným rodem jedněch z nejranějších ploutvonožců je Enaliarctos ze svrchního oligocénu a spodního miocénu západního pobřeží Severní Ameriky a Asie. Enaliarctos měl ještě stále výrazné trháky, ploutve již byly celkem dobře vyvinuty.[76][77] Původ kladu zahrnující současné čeledi lachtanovití (Otariidae) a mrožovití (Otariidae) je kladen do severního Tichomoří.[75] Nejstarší mrožovití jsou datováni do středního miocénu, a třebaže dnes žije pouze jediný druh mrože, historicky šlo o druhově a rodově bohatší skupinu. První jednoznačná lachtanovitá šelma pochází z pozdního miocénu Kalifornie.[76] Tuleňovití (Phocidae) se ve srovnání s lachtanovitými a mrožovitými pravděpodobně vyvíjeli v jihovýchodním Atlantiku a teprve později pronikli do chladných polárních vod severní a jižní polokoule.[75] Do období před středním miocénem, tj. před asi 16 miliony lety, je datován rozkol dvou hlavních tuleňovitých podčeledí tuleni šedí (Monachinae; primárně jižní polokoule) a tuleni praví (Phocinae; především studené severní vody).[78] Ze středního miocénu Evropy a Severní Ameriky pak pocházejí první fosilní nálezy tuleňovitých.[76]
Anatomie
Hlava
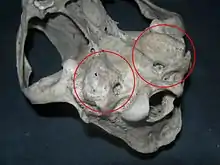
Lebka většiny šelem je adaptována na masožravý způsob života. Mozkovna je dobře vyvinutá a zaoblená, díky níž je korunový šev spojující čelní kost s temenními kostmi umístěn vpředu.[79] Mozek je v poměru k velikosti těla velký, dobře gyrifikovaný,[80] přičemž větší relativní velikost mozku mívají především masožravé nebo všežravé šelmy.[81] Mezi klíčový determinační znak, na základě něhož lze rozlišit kočkotvárné a psotvárné šelmy, patří anatomie bubínkové výdutě, což je kostěnné pouzdro obklopující sluchové kůstky středního ucha. Zatímco psotvárné šelmy mají bubínkovou výduť jednokomorovou, v případě kočkotvárných šelem je vytvořena výduť dvoukomorová, jejíž části jsou odděleny přepážkou (septem). U vyhynulé kočkotvárné skupiny nimravidů byla bubínková výduť chrupavčitá, případně postrádala pravé septum pozorované u ostatních kočkotvárných.[82][83] U žijících nandinií není výduť plně osifikována. Samotná kostěná bubínková výduť a její fúze s lebeční bází zřejmě představuje znak, jenž se v případě šelem vyvinul několikrát.[84]

Pohyb dolní čelisti šelem bývá omezen na svislou rovinu; u mnoha šelem je nicméně mandibulární symfýza spíše fibrózní, kvůli čemuž se mohou obě poloviny dolní čelisti do určité míry pohybovat i do stran. Čelisti se mohou sevřít se značnou silou, což zajišťuje dobře vyvinutý sval žvýkací (musculus masseter), a především pak sval spánkový (musculus temporalis). Pro upnutí mohutného spánkového svalu se na lebce vyvinuly přidružené struktury, jako je šípový hřeben.[85][80] Délka čelistí a lebky se mezi jednotlivými skupinami různí, a závisí mj. i na způsobu života jednotlivých šelem. Z hlediska anatomického umožňuje zkrácená lebka (jako u kočkovitých) soustředit sílu spíše na přední zuby, což pomáhá rychle zabít kořist, zatímco u druhů s prodlouženými lebkami (jako jsou psovití) je síla soustředěna spíše na zadní žvýkací zuby.[86]
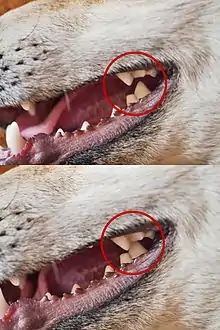
Základní zubní vzorec šelem je 3.1.4.33.1.4.3 × 2 = 44 zubů, nicméně u většiny šelem docházelo k dalším redukcím chrupu, takže výsledný počet zubů bývá nižší. Snižuje se především počet třenových zubů (premolárů) a stoliček.[87][88] Například kočkovité a hyenovité šelmy mají pouze jedinou horní stoličku, zatímco ostatní šelmy mívají stoliček více; rovněž primitivní počet čtyř premolárů bývá u většiny šelem redukován, a zatímco u některých druhů jsou premoláry relativně rozvinuté, u jiných jsou spíše malé.[85] Ploutvonožci mívají méně zubů než suchozemské šelmy; navíc počet zubů se může v případě ploutvonožců lišit i v rámci jednoho druhu.[80]
Většina druhů šelem má tři řezáky na jeden kvadrant čelisti, přičemž třetí řezák bývá vždy větší než první dva. Řezáky šelem jsou silné v úchopu a mohou sloužit k odtrhávání kůže a svalstva z mrtvého zvířete.[85] Špičáky slouží primárně k zabití kořisti. Podobně jako v případě délky lebky, i vzhled a tvar průřezu špičáků se liší v rámci jednotlivých skupin šelem; jakožto klíčová součást chrupu však bývají obecně velmi odolné. Špičáky kočkovitých jsou v průřezu spíše kulaté, u psovitých naopak eliptické.[86] U některých skupin kočkotvárných se objevily značně prodloužené, zakřivené a laterálně zploštělé šavlovité špičáky (někteří machairodi, barbourofelidé, …),[89] u mrože se pak horní špičáky přeměnily v mohutné kly.[90] Jsou-li čelisti zavřené, dolní špičáky šelem zapadají do diastemy mezi třetím řezákem a prvním špičákem v horní čelisti, horní špičáky pak do diastemy mezi prvním premolárem a špičákem v čelisti dolní.[85] U šelem se objevuje mléčný chrup, jenž předchází chrupu trvalému.[91]
Typickým znakem šelem je modifikace posledního třenového zubu horní čelisti a první stoličky dolní čelisti; tyto zuby dohromady vytvářejí tzv. trhákový komplex. Trháky jsou upraveny tak, že jako nůžky vzájemně působí proti sobě a pomáhají svému nositeli ukrajovat kusy masa ze zabité kořisti.[80][92] Tato klasická synapomorfie šelem je evolučně velmi stará a objevuje se již u paleocenních předků ze skupiny Viverravidae.[93] U šelem, které více navykly všežravému způsobu života, jsou trháky více přizpůsobeny i drcení ostatní potravy, nejen ke krájení masa. Trháky zcela ztratili všichni recentní ploutvonožci;[94] všechny zuby ploutvonožců vyjma řezáků a špičáků jsou homodontní a jejich stavba je spíše jednoduchá.[95]
Šelmy mívají velké a složité nosní skořepy, které poskytují značnou plochu pro čichové receptory, přičemž zvláště u psotvárných a kočkovitých šelem je čich velmi dobře vyvinut. Velmi dobrý je rovněž zrak, přičemž vidění je binokulární. U nočních šelem se mezi sítnicí a cévnatkou oka objevuje odrazivá vrstva tapetum lucidum, jež usnadňuje vidění za ztížených světelných podmínek.[9][96][79][97] Většina suchozemských savců má na sítnici pouze dva typy očních čípků (dichromatické vidění), u některých šelem se nicméně objevuje i monochromatičnost (pozorována primárně u ploutvonožců, dále např. u mývala severního nebo kynkažu).[98] Sluch je u většiny šelem vynikající, některé šelmy (především druhy lovící malé hlodavce) jsou schopny vnímat i ultrazvuk vydávaný jejich kořistí. Hmatové podněty jsou zaznamenávány u mnohých šelem skrze vibrisy, které vyrůstají především z obličeje, ale mohou se objevovat i na končetinách.[99]
Tělo



Šelmy jsou, co se týče různých biologických znaků, zřejmě nejrozmanitějším řádem mezi savci. Hmotnost těla osciluje mezi pěti řádovými velikostmi, od ca 50 g u lasice kolčavy (Mustela nivalis) po ca 5 000 kg u rypouše sloního (Mirounga leonina). Značnou variabilitu lze pozorovat dokonce i v rámci jednoho druhu; například hmotnost vlka obecného (Canis lupus) se v rámci celého areálu výskytu pohybuje mezi 15 a 80 kg.[7][100] U většiny druhů bývají samci větší než samice (pohlavní dimorfismus).[99]
Značně se liší samotný tvar těla a vzhled jednotlivých skupin. Mnoho šelem, jako jsou někteří zástupci lasicovitých, skunkovitých či promykovitých, se konvergentně vyvinulo v menší lovce s dlouhými a ohebnými těly, kratšíma nohama a dlouhým ocasem, což jim umožňuje pronikat i do menších prostor; i když i zde lze pozorovat mohutné druhy, jako je rosomák sibiřský (Gulo gulo). Naopak medvědovití bývají vesměs silné a mohutné šelmy, medvěd lední (Ursus maritimus) může dosáhnout hmotnosti až 800 kg. V rámci celého řádu jsou aktivní predaci nejvíce přizpůsobeni kočkovití. Ti mají dobře osvalené přední končetiny se zatahovacími drápy, které umožňují strhnutí kořisti, prstochodeckou lokomoci a různé adaptace lebky, které usnadňují zabití a sežrání ulovených zvířat.[100][101] Významné změny anatomie podstoupili ploutvonožci adaptovaní na život ve vodě. Jejich tělo bývá větší, což pomáhá udržení tělesné teploty v chladných vodách, a má hladký torpédovitý tvar, aby ve vodě kladlo co nejmenší odpor. Končetiny jsou tvarem přeměněny v ploutve a vyrůstají z těla distálně od lokte, resp. kolene. Ocas je rudimentární,[75][102] zadní končetiny proto míří dozadu a ocasní ploutev funkčně nahrazují. Tělo ploutvonožců bývá izolováno tukovou vrstvou a je rozličnými způsoby přizpůsobeno k potápění.[103]
Postkraniální skelet je u šelem spíše nediferencovaný, jedinou značně odlišnou skupinou jsou zmiňovaní ploutvonožci.[104] V rámci kostry je u všech šelem klíční kost zcela nevyvinutá, anebo pouze zbytková, končetiny tedy nejsou schopny bočního pohybu v podobném rozsahu, jako je tomu například u primátů. Dalším charakteristickým znakem kostry těla je fúze některých zápěstních kůstek (poloměsíčitá kost a člunkovitá kost), které dohromady vytvářejí jedinou strukturu.[7][79] Kosti předloktí a holeně naopak nesrůstají.[105] Končetiny jsou kratší u stromových, polovodních a částečně stromových druhů ve srovnání s čistě pozemními druhy. Končetiny mohou být vybaveny pěti prsty, případně může být jeden prst redukován. V takovém případě se vždy jedná o palec.[91] U suchozemských šelem lze – v závislosti na skupině – pozorovat buďto prstochodectví, anebo ploskochodectví. Přední končetiny mají pět prstů, první však bývá pouze zbytkový. V případě ploskochodců jsou zadní končetiny vybaveny pěti prsty, v případě prstochodců je počet prstů zadních končetin redukován na čtyři. Šelmy adaptované na prstochodectví, jako jsou kočkovití či psovití, bývají většinou zdatnými běžci či sprintery.[7][79] Prsty šelem jsou vybaveny drápy, které se u některých skupin mohou zatáhnout. Tato schopnost možná představuje prastarou synapomorfii celého řádu, různé skupiny ji však v průběhu vývoje potlačily, anebo ztratily.[84] Samci šelem mají pyjovou kost, jež umožňuje prodlouženou kopulaci. U některých skupin, jako u kočkovitých a hyenovitých, je pyjová kost pouze zbytková, anebo zcela chybí; u jiných šelem (jako je mrož) může být i přes 50 cm dlouhá.[7][80]
Biologie a ekologie
Sociální struktura

Co se týče škály chování a sociálních systémů, šelmy v něm mezi savci převyšuje pouze řád primátů.[106] Asi 85–90 % druhů šelem žije v dospělosti samotářským způsobem života, přičemž jednotlivci se společně vyhledávají pouze během rozmnožování nebo obrany teritoria. Tento trend je vysvětlitelný tím, že většina predátorů loví efektivněji sama (mnoho šelem zabíjí spíše menší kořist), a zároveň mláďatům většiny savců stačí pouze péče jejich matky. Běžná sociální organizace šelem je taková, že si jeden dominantní samec vydržuje území, které chrání proti ostatním samcům, přičemž toto území se může překrývat s menšími územími samic (v nichž případně žijí i mláďata).[81][107] Velikost domovského území a teritoria (tj. oblasti, již šelmy brání před ostatními zástupci stejného druhu) se může samozřejmě mezi druhy značně různit, a to až o šest řádových velikostí, od malých teritorií nosálů až po teritorium medvěda ledního, jež může zabírat i více než 190 000 km2. Obecně platí, že hypermasožravé formy si nárokují větší území než šelmy všežravé či plodožravé. Velikost teritoria se však může značně lišit i u různých populací v rámci jednoho druhu, například vlivem různé zeměpisné šířky a s ní spojené úživnosti stanoviště. Primárně masožravé druhy vykazují v tomto případě opět největších rozdílů.[108]
I u solitérně žijících druhů lze nicméně pozorovat určitou formu sociality v pozadí, komplikovanější sociální systémy, případně jistou úroveň flexibility. Například medvědi hnědí se sdružují na severozápadním pobřeží Severní Ameriky během pravidelné migrace lososů, ač jinak se jedná o samotářsky žijící šelmy.[106] U afrických nandinií se domovské oblasti samců často překrývají, nicméně fakticky jde o několik odlišných sektorů. Dominantní samec pravidelně obchází všechny tyto sektory, zatímco podřízení samci pobývají pouze na svém území a snaží se vyhýbat dominantnímu samci.[109]
_2.jpg.webp)
Přibližně 10–15 % druhů šelem vytváří sociální skupiny. Ekologických faktorů, které podporují skupinový život, je několik. Jedním z nich je hojnost a rozmístění zdrojů potravy, přičemž princip byl formulován jako hypotéza o rozptýlení zdrojů („The Resource Dispersion Hypothesis“; RDH). Původně vychází především z pozorování jezevců lesních, kteří žijí ve skupinách, aniž by jim to přinášelo významné výhody.[110] Základní myšlenkou hypotézy je, že si jedinec (nebo jiná sociální jednotka) vydržuje tak velké území, aby mu s jistou kritickou pravděpodobností za dané časové období poskytlo alespoň jednu plošku s dostatečným množstvím zdrojů. Stejná oblast však může – alespoň po určitou dobu – poskytovat dostatečné množství zdrojů pro více jedinců, aniž by se tím snížila jejich biologická zdatnost.[111] Mezi další selekční tlaky, které mohou podporovat různé formy skupinového života, lze zařadit nadměrné výdaje na rozptyl jednotlivců z rodné skupiny, početní výhodu skupiny během lovu velké kořisti nebo obrany teritoria, lepší ochranu proti predátorům nebo společnou péči o mláďata.[110] Život ve skupině naopak může vést k vyššímu riziku infekčních onemocnění nebo zvýšené konkurenci.[111]
Skupinový život se objevuje u mnohých čeledí šelem, ač existují i čistě samotářské čeledi, jako jsou například medvědovití.[81] Formy sociální struktury mohou být značně rozmanité, a to i v rámci jedné čeledi. Většina kočkovitých šelem je striktně solitérní, výjimkou je v tomto případě gepard štíhlý, jehož samci vytvářejí menší koalice, jež spolupracují na ochraně území, při lovu a obraně kořisti, a především vysoce sociální lev.[106] Lev kapský vytváří polygamní smečky tvořené 1 až 21 dospělými samicemi, jejich mláďaty/dospívajícími jedinci a typicky dvěma dospělými samci, kteří smečku hájí. Samice často rodí synchronně a mláďata jsou vychovávána komunálně. Při převzetí smečky cizími samci je častá infanticida.[112] Sociální systém psovitých bývá založen spíše na monogamii; jde navíc o jediné šelmy, u nichž se samci podílí na péči o mláďata. Psi hyenoví žijí ve stmelených smečkách o 20 a více jednotlivcích, rozmnožuje se zde však většinou pouze jediná samice. Vlci mohou rovněž vytvářet podobné smečky, ty ale bývají založeny spíše na štěpně-fúzním („fission-fusion“) základu.[81][106] Hyena skvrnitá vytváří klany o až 80 jedincích, které vede dominantní samice.[113] Hyenka hřivnatá naopak vytváří monogamní systém, ač samice se může nakrývat i se samci v okolí. Promykovití bývají spíše samotářské šelmy, ale mangusta žíhaná, mangusta trpasličí a surikata tvoří kolonie, v nichž se rozmnožuje několik samic, respektive dominantní pár (u mangusty trpasličí).[106]
Rozmnožování
.jpg.webp)
U šelem lze nejčastěji pozorovat polygamní rozmnožovací systém, kdy se většinou samci páří s větším množstvím samic. U podobných (polygynních) druhů se často rozvíjí zřetelný pohlavní dimorfismus, kdy jsou samci značně větší nežli samice a mnohdy se odlišují i některými výraznými sekundárními pohlavními znaky, jako je hříva u samců lva nebo výrazný chobot v případě samců rypouše sloního.[114] U rypoušů sloních je polygynie vyhnána do krajnosti nejen mezi šelmami ale savci obecně: vyspělí samci mohou svou hmotností více než desetkrát převyšovat pohlavně dospělé samice. Přibližně pouhá desetina samců rypoušů se dožije pohlavní dospělosti, přičemž pouze 2–3 % nejsilnějších samců se pravděpodobně rozmnoží; nicméně v takovém případě se mohou spářit i s více než stovkou samic za jediný rok.[115] Monogamie je v rámci šelem výrazně vzácnější a objevuje se u zmíněných psovitých nebo některých promykovitých. V některých případech se rozmnožuje pouze jeden pár a ostatní dospělci pomáhají s péčí o mláďata.[114]

Vzhledem k samotářské povaze mnohých šelem představuje problém, aby samec nakryl samici v plodném období. U rozličných druhů šelem samice pro informování samců hojně využívají pachových značek (v tomto případě jde především o rozstřikování moči), přičemž frekvence značkování se výrazně zvyšuje, když u samice nastupuje říje.[116] Další strategií samic některých šelem je takzvaná indukovaná ovulace, tj. samice přicházejí do říje, ale uvolnění vajíček je stimulováno až samotnou kopulací se samcem. Právě za účelem prodloužení pářícího aktu se samcům rozvíjí pyjová kost.[114][105] Samice suchozemských šelem se vyznačují dvourohou dělohou a pásovou placentou.[91]
Rozmnožování může probíhat buďto sezónně, anebo celoročně, přičemž v chladném podnebí bývá načasováno tak, aby mláďata přicházela na svět během jara a léta. Většina druhů rodí pouze jediný vrh mláďat za jeden až dva roky. Obecně platí, že menší druhy šelem se mohou rozmnožovat častěji a rodí i větší vrhy mláďat, zatímco u větších šelem, jako jsou například medvědovití nebo ploutvonožci, se samice páří i v několikaletých intervalech a rodí často jen jediné mládě.[99][117] Optimální načasování porodů mohou samice některých druhů řešit odloženou nidací, tj. vývoj zárodku se po oplození na určitou dobu pozastaví ve stádiu blastocysty. Odložená nidace převažuje například u ploutvonožců. Ti se v teplých měsících sdružují na ostrovech nebo v pobřežních oblastech, přičemž samci sem vnikají o něco dříve než samice, aby si mohli vymezit teritoria. Samice připlouvají těsně před porodem mláďat, načež se po porodu opětovně spáří, ale zárodek se v děložní stěně uchytí až za několik měsíců, aby mládě samice porodila opět až další rok.[114][118]
Samotná délka březosti, stupeň vývoje mláďat a délka rodičovské péče se může u jednotlivých šelem značně lišit. Například u tuleňovitých bývají mláďata vysoce prekociální (mládě tuleně obecného je schopno plavat již několik minut po narození) a rostou velice rychle, samice je výživným mateřským mlékem kojí pouze několik týdnů.[99][114] Mláďata suchozemských šelem se naopak rodí slepá a s řídkým osrstěním.[105] V případě mnohých druhů se navíc objevuje i delší období učení a zrání, kdy mláďata čerpají dovednosti do různých situací budoucího života. Tato potřeba je výsledkem vysokého poměru hmotnosti mozku k tělesné hmotnosti šelem.[117] Většina šelem se dožívá více než 10 let, ač některé lasicovité šelmy (Mustela sp.) se vyznačují kratší délkou života. Samice kolčavy dosahují pohlavní dospělosti již asi ve třech měsících a ve volné přírodě přežívají často méně než jeden rok. Naopak ploutvonožci mohou ve volné přírodě žít údajně i více než 40 let.[99][114]
Komunikace

Šelmy mají, i vzhledem k rozvinutým smyslům, značně pokročilé komunikační schopnosti. Komunikace probíhá na bázi akustické, vizuální i taktilní, především však ale olfaktorické (čichové), která má pro šelmy řadu výhod. Některé pachové značky mají tu důležitou vlastnost, že poskytují informaci i v nepřítomnosti jejich původce. Pachové signály slouží především pro vymezení území a pro sdílení informací o identitě, sociálním postavení a reprodukčním stavu.[99][80][119]
Coby pachové značky může sloužit vlastní moč a výkaly. Šelmy tímto způsobem značkují – často opakovaně – většinou některé nápadné objekty a strategická místa teritoria, přičemž objem moči či výkalů bývá často pouze symbolický. Takovéto teritoriální chování bylo zaznamenáno u mnohých čeledí šelem, hojné je například u některých psovitých, jako jsou lišky nebo vlci. Dominantní samice psů pralesních pravidelně močí na převislou vegetaci kolem stezek, přičemž moč následně na stezky odkapává, a zanechává tak pachovou stopu. Mnoho druhů šelem může také na určitých místech pravidelně defekovat a hromaděním výkalů zde vytvářet latríny. Podobné chování lze pozorovat například u jezevců lesních. Ti v některých částech svého areálu výskytu vytvářejí sociální skupiny, přičemž jejich teritoria vymezují právě podobné latríny, do nichž všichni dospělci ze skupiny pravidelně defekují. Množství latrín na hranicích teritoria přitom ovlivňuje počet jezevců ze sousední skupiny.[120][121]
Šelmy jsou zároveň schopny vytvářet vlastní aromatické sekrety coby produkt specializovaných žláz. K tomuto účelů bývají přizpůsobeny především žlázy mazové a potní (resp. apokrinní), jež bývají soustředěny do oblasti tváří, ocasu, hráze a řitního otvoru. Sekrety uvolňované z mazových žlázek bývají spíše olejovité, šelmy je používají k označkování předmětů, případně i jedinců stejného druhu. Apokrinní žlázy naopak vylučují látky těkavější povahy, jež jsou vhodné ke krátkodobější signalizaci. Pachové žlázy mohou být součástí anatomicky složitější struktury, jako jsou například zásobní pachové váčky mnohých šelem ústící v anální oblasti.[122] Hyeny čabrakové jsou známé specifickým značkováním, kdy ze svého specializovaného análního vaku vylučují na stonky rostlin výměšky mazových i apokrinních žláz, přičemž každý z nich má jiné zbarvení a asi i účel. Výměšek apokrinních žláz je černý, výměšek mazových žláz je bílý (hyeny jej umísťují níže).[123] Je možné, že bílá pasta představuje varování pro případné konkurenční skupiny, kdežto černá spíše slouží k vnitroskupinové komunikaci.[120]
Vyjma značkování teritoria mohou být sekrety využívány i ke vzájemnému sdílení pachů mezi jedinci stejného druhu. Toto chování se nazývá allomarking a lze jej opět demonstrovat na příkladu jezevce lesního. U jezevců lze pozorovat dva typy allomarkingu. Buďto jednotliví jezevci označují podocasní žlázou tělo druhého jedince, anebo si vzájemně tisknou podocasní žlázy k sobě.[121] Některé šelmy nakonec využívají pachů i jako specifické obrany proti nepřátelům a z análních váčků jsou schopny vystřikovat odporně páchnoucí sekrety. Mezi ně lze zařadit například medojeda, a především skunkovité šelmy (včetně „smrdutých jezevců“ Mydaus sp. z jihovýchodní Asie[124]).[125]
.jpg.webp)
Šelmy ke vzájemné komunikaci využívají i širokou škálu různých vokalizací, jež mohou sloužit k utužování sociálních vazeb, obraně teritoria před vetřelci, vábení jedinců opačného pohlaví, stejně jako k projevení emocí, případně coby varování před možným nebezpečím. Některé společenské druhy mohou vokální komunikaci využívat ke vzájemnému svolávání či k udržování kontaktu mezi členy skupiny. Smečky dhoulů vydávají specifické pískavé zvuky, které jim pravděpodobně pomáhají udržovat vzájemný kontakt během lovu v hustých lesích.[99][126] Experimenty s nahrávkami na smečce lišek polárních z ostrova Mednyj demonstrují, jak šelmy dokáží rozlišovat mezi vokalizacemi příslušníků vlastní sociální skupiny a sousedních skupin. Vokalizace členů skupiny nevyvolávaly teritoriální odezvy, naopak docházelo k pozdravům. Nahrávky sousedních skupin vedly k teritoriálnímu chování, naopak pozdravy se nikdy neobjevily.[126][127] Vokalizace slouží rovněž ke kontaktu mezi matkou a mláďaty. Například pro všechny ploutvonožce jsou charakteristická kontaktní volání, pomocí nichž je samice schopna najít svého potomka po návratu z potravní výpravy zpět na souš (v případě některých lachtanovitých samice opouštějí mládě až na 7 dní).[128] U některých šelem pak lze pozorovat i případy složitější mezidruhové vokalizace, přičemž pozoruhodným případem je mutualismus mezi mangustami trpasličími a některými zoborožci; tyto druhy se v případě nebezpečí varují před predátory.[129]
Co se týče vlastností zvukových signálů, suchozemské šelmy dokáží vydávat zvuky od asi 50 Hz (řev lva) až po více než 100 kHz (tísňová volání mláďat fretek), ačkoli vokalizace dospělců ve většině případů nepřekračuje frekvenci 10 kHz. Rovněž zvuky vydávané ploutvonožci jsou většinou v pro člověka slyšitelném spektru, i když pocházejí záznamy o ultrazvuku (volání tuleně leopardího pod vodou) i infrazvuku (některé infrazvukové vibrace rypouše severního). Některé signály jsou velmi krátké a trvají jen několik desítek milisekund; jiné, jako vytí vlků, mohou trvat několik sekund. Kočka domácí dokáže nepřetržitě příst až 120 minut.[128][130]
U šelem lze nakonec pozorovat i komunikaci taktilní, tj. hmatová sdělení, a vizuální, která zahrnuje především držení těla a ocasu. Mimické schopnosti se v rámci skupin šelem liší; mimiku mají omezenou například lasicovití, naopak jejich řeč těla je mnohdy velmi výrazná.[99][126]
Potrava a její získávání

Šelmy mají jednoduchý žaludek a chybějící nebo zmenšené slepé střevo. Vzhledem k tomu, že maso představuje obecně stravitelnější potravu než jsou rostliny, u většiny šelem živících se primárně masem je zmenšena délka i vnitřní plocha střev.[80] Šelmy (a živočichové obecně), jejichž potravní skladba je z více než 70 % tvořena masem, jsou nazýváni hypermasožravci; tyto druhy obyčejně také vykazují specifické změny v anatomii lebky a chrupu, včetně zkrácení obličejové části lebky a redukce stoliček. Jedněmi z nejspecializovanějších hypermasožravců jsou kočkovité šelmy, z vyhynulých forem například nimravidé. Hypermasožravé formy je však možné nalézt v rámci mnoha skupin šelem, jako jsou medvědovití (medvěd lední), psovití (pes hyenový), hyenovití (hyena skvrnitá) nebo amphicyonidé (vyhynulá čeleď).[131]
Pouze málo druhů šelem jsou výhradní masožravci, strava většiny ostatních zástupců je pestřejší. Ve srovnání s kočkovitými bývají všežravější především šelmy medvědovité, cibetkovité, psovité či hyenovité. Flexibilní všežravé druhy, jako je kojot prérijní, mohou využívat širokou škálu potravních zdrojů, a prosperovat tak v rozličných oblastech. Dalšími typicky všežravými druhy jsou například šakal obecný, kuna skalní nebo medvěd hnědý. Některé šelmy se přizpůsobily konzumaci hmyzu; zvláště specializovaným druhem je z tohoto pohledu hyenka hřivnatá, jež se živí výhradně termity. Někteří ploutvonožci se, vyjma ryb a dalších obratlovců, soustředí na i na lov korýšů a měkkýšů, pro tuleně krabožravého představuje hlavní součást potravy kril. Některé šelmy jsou primárně plodožravé, a to především v tropických oblastech, kde je ovoce k dispozici během celého roku. Medvídkovitá šelma kynkažu ze Střední a Jižní Ameriky se živí takřka výhradně ovocem; pro africké nandinie tvoří plody asi 4/5 jídelníčku. Listy a stonky rostlin jsou špatně stravitelnou a málo výživnou potravou; v rámci šelem jsou jejich konzumaci nejvíce přizpůsobeny panda červená, jež však tento jídelníček prokládá častěji i jinou potravou, a především pak panda velká, přičemž oba dva druhy se živí primárně bambusem.[132][133] V případě pandy velké, jejíž trávicí trakt a složení střevního mikrobiomu není navzdory stravě podobné býložravcům, je denně zužitkováno pouhých 17 % z přijaté potravy.[134]
_with_springbok.jpg.webp)
U šelem, které získávají maso aktivním lovem, se objevují určité preference mezi vlastní tělesnou hmotností a tělesnou hmotností kořisti. Predátoři, kteří hmotnostně nepřesahují hranici 21,5 kg, se nejčastěji zaměřují na lov živočichů o více než polovinu lehčích, častou kořistí je z tohoto pohledu i hmyz. Predátoři o hmotnosti nad 21,5 kg naopak loví primárně zvířata o jim podobné tělesné hmotnosti.[132] Šelmy mohou lovit samotářsky, v párech nebo ve skupinách. Smečkový lov, jenž lze pozorovat například u vlků nebo psů hyenových, umožňuje ulovit větší nebezpečnější druhy, hmotnostně převyšující jednotlivé lovce. Metody lovu se v rámci jednotlivých šelem liší. Množství šelem běžně vyhledává spíše menší kořist, například pomocí čichu. Kočkovité šelmy se spoléhají na ostrý zrak, loví ze zálohy a využívají náhlého momentu překvapení. U podobných druhů se často objevují různé vzory na srsti, které jim poskytují kamufláž a ztěžují potenciální kořisti spatření lovce. Lasice svou kořist nejprve určitou dobu sledují a následně se ji kratším během snaží uchvátit. Pro některé psovité nebo hyenovité šelmy je pak typická delší štvanice.[81][135] Menší živočichy šelmy obvykle jednoduše rozkoušou a sežerou; jindy lze pozorovat specifické způsoby zabíjení. Například kočkovité šelmy uloveným obratlovcům často přeruší míchu, větší kořist pak zabíjejí prokousnutím hrdla, nebo ji udusí stiskem čenichu a tlamy. Naopak smečkoví psi hyenoví se nesoustředí na krk oběti, ale štvané zvíře napadají spíše zezadu.[81][136]

Maso je sice výživnou, ale ne tak snadno dostupnou potravou. Mnoho druhů šelem je adaptováno na určité období hladovění, u zabitého těla zvířete však následně dovedou zkonzumovat velké množství masa. Hyena skvrnitá je schopna sežrat objem masa o až třetině její tělesné hmotnosti. Navíc má nesmírně silné čelisti, s pomocí nichž je schopna rozdrtit kosti a sežrat i výživný morek.[133][136] Vyjma samotného ulovení kořisti představuje pro šelmy problém i kleptoparazitismus, především pak v otevřených oblastech. Kupříkladu v afrických savanách si mohou kořist ulovenou gepardy nebo levharty často přivlastnit větší a silnější predátoři. Levharti tento problém řeší tak, že se snaží mrtvá zvířata vytáhnout do korun stromů. Mnoho druhů šelem si nesežrané zbytky ulovených zvířat ukrývá před ostatními masožravci na skrytá místa, aby je mohly sežrat později.[132]
Některé druhy šelem se vyznačují specifickými znaky, které jim umožňují konzumaci jejich preferované potravy. Například medvěd pyskatý, jenž se soustředí primárně na lov mravenců a termitů, má dlouhé drápy, pomocí nichž může pronikat do termitišť. V horní čelisti mu scházejí dva přední řezáky, díky čemuž může snadněji nasávat hmyz do tlamy.[137] Specializovanou strukturou, jež slouží k získávání potravy, je například také radiální sezamoidní kůstka, funkčně šestý prst, jenž se konvergentně vyvinul u pand velkých i pand malých a slouží k přidržování bambusových stébel. Někdy je nazýván také pandí palec. Tato struktura se objevuje i u rodu Simocyon a původně pravděpodobně sloužila k pohybu ve větvích, teprve druhotně ji pandy začaly využívat k získávání potravy.[72]
Výskyt a vliv na ekosystém

Šelmy jsou rozšířeny takřka celosvětově od pólu k pólu a vyskytují se prakticky ve všech biotopech, ať už jsou to savany, pouště, různé typy lesů, studená tundra i mořský led. Některé šelmy, jako jsou vydry, žijí v řekách a jezerech, zatímco ploutvonožci většinou loví na otevřeném moři, z toho rypouš sloní se může potopit až do hloubky okolo 1 500 metrů.[9][138] Některé šelmy žijí synantropně, případně byly domestikovány.
Šelmy mají pro své životní prostředí klíčový vliv a přispívají k dynamické rovnováze ekosystémů. Pomáhají regulovat množství kořisti, vyrovnávají výkyvy v její početnosti, z populace třídí slabé a nemocné kusy; tyto dopady jsou navíc často ovlivňovány i podmínkami prostředí (tuhé zimy, sucha). Šelmy tímto způsobem obecně pomáhají udržovat populace kořisti zdravější. Velké druhy šelem snižují množství kopytníků, a tím omezují jejich dopad na vegetaci, čímž umožňují udržovat rozmanitější rostlinná společenstva (a spolu s nimi pak i společenstva živočišná). Menší druhy šelem mají zase velký vliv na množství hlodavců, navíc bývají plodožravější, čímž představují významné rozptylovače semen. Počty samotných masožravců jsou pak omezeny dostupností potravy, přítomností větších predátorů nebo nemocemi.[9][80][138]

Vliv šelem na ekosystém demonstruje několik historických příkladů. V polovině 90. let 20. století byl do Yellowstonského národního parku znovuvysazen vlk obecný, jenž z oblasti vymizel asi 70 let předtím. Před reintrodukcí vlků v oblasti značně kolísala početnost jelenů wapiti; ta v průběhu desítek let střídavě procházela populační explozí v příhodných obdobích, ale během tuhých zim velká jelení stáda hynula kvůli nedostatku potravy. Výskyt vlků se výrazně podepsal na poklesu jelení populace, což se mimo jiné promítlo na růstu rozmanitosti flóry pastvin, snížení eroze podél říčních toků a rozkvětu pobřežních ekosystémů. Populace jelenů se po reintrodukci vlků dlouhodobě stabilizovala na 6 000 až 8 000 kusech, přičemž tato stáda již nepostihují extrémní výkyvy. S přítomností vlků se rovněž snížila početnost kojotů, pro něž představují vlci přirozené predátory; s jejich úbytkem naopak došlo ke zvýšení početnosti vidlorohů. Další ukázkou vlivu šelem je úbytek ploutvonožců a mořských vyder v oblasti severního Tichomoří. Po druhé světové válce klesla vlivem velrybářství populace velryb. Jejich predátoři, kosatky, se pak musely přeorientovat na lov mořských šelem. Vydry jsou však přirozenými predátory ježovek, proto úbytek vyder vedl i k přemnožení ježovek živících se místními chaluhovými lesy, které byly tímto způsobem narušeny. To mělo negativní vliv na místní populace bezobratlých, ryb a mořských ptáků. Navíc chaluhy jsou významným pohlcovačem oxidu uhličitého a přítomnost či nepřítomnost mořských vyder tedy ovlivňuje globální klima.[9][139] Podobný vliv vyder byl popisován i v jedné hypotéze, mající osvětlit vyhynutí korouna bezzubého (Hydrodamalis gigas), sirény z oblasti Komandorských ostrovů. Lov vyder ze strany lidí, následná populační exploze ježovek rodu Strongylocentrotus a úbytek vodních řas, jimiž se koroun živil, mohly představovat ekologický tlak, jenž nakonec vedl k vyhynutí sirény (spolu s lovem lidmi).[140]
Naopak katastrofou pro původní faunu může skončit introdukce šelem do izolovaných stanovišť, převážně na ostrovy, kde živočichové do té doby nepodléhali predačnímu tlaku. Zavlečení predátorů do uzavřených ostrovních ekosystémů vedlo k úbytku či vyhynutí velkého množství původních druhů.[9] Vliv predace demonstruje například vyhynutí pokřovníka ostrovního (Traversia lyalli), nelétavého ptáka, jehož reliktní populace žily na Stephensově ostrově u Nového Zélandu. Vlivem jediné březí kočky, která sem byla dovezena na začátku roku 1894, na ostrově vzešla početná populace koček, jež se výrazně podepsala na stavu místní fauny, přičemž pokřovník ostrovní vyhynul do několika let.[141] Častými oběťmi podobných biologických invazí domácích koček, ale i lišek, psů, lasic či promyk, byly právě endemitní druhy ptáků, avšak i někteří savci nebo plazi.[81]
Vztah s lidmi
Odraz v kultuře
Šelmy mají v lidské kultuře prakticky od počátku dvojí roli a jsou zvířaty uctívanými i pronásledovanými. Šelmy vzbuzovaly obdiv pro svou krásu i lovecké dovednosti a na základě toho si rozličné druhy šelem zajistily důležité postavení v mnohých kulturách.[142] Například pro evropskou civilizaci představoval lev dokonce „krále zvířat“, jehož zmiňují nesčetněkrát dobové prameny eurasijského i afrického kontinentu (jenom v bibli je zmíněn více než stokrát); naopak směrem na východ tuto roli přebíral tygr, „pán všeho tvorstva“, „král džunglí a pralesů“, „car tajgy“, mytické zvíře, pro některá etnika dokonce božstvo.[143] Vyjma různých suchozemských šelem mají v kultuře významné místo i ploutvonožci, kteří jsou výrazně zakořeněni například v kultuře Inuitů (bohyně Sedna)[144] či v tradicích severozápadní Evropy (selkie).[145] Šelmy pro své vlastnosti často vystupují jako heraldická zvířata, například jako státní symboly nebo jako erbovní zvířata královských rodin. Během historie byly různé šelmy člověkem často loveny pro maso nebo kožešiny, později i jako trofejní zvířata. Domestikace dala vzniknout domácím kočkám nebo psům, s pozdějším rozličným množstvím vyšlechtěných plemen, která slouží člověku k různým účelům.[142]

 Velký státní znak České republiky se dvěma stříbrnými dvouocasými lvy
Velký státní znak České republiky se dvěma stříbrnými dvouocasými lvy Medvěd na znaku města Berlína
Medvěd na znaku města Berlína Soška bohyně Sedna
Soška bohyně Sedna Panda červená na historické indické poštovní známce
Panda červená na historické indické poštovní známce
Konkurence s člověkem, lov
Na druhou stranu však šelmy představují prakticky odjakživa konkurenty člověka. S rozvojem zemědělství a chovu zvířat se tyto konflikty prohloubily, protože šelmy všech velikostí mají tendenci zabíjet chovaná zvířata, která jsou pro člověka z ekonomického hlediska důležitá, například drůbež a dobytek. Větší druhy mohou obrátit pozornost i na člověka a stát se lidožrouty, a to včetně některých extrémních případů, jako byl čampávatský lidožrout. Snaha o snížení počtu predátorů probíhala tisíce let. S rozvojem účinnějších metod zabíjení (střelné zbraně, jedy) se lov zrychlil a došlo k radikálnímu zmenšení populací mnohých druhů. Eliminace šelem byla často organizována a financována státem, například koloniální správou.[142][146]

Lov šelem v současnosti probíhá ze dvou hlavních důvodů, které se mohou prolínat. Prvním z nich je domnělé, anebo skutečné ohrožení hospodářských zvířat, případně lidí. V takovém případě se zabíjení soustředí především na velké druhy šelem;[147] nicméně hospodářské škody, jež velké šelmy způsobují, mohou být menší ve srovnání s kumulativním účinkem většího počtu menších druhů (lišky, šakalové, kojoti). Například ve Spojených státech je 79 % škod na ovcích a 83 % škod na skotu přičítáno psovitým šelmám, z toho 15 %, resp. 18 % je způsobeno domácími psy, zbytek většinou kojoty.[148] Vyjma odvetného zabíjení představuje významnou hrozbu lov šelem za účelem získávání částí těl, k němuž sice dochází globálně, ale rozšířen je převážně v asijských zemích, kde části těl zvířat představují cennou komoditu, protože jsou důležitou součástí tradiční medicíny.[149] Neudržitelný lov se výrazně podepsal například na populaci tygrů, pro něž představuje primární hrozbu. Jejich nepřítomnost na zhruba 1 milionu čtverečních kilometrů vhodných stanovišť svědčí o výrazném vlivu pytláctví na tygří populace.[147] Lokální hrozbou může pro populace šelem být rovněž odstřel pro trofeje nebo kožešiny.[149] Ještě ve 20. století představoval lov pro kožešiny či olej výraznou hrozbu pro mnohé tuleňovité, a teprve výrazný globální úbytek tuleňů po druhé světové válce vedl k přijetí mezinárodních dohod omezujících lov, které vedly ke zotavení populací některých vážně ohrožených druhů.[150] Vyjma ploutvonožců byly ze stejného důvodu pronásledovány i některé lasicovité šelmy. Například vydry mořské (Enhydra lutris) byly lovem probíhajícím od konce 18. století prakticky vyhubeny, smlouva zakazující jejich komerční lov byla podepsána Spojenými státy, Velkou Británií, Ruskem a Japonskem až v roce 1911.[151] Nadále však probíhal lov jiných druhů, například jenom mezi lety 1910 až 1954 bylo z Chile vyvezeno na 38 000 kožešin zdejších mořských vyder náležících ke druhům vydra pobřežní a vydra jižní. Další omezení a obecný pokles poptávky na trhu s kožešinami však tyto hrozby postupně eliminovaly.[152]
Další hrozby a ochrana
.jpg.webp)
Primární hrozba pro většinu druhů šelem je ztráta přirozeného prostředí. Zbývající přírodní stanoviště ustupují zemědělským oblastem, pastvinám či městům, přičemž mizí odhadovanou rychlostí asi 1 % ročně. Přidruženou hrozbou je pak úbytek kořisti; i relativně neporušená stanoviště nemohou poskytovat útočiště pro šelmy, pokud z nich lidé vyčerpají potravní zdroje. Druhy citlivé k těmto hrozbám se vyznačují především velkými tělesnými rozměry, nízkou populační hustotou, neschopností rychlé obnovy populací či omezenými možnostmi rozptýlení, tj. většinou se jedná o velké masožravce/vrcholové predátory ekosystému, jako jsou medvědi, některé psovité šelmy, hyeny nebo kočkovití (ač při porovnání s jejich kořistí lze vztáhnout tyto proměnné i na mnohé menší druhy). Například pro udržení populace deseti tygrů ve vyváženém biotopu po dobu jednoho roku je zapotřebí alespoň 5000 kusů jejich kořisti, středních nebo velkých kopytníků, nepočítaje v to ostatní druhy šelem, jež s tygry v přirozeném prostředí koexistují. Roztříštění stanovišť a izolace menších populací může přinášet další problémy následkem hybridizace, inbreedingu či ztráty genetického bohatství v důsledku genetického driftu. Populace jsou navíc vystaveny zmiňovaným ohrožujícím faktorům, jako je uvedené pytláctví/odvetné zabíjení ze strany člověka.[147][153]
Dalším problematickým faktorem je šíření infekčních onemocnění, přičemž hojné druhy šelem mohou sloužit jako přenašeči chorob. Například mezi psovitými, skunkovitými, medvídkovitými či promykovitými může cirkulovat virus vztekliny, jenž může být přenesen na člověka nebo hospodářská zvířata. Výrazné škody působí zavlečení různých infekcí do menších populací vzácnějších druhů šelem: například domácím psům je přisuzováno zavlečení vztekliny a psinky do Serengeti, což se výrazně podepsalo na populacích lvů a divokých psů, a ze stejného důvodu byl dramatický úbytek vlčků etiopských v pohoří Bale.[154]

Šelmy představují pátý nejpočetnější řád savců, jenž zahrnuje přes 270 žijících druhů.[147][80] Na základě souhrnu dat od Mezinárodního svazu ochrany přírody (IUCN) z roku 2018 však spadá celkem 74 druhů šelem (včetně 10 ploutvonožců) do jednoho ze stupňů pro ohrožené druhy, přičemž 3 druhy jsou hodnoceny jako kriticky ohrožené, 28 patří mezi ohrožené druhy a 43 pak mezi druhy zranitelné. Čtveřice recentních druhů již byla vyhubena, a sice norek mořský (Neovison macrodon), lachtan japonský (Zalophus japonicus), tuleň karibský (Monachus tropicalis) a pes bojovný (Dusicyon australis). Oba druhy ploutvonožců byly vyhubeny do 50. let 20. století, norek mořský a pes bojovný již v poslední čtvrtině 19. století, přičemž za zmizení všech těchto druhů byl zodpovědný člověk.[147] Například pes bojovný, endemit Falklandských ostrovů, byl vybit chovateli ovcí do 80. let 19 století. A ač jiné druhy šelem stále přežívají, některé z nich byly z určitých částí areálu výskytu člověkem rovněž vymýceny.[81]
IUCN hraje zásadní koordinační roli v ochraně šelem, v rámci Komise pro přežití druhů IUCN (Species Survival Commission) byly zřízeny specializované skupiny pro psy, kočky, hyeny, vydry, lední medvědy, malé šelmy, ploutvonožce a velké evropské šelmy.[155] Mezinárodní obchod je korigován skrze úmluvu CITES, potřeba ochrany šelem se odráží v koordinovaných chovných programech v zajetí, které mohou být poslední šancí na přežití některých ohrožených druhů, a vytvářením sítě národních parků a rezervací ve volné přírodě. Některé druhy, které byly v minulosti člověkem vyhubeny, byly úspěšně reintrodukovány do původních biotopů (vlci v Yellowstonu). Snaha o ochranu se však musí soustředit i na eliminování nejhorších dopadů (postupující ztráta biotopů, lov) stále rostoucí lidské populace, která vyžaduje stále větší množství přírodních zdrojů. Mnoho šelem je totiž schopno existovat i v upravených stanovištích, a dokonce i velké šelmy se specifickými nároky na prostředí mohou přežívat v hospodářských oblastech, pokud dojde ke zmírnění problémů, které soužití mezi nimi a člověkem vytváří. Příkladem může být upravení metod chovu hospodářských zvířat tak, aby byla méně ohrožena ze strany útoků šelem, či zisk z příjmů z ekoturistiky.[99][147]
Vyjma druhů ohrožených lidskou činností existují i některé přizpůsobivé šelmy, které úspěšně přežívají v synantropních oblastech, a to někdy i navzdory snaze o jejich eliminaci. Existují populace lišek obecných, kojotů či mývalů, co dokáží prosperovat ve městech po boku člověka. Oviječ skvrnitý (Paradoxurus hermaphroditus), cibetkovitá šelma z jihovýchodní Asie, dokonce žije i v Hongkongu.[156]
Odkazy
Poznámky
- Podle novějších pohledů by měli být zástupci rodu Pseudaelurus děleni do alespoň dvou samostatných rodů. Druhy příbuzné moderním podčeledím kočkovitých mohou být řazeny spíše do rodu Styriofelis.[53]
- Nezaměňovat s podčeledí Amphicynodontinae. Anglické jméno skupiny Amphicyonidae, „bear dog“, poukazuje na její podobnost jak s psovitými, tak s medvědovitými šelmami. Česky je skupiny někdy nazývána „psi medvědovití“[68]
- Systematika vyhynulých druhů řazených k medvídkovitým je velmi kontroverzní.[73]
Reference
- Order Feræ Linnaeus, 1758 [online]. The Taxonomicon [cit. 2021-04-14]. [Classification by: Linnaeus (1758) Tomus I]. Dostupné online.
- Linnaeus, C. (1758). Sistema naturae per regna tria Naturae, secundum classes, ordines, genera, species, cum characteribus differentiis, synonimis locis. Tomus I. Impensis direct. Laurentii Salvii, Holmia. pp. 20–32.
- BOWDICH, Thomas Edward. An Analysis of the Natural Classifications of Mammalia: For the Use of Students and Travellers. [s.l.]: J. Smith, 1821. Dostupné online.
- carnivorous [online]. dictionary.com [cit. 2021-04-14]. Dostupné online. (anglicky)
- carnivora | Etymology, origin and meaning of the name carnivora by etymonline. www.etymonline.com [online]. [cit. 2021-11-29]. Dostupné online. (anglicky)
- SIMPSON, George Gaylord. The Principles of Classification and Classification of Mammals. New York: American Museum of Natural History (Volume 85), 1945. Dostupné online. (anglicky)
- McDade 2003, s. 256.
- Goswami a Friscia 2010, s. 6–7.
- VAN VALKENBURGH, Blaire; WAYNE, Robert K. Carnivores. Current Biology. 2010-11, roč. 20, čís. 21, s. R915–R919. Dostupné online [cit. 2021-07-14]. DOI 10.1016/j.cub.2010.09.013. (anglicky)
- Carnivora: Systematics [online]. ucmp.berkeley.edu [cit. 2021-07-14]. Dostupné online. (anglicky)
- Kirkwood 2013, s. 8.
- List of Marine Mammal Species and Subspecies [online]. The Society for Marine Mammalogy [cit. 2021-07-14]. Dostupné online. (anglicky)
- Vaughan, Ryan a Czaplawski 2015, s. 283.
- NAISH, D. Surprises From Placental Mammal Phylogeny 2: Skunks Are Not Weasels [online]. Scientific American, 2015-07-25 [cit. 2021-07-15]. Dostupné online. (anglicky)
- LAW, Chris J.; SLATER, Graham J.; MEHTA, Rita S. Lineage Diversity and Size Disparity in Musteloidea: Testing Patterns of Adaptive Radiation Using Molecular and Fossil-Based Methods. Systematic Biology. 2017-05-31, roč. 67, čís. 1, s. 127–144. Dostupné online [cit. 2021-07-15]. ISSN 1063-5157. DOI 10.1093/sysbio/syx047. (anglicky)
- Goswami a Friscia 2010, s. 64–66, 83.
- FAMILY Eupleridae [online]. Mammal Species of the World, 3rd Edition [cit. 2021-08-04]. Dostupné online. (anglicky)
- Goswami a Friscia 2010, s. 75.
- Goswami a Friscia 2010, s. 70.
- YODER, Anne D.; BURNS, Melissa M.; ZEHR, Sarah. Single origin of Malagasy Carnivora from an African ancestor. Nature. 2003-02, roč. 421, čís. 6924, s. 734–737. Dostupné online [cit. 2021-10-03]. ISSN 0028-0836. DOI 10.1038/nature01303. (anglicky)
- VERON, Géraldine; CATZEFLIS, François M. Phylogenetic relationships of the endemic malagasy carnivoreCryptoprocta ferox (Aeluroidea): DNA/DNA hybridization experiments. Journal of Mammalian Evolution. 1993-09, roč. 1, čís. 3, s. 169–185. Dostupné online [cit. 2021-10-03]. ISSN 1064-7554. DOI 10.1007/BF01024706. (anglicky)
- Feldhamer 2015, s. 806–838.
- Hunter a Barrett 2018, s. 7–11.
- Feldhamer 2015, s. 837.
- FAMILY Mustelidae [online]. Mammal Species of the World, 3rd Edition [cit. 2021-09-14]. Dostupné online. (anglicky)
- Gaisler a Zima 2007, s. 601.
- ZHOU, Xuming; XU, Shixia; XU, Junxiao. Phylogenomic Analysis Resolves the Interordinal Relationships and Rapid Diversification of the Laurasiatherian Mammals. Systematic Biology. 2012-01-01, roč. 61, čís. 1, s. 150–150. Dostupné online [cit. 2021-07-16]. ISSN 1063-5157. DOI 10.1093/sysbio/syr089. (anglicky)
- MURPHY, William J.; EIZIRIK, Eduardo; JOHNSON, Warren E. Molecular phylogenetics and the origins of placental mammals. Nature. 2001-02, roč. 409, čís. 6820, s. 614–618. Dostupné online [cit. 2021-07-16]. ISSN 1476-4687. DOI 10.1038/35054550. (anglicky)
- LV, Xue; HU, Jingyang; HU, Yiwen. Diverse phylogenomic datasets uncover a concordant scenario of laurasiatherian interordinal relationships. Molecular Phylogenetics and Evolution. 2021-04, roč. 157, s. 107065. Dostupné online [cit. 2021-07-16]. DOI 10.1016/j.ympev.2020.107065. (anglicky)
- SOLÉ, Floréal; SMITH, Thierry. Dispersals of placental carnivorous mammals (Carnivoramorpha, Oxyaenodonta & Hyaenodonta) near the Paleocene-Eocene boundary: a climatic and almost worldwide story. Geologica Belgica. 2013-01-01. Dostupné online [cit. 2021-07-16]. ISSN 1374-8505. (anglicky)
- Rose 2006, s. 130.
- HASSANIN, Alexandre; VÉRON, Géraldine; ROPIQUET, Anne. Evolutionary history of Carnivora (Mammalia, Laurasiatheria) inferred from mitochondrial genomes. bioRxiv. 2020-10-05, s. 2020.10.05.326090. Dostupné online [cit. 2021-07-22]. DOI 10.1101/2020.10.05.326090. (anglicky)
- Goswami a Friscia 2010, s. 37.
- Benton 2015, s. 379.
- Goswami a Friscia 2010, s. 17.
- HALLIDAY, Thomas J. D.; UPCHURCH, Paul; GOSWAMI, Anjali. Resolving the relationships of Paleocene placental mammals: Paleocene mammal phylogeny. Biological Reviews. 2017-02, roč. 92, čís. 1, s. 521–550. Dostupné online [cit. 2021-07-22]. DOI 10.1111/brv.12242. (anglicky)
- Therese Flink & Lars Werdelin (2022). Digital endocasts from two late Eocene carnivores shed light on the evolution of the brain at the origin of Carnivora. Papers in Palaeontology. 8 (2): e1422. doi: https://doi.org/10.1002/spp2.1422
- Benton 2015, s. 362.
- Kemp 2005, s. 247–250.
- Rose 2006, s. 120.
- Rose 2006, s. 121.
- Floréal Solé, Valentin Fischer, Kévin Le Verger, Bastien Mennecart, Robert P Speijer, Stéphane Peigné & Thierry Smith (2022). Evolution of European carnivorous mammal assemblages through the Palaeogene. Biological Journal of the Linnean Society. blac002. doi: https://doi.org/10.1093/biolinnean/blac002
- Goswami a Friscia 2010, s. 5–6.
- Rose 2006, s. 131–133.
- Goswami a Friscia 2010, s. 9–10.
- Antón 2013, s. 89–90.
- Turner 1997, s. 19.
- Antón 2013, s. 104.
- Antón 2013, s. 220.
- Goswami a Friscia 2010, s. 7.
- Turner 1997, s. 25.
- Kemp 2005, s. 259.
- Antón 2013, s. 107.
- Turner 1997, s. 26–27.
- Benton 2015, s. 390.
- Goswami a Friscia 2010, s. 9.
- Goswami a Friscia 2010, s. 8, 75.
- Vaughan, Ryan a Czaplawski 2015, s. 288, 290.
- Goswami a Friscia 2010, s. 8.
- Castelló 2020, s. 238.
- Vaughan, Ryan a Czaplawski 2015, s. 292.
- WANG, Xiaoming; TEDFORD, Richard. Dogs : their fossil relatives and evolutionary history. Paperback edition, 2010. vyd. New York: Columbia University Press, 2008. ISBN 978-0-231-50943-5, ISBN 0-231-50943-X. OCLC 822229250 S. 24. (anglicky)
- Goswami a Friscia 2010, s. 16–17.
- Rose 2006, s. 134–135.
- Kemp 2005, s. 260.
- Goswami a Friscia 2010, s. 15.
- Vaughan, Ryan a Czaplawski 2015, s. 296.
- Koexistence šavlozubých koček a medvědovitých psů [online]. ČRo Plus, 2012-07-12 [cit. 2021-08-10]. Dostupné online.
- Goswami a Friscia 2010, s. 12.
- Goswami a Friscia 2010, s. 11–12, 125–126.
- Vaughan, Ryan a Czaplawski 2015, s. 302.
- ROBOVSKÝ, Jan. Neopakovatelný pandí palec. Vesmír. 2008, roč. 87, čís. 4. Dostupné online.
- Goswami a Friscia 2010, s. 10.
- Vaughan, Ryan a Czaplawski 2015, s. 305.
- Vaughan, Ryan a Czaplawski 2015, s. 298.
- Goswami a Friscia 2010, s. 13–14.
- Benton 2015, s. 382.
- Kirkwood 2013, s. 10.
- BELLANI, Giovanni Giuseppe. Felines of the World: Discoveries in Taxonomic Classification and History. [s.l.]: Academic Press, 2019. DOI 10.1016/b978-0-12-816503-4.00001-5. Kapitola Order of carnivores (Carnivora). (anglicky)
- carnivore [online]. E. Britannica [cit. 2021-08-27]. Dostupné online. (anglicky)
- KRUUK, H. Carnivores. In: LEVIN, S. A. Encyclopedia of biodiversity. 1, A-C.. San Diego etc.: Academic Press, cop. 2001. DOI 10.1016/b0-12-226865-2/00043-2. S. 629–640. (anglicky)
- Vaughan, Ryan a Czaplawski 2015, s. 285.
- Turner 1997, s. 19–20.
- Kingdon 2013, s. 26.
- Berkovitz a Shellis 2018, s. 268–270.
- Nowak 2005, s. 6–7.
- Natural History Collections: Carnivores. www.nhc.ed.ac.uk [online]. The University of Edinburgh [cit. 2021-08-31]. Dostupné online. (anglicky)
- McDade 2003, s. 256–257.
- Antón 2013, s. 19, 104.
- McDade 2003, s. 409–410.
- Dungel a Gaisler 2002, s. 88, 90.
- Gaisler a Zima 2007, s. 620.
- Kingdon 2013, s. 36.
- Berkovitz a Shellis 2018, s. 268.
- Berkovitz a Shellis 2018, s. 296.
- Vaughan, Ryan a Czaplawski 2015, s. 284.
- ROČEK, Zbyněk. Obecná morfologie živočichů, Smysly [online]. Dostupné online.
- PEICHL, Leo; BEHRMANN, Günther; KRÖGER, Ronald H. H. For whales and seals the ocean is not blue: a visual pigment loss in marine mammals*: A visual pigment loss in marine mammals. European Journal of Neuroscience. 2001-04, roč. 13, čís. 8, s. 1520–1528. Dostupné online [cit. 2021-09-06]. DOI 10.1046/j.0953-816x.2001.01533.x. (anglicky)
- MYERS, Phil; POOR, Allison. Carnivora (carnivores). Animal Diversity Web [online]. [cit. 2021-09-03]. Dostupné online. (anglicky)
- Nowak 2005, s. 4–6.
- Vaughan, Ryan a Czaplawski 2015, s. 285–286.
- Kirkwood 2013, s. 19.
- Gaisler a Zima 2007, s. 623.
- Feldhamer 2015, s. 801.
- Anděra 1999, s. 7–8.
- McDade 2003, s. 257–260.
- Nowak 2005, s. 17–18.
- Nowak 2005, s. 13–14.
- Kingdon 2013, s. 143.
- Nowak 2005, s. 18–19.
- MACDONALD, D. W.; JOHNSON2, D. D. P. Patchwork planet: the resource dispersion hypothesis, society, and the ecology of life. Journal of Zoology. 2015, s. 76. Dostupné online. (anglicky)
- Castelló 2020, s. 51.
- Castelló 2020, s. 241.
- McDade 2003, s. 261–262.
- MCCANN, T. S.; FEDAK, M. A.; HARWOOD, J. Parental investment in southern elephant seals, Mirounga leonina. Behavioral Ecology and Sociobiology. 1989-08, roč. 25, čís. 2, s. 81–87. Dostupné online [cit. 2021-11-05]. ISSN 0340-5443. DOI 10.1007/BF00302924. (anglicky)
- Gittleman 1989, s. 76–77.
- Feldhamer 2015, s. 794–795.
- Anděra 1999, s. 72.
- Gittleman 1989, s. 57.
- Gittleman 1989, s. 58.
- Nowak 2005, s. 30.
- Gittleman 1989, s. 58–60.
- Kingdon 2013, s. 265.
- Nowak 2005, s. 166–167.
- Gittleman 1989, s. 73.
- Nowak 2005, s. 28–29.
- FROMMOLT, Karl-Heinz; GOLTSMAN, Mikhail E.; MACDONALD, David W. Barking foxes, Alopex lagopus: field experiments in individual recognition in a territorial mammal. Animal Behaviour. 2003-03-01, roč. 65, čís. 3, s. 509–518. Dostupné online [cit. 2021-11-04]. ISSN 0003-3472. DOI 10.1006/anbe.2003.2064. (anglicky)
- DUBZINSKI, K. M.; THOMAS, J. A.; GREGG, J. D. Communication in Marine Mammals. In: Perrin, W. F.; Würsig, B.; Thewissen, J. G. M. Encyclopedia of Marine Mammals. 2. vyd. San Diego (California) ; London: Academic Press, 2009. S. 266–267. (anglicky)
- RASA, O. Anne E. Dwarf Mongoose and Hornbill Mutualism in the Taru Desert, Kenya. Behavioral Ecology and Sociobiology. 1983, roč. 12, čís. 3, s. 181–190. Dostupné online [cit. 2021-11-04]. ISSN 0340-5443. (anglicky)
- Gittleman 1989, s. 18–19.
- HOLLIDAY, J. A.; STEPPAN, S. J. Evolution of hypercarnivory: the effect of specialization on morphological and taxonomic diversity. Paleobiology. 2004, roč. 30, čís. 1, s. 108–128 (109–110). Dostupné online. DOI 10.1666/0094-8373(2004)030<0108:EOHTEO>2.0.CO;2. (anglicky)
- Nowak 2005, s. 15–17.
- McDade 2003, s. 260–261.
- DEVLIN, H. Hard to bear: pandas poorly adapted for digesting bamboo, scientists find. the Guardian [online]. 2015-05-19 [cit. 2021-09-09]. Dostupné online. (anglicky)
- Feldhamer 2015, s. 795–796.
- KOLEKTIV AUTORŮ. Velký atlas živočichů jedinečný obraz života na Zemi. Bratislava: Príroda, 2005. 208 s. ISBN 80-07-01395-4. S. 64–65.
- Sloth bear. Smithsonian's National Zoo [online]. 2016-04-25 [cit. 2021-09-10]. Dostupné online. (anglicky)
- McDade 2003, s. 257.
- PETERSON, Ch. 25 years after returning to Yellowstone, wolves have helped stabilize the ecosystem. National Geographic [online]. 2020-07-10 [cit. 2021-09-13]. Dostupné online. (anglicky)
- DOMNING, D. Hydrodamalis gigas [online]. The IUCN Red List of Threatened Species 2016: e.T10303A43792683., 2016-04-04 [cit. 2021-12-27]. Dostupné online. DOI https://dx.doi.org/10.2305/IUCN.UK.2016-2.RLTS.T10303A43792683.en. (anglicky)
- GALBREATH, R. The tale of the lighthouse-keeper's cat: Discovery and extinction of the Stephens Island wren (Traversia lyalli). Notornis. 2004, roč. 51, čís. 4, s. 193–200 (200). Dostupné online [cit. 2021-09-13]. (anglicky)
- McDade 2003, s. 262–263.
- Mazák 1980, s. 86, 9 ff.
- RIEDMAN, M. The Pinnipeds: Seals, sea lions, and walruses. California: University of California Press, 1990. Dostupné online. ISBN 978-0-520-06497-3. S. xxi. (anglicky)
- JØN, A. Asbjørn. Dugongs and Mermaids, Selkies and Seals. S. 94–98. Australian Folklore [online]. 1998 [cit. 2021-03-12]. Roč. 13, s. 94–98. Dostupné online. (anglicky)
- Nowak 2005, s. 35.
- Hunter a Barrett 2018, s. 11–12.
- Nowak 2005, s. 36–37.
- Goodrich, J., Lynam, A., Miquelle, D., Wibisono, H., Kawanishi, K., Pattanavibool, A., Htun, S., Tempa, T., Karki, J., Jhala, Y. & Karanth, U. 2015. Panthera tigris. The IUCN Red List of Threatened Species 2015: e.T15955A50659951. https://dx.doi.org/10.2305/IUCN.UK.2015-2.RLTS.T15955A50659951.en. Downloaded on 11 November 2021.
- seal [online]. E. Britannica [cit. 2021-11-13]. Dostupné online. (anglicky)
- Doroff, A., Burdin, A. & Larson, S. 2021. Enhydra lutris. The IUCN Red List of Threatened Species 2021: e.T7750A164576728. https://dx.doi.org/10.2305/IUCN.UK.2021-3.RLTS.T7750A164576728.en. Accessed on 29 December 2021.
- Valqui, J. & Rheingantz, M.L. 2021. Lontra felina. The IUCN Red List of Threatened Species 2021: e.T12303A95970132. https://dx.doi.org/10.2305/IUCN.UK.2021-3.RLTS.T12303A95970132.en. Accessed on 29 December 2021.
- BREITENMOSER, Urs; BREITENMOSER-WÜRSTEN, Christine; BOITANI, Luigi. Assessing conservation status and units for conservation. In: BOITANI, Luigi; POWELL, Roger. Carnivore Ecology and Conservation. New York: Oxford University Press, 2012. S. 363. (anglicky)
- Nowak 2005, s. 40.
- Mammals [online]. IUCN SSC Groups, 2016-01-21 [cit. 2021-12-27]. Dostupné online. (anglicky)
- Nowak 2005, s. 32.
Literatura
- Česky
- ANDĚRA, Miloš, 1999. Savci (2): šelmy, luskouni, hrabáči, hlodavci. Praha: Albatros. (Svět zvířat).
- DUNGEL, Jan; GAISLER, Jiří, 2002. Atlas savců České a Slovenské republiky. Praha: Academia. 150 s. ISBN 80-200-1026-2. S. 88–117. [Přehled šelem ČR a SR s obecnou charakteristikou řádu].
- GAISLER, Jiří; ZIMA, Jan, 2007. Zoologie obratlovců. 2. vyd. Praha: Academia. ISBN 978-80-200-1484-9. S. 619–624.
- HERÁŇ, Ivan, 1978. Medvědi a pandy. Praha: Státní zemědělské nakladatelství. (Zvířata celého světa; sv. 4).
- HERÁŇ, Ivan, 1982. Kunovité šelmy. Praha: Státní zemědělské nakladatelství. (Zvířata celého světa; sv. 9).
- MAZÁK, Vratislav, 1980. Velké kočky a gepardi. Praha: Státní zemědělské nakladatelství. (Zvířata celého světa; sv. 7).
- Anglicky
- ANTÓN, Mauricio, 2013. Sabertooth. Indiana: Indiana University Press. 243 s. ISBN 80-7234-213-4. (anglicky)
- BENTON, Michael, 2015. Vertebrate palaeontology. 4. vyd. Chichester, West Sussex ; Hoboken: Blackwell Publishers. (anglicky)
- BERKOVITZ, Barry; SHELLIS, Peter, 2018. The Teeth of Mammalian Vertebrates. 1. vyd. London, United Kingdom ; San Diego, CA, United States: Academic Press, an imprint of Elsevier. ISBN 978-0-12-802818-6. DOI 10.1016/b978-0-12-802818-6.00015-6. S. 267–304. (anglicky)
- CASTELLÓ, J. R, 2020. Felids and Hyenas of the World: Wildcats, Panthers, Lynx, Pumas, Ocelots, Caracals, and Relatives. Princeton & Oxford: Princeton University Press. 280 s. ISBN 0691211868, ISBN 9780691211862. (anglicky)
- FELDHAMER, George A. & kol, 2015. Mammalogy – Adaptation, Diversity, Ecology. 4. vyd. Baltimore: Johns Hopkins University Press. Kapitola CHAPTER 17 – Carnivora. (anglicky)
- GITTLEMAN, John L, 1989. Carnivore Behavior, Ecology, and Evolution. Ithaca: Comstock Pub. Associates. ISBN 978-0-412-34360-5. (anglicky)
- GOSWAMI, Anjali; FRISCIA, Anthony, 2010. Carnivoran evolution : new views on phylogeny, form, and function. Cambridge ; New York: Cambridge University Press. ISBN 9780521515290, ISBN 9780521735865. (anglicky)
- HUNTER, Luke; BARRETT, Priscilla, 2018. A Field Guide to the Carnivores of the World. 2. vyd. London: Bloomsbury Wildlife. (anglicky)
- KEMP, Thomas Stainforth. The Origin and Evolution of Mammals. Oxford ; New York: Oxford University Press, 2005. (anglicky)
- KINGDON, Jonathan & kol, 2013. Mammals of Africa – Carnivores, Pangolins, Equids and Rhinoceroses. London ; New York: Bloomsbury Natural History. ISBN 978-1-4081-2255-6, ISBN 978-1-4081-8994-8. (anglicky)
- KIRKWOOD, Roger, 2013. Fur seals and sea lions. Collingwood, VIC: CSIRO Pub. ISBN 9780643109841. (anglicky)
- MCDADE, M. C. & kol, 2003. Grzimek’s Animal Life Encyclopedia – Volume 14 Mammals III. 2. vyd. Farmington Hills, MI: Gale. Kapitola Carnivora. (anglicky)
- NOWAK, Ronald M, 2005. Walker's Carnivores of the World. Baltimore: John Hopkins University Press. ISBN 0801880335, ISBN 0801880327. (anglicky)
- ROSE, Kenneth D, 2006. The Beginning of the Age of Mammals. Baltimore: The Johns Hopkins University Press. (anglicky)
- TURNER, Alan, 1997. The Big Cats and Their Fossil Relatives: An Illustrated Guide to Their Evolution and Natural History. New York: Columbia University Press. ISBN 978-0-231-10229-2. (anglicky)
- VAUGHAN, Terry A.; RYAN, James M.; CZAPLAWSKI, Nicholas J, 2015. Mammalogy. 6. vyd. Burlington, MA: Jones & Bartlett Learning. (anglicky)
Externí odkazy
- MYERS, Phil; POOR, Allison. Carnivora (carnivores). Animal Diversity Web [online]. [cit. 2021-09-03]. Dostupné online. (anglicky)
 Obrázky, zvuky či videa k tématu šelmy na Wikimedia Commons
Obrázky, zvuky či videa k tématu šelmy na Wikimedia Commons  Taxon Carnivora ve Wikidruzích
Taxon Carnivora ve Wikidruzích Slovníkové heslo šelma ve Wikislovníku
Slovníkové heslo šelma ve Wikislovníku