Ptáci
Ptáci (Aves) jsou třída teplokrevných obratlovců, pro kterou je charakteristická přítomnost peří, bezzubého zobáku, snášení skořápkových vajec, vysoká rychlost metabolismu, čtyřdílné srdce a pevná lehká kostra s častými srůsty kostí. Přední končetiny jsou přeměny v křídla (jedinou známou výjimku představují epyornisovití a moa). Celkem je dnes známo asi 10 400 druhů žijících ptáků (z toho přes 4083 druhů nepěvců a 5895 druhů pěvců)[2][3] a přes 2000 druhů fosilních.[4][5] Je ale možné, že skutečná biodiverzita současného ptactva je ještě mnohem vyšší a čítá asi 11 000 až 18 000 druhů.[6] Odhaduje se, že na Zemi žije 50 miliard jedinců ptáků.[7]
Stratigrafický výskyt: Svrchní jura až současnost, 160 až 0 milionů let[1] | |
|---|---|
 Koláž znázorňující diverzitu ptáků | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Podkmen | obratlovci (Vertebrata) |
| Nadtřída | čelistnatci (Gnathostomata) |
| Třída | ptáci (Aves) |
| Řády | |
viz Seznam řádů | |
| Některá data mohou pocházet z datové položky. | |
Ptáci představují teropodní dinosaury a obecněji diapsidy.[8][9] Dá se konstatovat, že jsou přežívající specializovanou skupinou teropodních dinosaurů (maniraptorů), která jako jediná přežila vymírání na konci křídy před 66 miliony let.[10][11]
Věda zabývající se touto skupinou živočichů se nazývá ornitologie.
Evoluce
Ptáci jsou pokládáni za potomky drobných teropodních dinosaurů, konkrétně maniraptorů (klad Pennaraptora a Avialae), z nichž se vyvinuli v období střední až svrchní jury, před více než 160 miliony let.[12][13] Dinosaury jako předky ptáků označil již německý anatom Carl Gegenbaur v roce 1863[14] a jedná se tedy o nejstarší přesně zformulovanou hypotézu o původu ptáků. O několik let později postuloval anatomickou podobnost teropodních dinosaurů a ptáků také britský přírodovědec Thomas Henry Huxley.[15] V posledních čtyřiceti letech je díky výzkumům Johna Ostroma respektována naprostou většinou paleontologů a mnoha ornitology.[16]
Evoluční původ ptáků
Evoluce ptáků prošla patrně třemi výraznými diverzifikačními skoky, jak ukazují biomolekulární rozbory. První se přitom odehrál asi před 170 až 160 miliony let (na přelomu střední a pozdní jury), druhý před 65 až 55 miliony let (po vyhynutí neptačích dinosaurů na přelomu křídy a paleocénu) a poslední v období eocénu, asi před 45 až 40 miliony let.[17]
První „praví“ ptáci se nepochybně objevili již v období druhohor, pravděpodobně pak v období střední až pozdní jury.[18] Určení nejstaršího známého ptáka závisí na definici jména Aves, která je sporná. Klad žijících ptáků, který za Aves označuje Jacques Gauthier a většina neontologů,[19] zřejmě pochází ze spodní křídy. Fosilní záznam z této doby je sporný, podle molekulárně biologické evidence však na konci křídy už existovalo 37 skupin žijících ptáků.[20] Většina paleontologů ale upřednostňuje zahrnout do skupiny Aves i taxony stojící mimo skupinu tvořenou všemi žijícími druhy a definovat jej jako Archaeopteryx, žijící ptáci, jejich poslední společný předek a všichni jeho potomci“.[21]
Pak se nejstarším ptákem stává sám pozdně jurský Archaeopteryx, žijící asi před 150 miliony let. Příbuznosti mezi ptáky a dinosaury nasvědčuje mnoho společných znaků, významným potvrzením této vývojové vazby byl především objev opeřených dinosaurů v Číně od roku 1996 (dnes je známo již 30 rodů opeřených dinosaurů). Od nástupu fylogenetické systematiky na začátku sedmdesátých let jsou ptáci považování nejen za potomky dinosaurů, ale přímo za jejich podskupinu.[22] Vědecká studie z roku 2018 dokazuje, že tělesné opeření a metabolismus se u ptačích předků zřejmě do značné míry vyvíjely společně. Maniraptorní dinosauři a primitivní jurští a křídoví ptáci (enantiornité) však měli zřejmě nižší bazální teplotu než současní ptáci.[23] Ptáci prošli vývojovou ztrátou zubů (edentulismem) již v době před asi 90 miliony let, jak ukazují biomolekulární výzkumy.[24]
Podle některých teorií se ptačí let vyvinul ze schopnosti plachtění na membranózních křídlech u nedinosauřích diapsidních plazů.[25] Ptačí let vznikal postupnými kroky, pravděpodobně přes stadium tzv. křídly asistovaného běhu po nakloněné rovině (WAIR).[26][27] Vývoj ptáků nicméně významně ovlivňovaly změny atmosférických podmínek a klimatu v průběhu druhohorní éry.[28] V současnosti již o přímé vývojové vazbě ptáků a dinosaurů pochybuje jen málokdo.[29]
Vznik moderních ptáků v období křídy
Jak ukázal například objev malého praptáka druhu Asteriornis maastrichtensis z nejpozdnější křídy Belgie (stáří 66,7 m. l.), „moderní“ ptáci se začali vyvíjet nejpozději koncem křídy, ještě v době nadvlády dinosaurů nad pevninami naší planety.[30] Někteří z nich již měli vyvinuté relativně pokročilé adaptace, například výrazně tvarované zobáky (tato schopnost se přitom dosud předpokládala pouze u moderních Neornithes). Takovým příkladem je madagaskarský taxon Falcatakely forsterae, formálně popsaný v listopadu roku 2020.[31]
Rozsáhlý srovnávací výzkum doložil, že relativní velikost mozku ptáků se začala výrazně zvyšovat u některých kladů (vývojových skupin) krátce po vymírání na konci křídy. Až do doby před 66 miliony let byla relativní velikost mozkovny praptáků a příbuzných neptačích dinosaurů podobná. Nejvyšší relativní objem mozkovny dnes nalezneme u papoušků a zejména krkavcovitých ptáků.[32][33][34] Jak také ukázal výzkum, morfologická evoluce ptačích lebek se po vyhynutí dinosaurů v době před 66 miliony let dramaticky zpomalila.[35]
Vznik ptáků je velmi dobře zdokumentován díky fosilnímu záznamu, ve kterém je patrný vývoj peří a změny kostry nutné pro přechod od pasivního k aktivnímu letu. Tento pohled má původ v popisu kostry menšího teropodního dinosaura z čeledi Dromaeosauridae jménem Deinonychus antirrhopus, který byl publikován roku 1969 americkým paleontologem Johnem Ostromem.[36] Určení sesterské skupiny ptáků opět závisí na definici jména Aves: v Gauthierově definici založené na žijících druzích to jsou Ichthyornis nebo Iaceornis (velmi podobní žijícím ptákům a s jediným nápadným primitivním znakem, který představují zuby v čelistech), v Chiappeho definici zase nejčastěji dinosauři ze skupiny Scansoriopterygidae nebo Deinonychosauria, která zahrnuje klady Troodontidae a Dromaeosauridae. (Do druhé z nich spadá i známý Microraptor, popsaný v roce 2000[37], Velociraptor a další.) Peří ale měli již méně odvození (ne však nutně starší) dinosauři, jako Sinosauropteryx nebo Dilong (vzdálený příbuzný legendárního druhu Tyrannosaurus rex). Zatímco znaky, jako dýchání usnadňované vzdušnými vaky[38], duté obratle, endotermie[39] nebo dokonce jednoduché vláknité peří[40] byly přítomny u většiny dinosaurů a někdy dokonce i u jiných skupin (pterosauři), další rysy, jako revertovaný palec na noze[41], alula, aktivní let[42] nebo karpometakarpus se nevyskytují ani u všech taxonů historicky pokládaných za ptáky. V takovéto situaci, kdy typické ptačí znaky přibývají postupně na velké časové škále, je těžké rozhodnout, co již může být považováno za ptáka a co ještě ne. Ztráta zubů u vývojově primitivních ptáků nebyla vázána na aktivní let – ptačí tělo totiž „odlečila“ jen nepatrně.[43]
Za prvního ptáka je především pro svůj historický význam často považován Archaeopteryx, jehož nález roku 1860 (popsán 1861) způsobil senzaci. Mezi další významné fosilní taxony patří čínský Confuciusornis, jeden z prvních ptáků se zkráceným ocasem a pygostylem, Ichthyornis, který vzdáleně připomíná rybáka se zuby, a dravý Hesperornis s nepneumatizovanými kostmi a redukcí křídel podobného dnešním potápkám. Velkým kladem, zahrnující jak mnoho fosilních druhů, tak i všechny žijící ptáky, je Ornithothoraces. U báze této skupiny se poprvé vyskytuje množství odvozených znaků, jako je křidélko (alula), karpometakarpus, tarzometatarzus, revertovaný palec, prsní kost s masivním hřebenem, do stran orientovaná jamka na lopatce pro skloubení s pažní kostí a aktivní let.[44] Přední končetina dinosauřích předků ptáků prošla výrazným vývojem a zásadními anatomickými změnami.[45] Ornitotoracini se dělí na tzv. protiptáky (Enantiornithes), kam patří např. Gobipteryx nebo Sinornis, a pravé ptáky (Euornithes), jejichž nejbazálnějšími zástupci je klad Hongshanornithidae a nelétavý Patagopteryx a kam patří i všichni moderní ptáci (Neornithes). Ty se u své báze rozdělují na skupiny Palaeognathae – zahrnující nelétavé běžce (kivi, emu, nandu, pštrosy, kasuáry a vyhynulé ptáky moa) a létavé tinamy – a Neognathae, zahrnující všechny ostatní ptáky. K rozdělení na paleognáty a neognáty došlo zřejmě ještě před koncem křídy. Systém neognátů je zatím značně nestálý, což je způsobeno velmi rychlou evoluční radiací. Za prokázané lze považovat další rozdělení na Galloanserae (drůbež), kam patří hrabaví a vrubozobí, a Neoaves. Vnitřní struktura Neoaves není dosud stabilní, některé studie však odhalily existenci dalších dvou obsáhlých podskupin, Metaves a Coronaves.[46] Množství shodných znaků lze vysledovat také při ontogenetickém vývoji lebky a zadních končetin ptáků a jejich dinosauřích předchůdců.[47][48]
Nejbližšími žijícími příbuznými ptáků jsou krokodýli – společně představují jediné žijící zástupce kladu Archosauria.[49]
Systematika
Všichni žijící ptáci spadají do několika desítek přirozených (monofyletických) skupin, jejichž složení a často i vnitřní příbuzenství je poměrně stabilní[50]. Tyto skupiny často odpovídají „čeledím“ nebo v několika případech „řádům“ klasické systematiky Wetmore'a[51]. Rovněž jsou známy nejzákladnější (nejhlubší) příbuzenské vztahy mezi těmito skupinami: nelétaví běžci (kivi, emu, nandu, pštrosi, kasuáři, vyhynulí ptáci moa) a tinamy dohromady tvoří klad Palaeognathae, který je sesterskou skupinou vůči všem zbývajícím žijícím ptákům – nazývaným Neognathae. Uvnitř neognátů je dobře rozeznatelný klad Galloanserae (drůbež), kam patří hrabaví (Galliformes) a vrubozobí (Anseriformes). Všichni neognáti mimo drůbež tvoří další přirozenou skupinu, Neoaves. Problematické tak zůstávají vztahy na střední úrovni, to je mezi jednotlivými „řády“ neoavianů.[50]
Z metodologického hlediska je dnešní ptačí systematika založená na kladistických analýzách genomů, (mitochondriálního i jaderného) nebo – v menší míře – morfologických znaků. Aplikace kladistiky na DNA vytlačila dřívější fenetickou techniku DNA-DNA hybridizace, která měří podobnost bez ohledu na to, zda je zděděná od společného předka. Na hybridizaci byly založeny velké analýzy 90. let, včetně té, kterou předložili Sibley & Ahlquist (1990).[52] Její závěry jsou dosud považovány za poměrně spolehlivé, pokud jde o seskupování jednotlivých druhů do větších celků, ne však, co se týče vzájemných vztahů mezi těmito většími skupinami.
Seznam řádů
Mezinárodní ornitologická unie rozeznává k roku 2021 následující řády[53] (české názvy doplněny podle Názvoslovná komise ČSO[54]):
- Ptáci (Aves)
- Běžci (Palaeognathae)
- Letci (Neognathae)
- Galloanserae
- Neoaves
- Brodiví a pelikáni
- Čápi
- Dlouhokřídlí
- Dravci
- Dropi
- Faetoni
- Gvačarové
- Hoacinové
- Krátkokřídlí
- Kukačky
- Kurolové
- Lelčíkové
- Lelkové
- Lelkouni
- Měkkozobí
- Mesitové
- Myšáci
- Papoušci
- Pěvci
- Plameňáci
- Potápky
- Potáplice
- Potuové
- Seriemy
- Slunatci a kaguové
- Sokoli
- Sovy
- Srostloprstí
- Stepokurové
- Svišťouni
- Šplhavci
- Terejové
- Trogoni
- Trubkonosí
- Tučňáci
- Turakové
- Zoborožci
Fylogeneze
Všechny moderní studie potvrdily rozdělení recentních ptáků do dvou hlavních skupin - Palaeognathae (česky nepřesně běžci) a Neognathae (letci). V rámci druhé skupiny je rovněž obecně přijímáno rozdělení na dva samostatné klady Galloanserae (zahrnující hrabavé a vrubozobé) a Neoaves (všechny ostatní skupiny).[55][5] Další dělení těchto skupin, především vnitřní uspořádání Neoaves, je předmětem probíhajících výzkumů.[56]
Jeden z posledně publikovaných kladogramů zohledňující moderní výzkumy DNA pochází o Heinera Kuhla et al., který v roce 2020 publikoval následující kladogram:[5]
| Aves |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
_white_background.jpg.webp)




_(Tauraco_persa).jpg.webp)
_(Antwerpener_Breiftaube).jpg.webp)

_white_background.jpg.webp)




Popis
Dnešní ptáci jsou charakterizováni peřím, čelistmi bez zubů, přeměněnými rohovinovou ramfotékou v zobák, vysokým stupněm metabolismu, dýcháním skrze vzdušné vaky, pronikající do postkraniální kostry, a kostrou s výskytem mnoha srostlých elementů (pygostyl, karpometakarpus, tarzometatarzus a tibiotarzus). Ptáci mají přední končetiny přeměněny v křídla umožňující let, přestože běžci a někteří další, zejména endemické ostrovní druhy, tuto schopnost k letu ztratily. Létavé ptáky dále charakterizuje kost prsní s kýlem pro úpon hrudních svalů, jehož velikost přímo odpovídá letovým schopnostem. Kostru mají lehkou, kosti jsou duté.
Obecně jsou zástupci velké většiny ptáků pouze malými obratlovci s hmotností zhruba do 1 kilogramu. V geologické minulosti ale existovaly často velké formy nelétavých ptáků s hmotností nad 100 kilogramů. Fosilie těchto fosilních až (sub)recentních zástupců známe například z Jižní Ameriky, Madagaskaru, Austrálie i Eurasie.[58]
Rozměry
Ptáci dosahují různých velikostí. Rozdíly ve velikosti ptáků nejsou tak velké jako u plazů nebo savců. Největší ptáci převyšují svojí hmotností ty nejmenší jen asi 45tisíckrát (létavé formy jen 10tisíckrát), zatímco například u savců je tento vztah v poměru 1:60 milionům. Za nejmenší ptáky světa jsou považováni kolibříci (hmotnost 1,5–2,5 g) a pěvci. Zpravidla za vůbec nejmenšího se považuje kubánský druh kalypta nejmenší (Mellisuga helenae), který i s ocasem měří 6 cm a váží 1,5 gramu, takže je nejmenším ptákem.[59] V Evropě jsou nejmenší králíčci (Regulus spp.), kteří mají hmotnost 4,5–7 g. Největší z žijících ptáků je pštros dvouprstý (2,5 m výšky, hmotnost až 150 kg). Z létavých pak někteří albatrosi (rozpětí křídel až 3,5 m), kondor velký (rozpětí 3 m) nebo orel mořský (rozpětí až 2,5 m). Ještě větší byly některé vymřelé druhy. Novozélandští ptáci moa (Dinornithiformes) měřili na výšku až 3,6 m a vážili až 250 kg. Madagaskarští běžci řádu Aepyornithiformes byli nižší (3 m), ale těžší (500 kg) a snášeli také vůbec největší vejce, velká 25 × 34 cm a těžká 10 kg (odpovídá 7 pštrosím vejcím). Zřejmě největším známým druhem ptáka byl nelétavý madagaskarský Vorombe titan, jehož hmotnost dosahovala pravděpodobně až 800 kilogramů.[60][61] Z létavých druhů byl zřejmě největší Argentavis magnificens, který vážil 60 až 110 kg a v rozpětí křídel měřil přes 6 metrů.[62] Ještě větší rozpětí křídel mohl mít rod Pelagornis, u nějž činilo asi 6 až 7 metrů.
Také druhohorní formy praptáků však dosahovaly miniaturních rozměrů. Například druh Oculudentavis khaungraae z myanmarského jantaru (formálně popsaný roku 2020) měl lebku dlouhou jen 14 mm a celkovou délku těla zhruba do 9 centimetrů, hmotnost tohoto malého teropoda dosahovala pouhých několika gramů.[63]
Největší formy ptáků známe z některých období pravěku či ještě nedávného recentu. Úplně největším známým druhem byl zřejmě před několika tisíciletími žijící druh „sloního ptáka“ Vorombe titan z Madagaskaru (s odhadovanou hmotností až kolem 800 kilogramů)[64], zatímco největším dosud známým dravým ptákem byl nejspíš Kelenken guillermoi (hmotnost kolem 250 kg, výška přes 3 metry) z miocénu Argentiny.[65]
Vnější vzhled současných ptáků je velmi podobný. Liší se pouze velikostí těla, tvarem a velikostí zobáku, v délce krku a typu nohy. Většina druhů ptáků patří mezi suchozemské obratlovce se schopností aktivního letu.
Rozšíření

Ptáci se rozmnožují na všech sedmi kontinentech, přičemž největší rozmanitosti dosahují v tropických oblastech; to může být způsobeno buď vyšší rychlostí speciace v tropech nebo vyšší rychlostí vymírání ve vyšších zeměpisných šířkách. Jsou schopni žít a nacházet potravu na většině lokalit naší planety, na jihu např. sněžní buřňáci hnízdí v některých koloniích až 440 km hluboko ve vnitrozemí Antarktidy. Několik příbuzných druhů se adaptovalo na život na světových oceánech – s některými druhy mořských ptáků se na pobřeží setkáme pouze v době rozmnožování a některé druhy tučňáků se potápějí do rekordních hloubek okolo 300 metrů. Mnoho druhů kolonizovalo oblasti, do kterých byly vysazeny lidmi. Některá z těchto zavádění byla dobře uvážena: např. bažant obecný byl vysazován po celém světě jako lovná pernatá zvěř. Někteří ptáci jsou kolonizátoři náhodní, jako např. papoušek mniší, který po útěcích z klecí vytvořil kolonie v některých severoamerických městech. Některé jiné druhy, jako např. volavka rusohlavá, čimango žlutavý a kakadu růžový se rozšířily hluboko za hranice svého původního výskytu díky tomu, jak jim zemědělská činnost člověka vytváří nové vhodné lokality.
Anatomie

Ve srovnání s ostatními obratlovci mají ptáci uspořádání těla vykazující mnoho neobvyklých přizpůsobení, většinou k usnadnění letu.
Kostra je složena z velmi lehkých kostí. Kosti jsou duté a vyplněné vzduchem, přičemž dutiny jsou spojeny s dýchacím ústrojím. Tyto duté kosti jsou přitom velmi pevné. Lebeční kosti jsou spojené a bez zřetelných švů. Očnice jsou mohutné a oddělené kostní přepážkou. Páteř je rozdělena na krční, hrudní, křížový a ocasní oddíl s ohebnými krčními obratli, jejichž počet je vysoce proměnlivý. Pohyb je omezen u obratlů počínaje posledními hrudními obratli a není možný u dalších obratlů. Kost kyčelní je srostlá po celé délce s páteří. Žebra jsou zploštěná a jsou upnuta k mohutné deskovité prsní kosti, k níž jsou připojeny létací svaly. Pouze nelétaví ptáci mají prsní kost poměrně malou a bez hřebene. Přední končetiny jsou přeměněny v křídla. Výzkum ukázal, že ptačí „šupiny“ na nohách nemají stejný evoluční základ jako plazí šupiny a vznikly spíše jako morfologické odvozeniny od peří.[66]
Podobně jako plazi ani ptáci nemočí. Ledviny extrahují z krevního řečiště dusíkatý odpad, ten však místo vylučování jako močoviny rozpuštěné v moči je vylučován ve formě kyseliny močové. Ptáci také vyměšují spíše kreatin než kreatinin, jako je tomu u savců. Kyselina močová má velmi nízkou rozpustnost ve vodě, takže po zbavení se zbytku vody se objevuje jako bílý povlak. Od každé ledviny vede močovod, který ústí do středního oddílu kloaky. Právě v kloace se moč zbavuje vody a při průchodu trusu je stírána a s trusem vylučována. Na trusu se moč jeví jako bílý povlak. Kloaka je společným vývodem trávicí, vylučovací a pohlavní soustavy. Navíc mnoho druhů ptáků nestravitelné zbytky potravy vyvrhuje v chuchvalcích (tzv. vývržky) zpět jícnem a ústní dutinou.
Ptáci mají jedno z nejkomplexnějších dýchacích ústrojí ze všech zvířecích skupin. Když se pták nadechuje, 75 % objemu čerstvého vzduchu obtéká plíce a vtéká rovnou do vzdušných vaků, probíhajících od plic a spojených s dutinami v kostech, které plní vzduchem. Zbylých 25 % vzduchu jde přímo do plic. Když pták vydechuje, je pro výdech používán vzduch z plic a současně se do plic dostává čerstvý vzduch ze vzdušných vaků. Ptačí plíce tak přijímají čerstvý vzduch jak při nádechu, tak i při výdechu. Ptačí zpěv vzniká v orgánu zvaném syrinx, svalnaté komoře s několika bubínkovými blánami, který se nachází v dolní části průdušnice. Ptačí srdce má čtyři oddíly, z levé, silně svalnaté komory, vystupuje pravý aortální oblouk a dává vzniknout systémové aortě (na rozdíl od savců, kde je použit levý oblouk). Zadní dutá žíla přijímá krev z končetin přes vstupní ledvinový systém. Ptáci, na rozdíl od savců, mají jádrové červené krvinky, tj. červené krvinky, které mají jádro.
Zažívací soustava ptáků je jedinečná, mají hrdelní vak nebo vole pro uskladnění potravy a dva žaludky, z nichž ve druhém jsou spolykané kamínky, které vzhledem k absenci zubů pomáhají ptákům drtit potravu. Trávicí soustava dokáže zajistit rychlé strávení potravy a je přizpůsobena letu. Někteří stěhovaví ptáci mají přidanou schopnost snížit obsah části střev před migrací.
Nervová soustava u ptáků je v rozsahu přiměřeném velikosti ptáka. Nejvyvinutější částí mozku je mozeček, který koordinuje pohyb a ovládá funkce souvisící s letem, zatím co koncový mozek kontroluje prvky chování, plavání, páření a stavbu hnízd. Zrakový systém je obvykle u ptáků vysoce vyvinut. Oko ptáků je v základní stavbě shodné s okem savců. Ptáci s očima na stranách hlavy mají široké zorné pole, zatímco ptáci s očima na čelní straně hlavy, jako sovy, mají binokulární vidění a mohou odhadovat hloubku pole. Vodní ptáci mají zvláštní pružné čočky, které jim umožňují vidění na vzduchu i pod vodou. Některé druhy mají také dvojitou foveu. Ptáci jsou tetrachromatičtí díky přítomnosti očních buněk citlivých na ultrafialové záření, stejně jako na zelenou, modrou a červenou barvu. To jim umožňuje vnímat ultrafialové záření, což se využívá při krmení a námluvách. Mnoho ptáků má totiž na peří vzory viditelné v ultrafialovém světle a tudíž nerozeznatelné lidským okem. Ultrafialové světlo se využívá také při hledání potravy; např. poštolky při pátrání po kořisti často na zemi sledují močovou stopu hlodavců, která vyzařuje UV záření. Vnější ochranu oka zajišťují svrchní a spodní víčka, která však nejsou tak pohyblivá jako u savců, takže oko je čištěno jemnou blankou, umístěnou ve vnitřním koutku oka, tzv. mžurkou, která může oko překrýt směrem napříč. Mžurka také kryje oko a slouží tak jako kontaktní čočka u mnoha vodních ptáků. Když ptáci spí, jsou spodní víčka zvednutá. Ptačí sítnice má pro rozvod krve ve sklivci bohatě prokrvený výrůstek zvaný hřebínek. Ptačímu zevnímu zvukovodu chybí vnější ušní boltec, ale je překryt peřím, i když u některých druhů (např. kalous, výr, výreček) toto peří trochu uši připomíná. Ve vnitřním uchu se nalézá hlemýžď, který ale na rozdíl od savců není spirálovitý. Kvůli velmi dobře vyvinutému zraku se dříve předpokládalo, že ptáci jsou buď zcela anosmičtí (bez čichu) anebo cítí jen velice málo. Na základě rozboru mnoha současných studií se však ukazuje, že ptáci využívají čich v každodenním životě jako nástroj při hledání potravy, při hledání ideálního partnera pro reprodukci anebo při navigaci a orientaci v prostoru.[67]
Někteří ptáci také používají proti dravcům chemickou obranu. Někteří trubkonosí mohou vyvrhnout proti útočníkovi nepříjemný olejovitý výměšek z jícnu, zatímco některé druhy pěvců z rodu Pitohui, žijících na Nové Guineji, mají jedovatou látku homobatrachotoxin obsaženou nejen v těle, ale hlavně v kůži a peří.
Při pohybu po zemi využívají ptáci svých anatomických předností, jako je efektivní dýchací soustava, pevné a lehké kosti a relativně silné svaly na horních částech nohou, díky kterým dokážou některé druhy rychle běhat či skákat.[68]
Peří

Jednou z vlastností, která odlišuje ptáky od ostatních žijících skupin živočichů (kromě dinosaurů), je pokrytí kůže peřím. Peří je epidermální kožní porost, jako např. šupiny u plazů nebo srst u savců, který má u ptáků mnoho různorodých funkcí: plní termoregulační funkci tím, že tepelně izoluje tělo ptáka za chladného počasí nebo od vody, peří je nezbytné pro ptačí let a je také používáno při toku, maskování a signalizaci. Ptačí peří, vzhledem k tomu, že slouží různým účelům, není pouze jednoho typu. Peří potřebuje pravidelnou údržbu a ptáci tak provádí jeho čištění a rovnání každý den. Používají k tomu zobák, pomocí kterého zbavují peří cizích částeček a roztírají si po peří olejovité výměšky z kostrční žlázy, což zajistí, že se peří nepromáčí, udržuje mu pružnost, působí jako antibakteriální prostředek potlačující růst baktérií snižujících kvalitu peří. Dalším způsobem péče o peří je používání kyseliny mravenčí, která pomáhá z peří zahánět parazity.
Peří se souvislým praporem vytvářející vzhled ptačího těla se také nazývá krycí nebo konturové peří. Toto peří má zřetelný osten i brk a patří k němu většina peří rostoucího na těle. Krátké a měkké brky s navzájem nespojenými paprsky a větvičkami jsou znaky prachového peří. To tvoří první chmýřitý šat mladých ptáků a také obal pod krycím peřím ptáků dospělých. Druh peří tak může kolísat podle stáří jedince, jeho sociálního postavení, poškození peří podle postavení ve skupině, ale hlavně se jedinci liší v opeření podle pohlaví. Peří ptákům pravidelně vypadává vytlačováním starých per z kůže pery novými. Pelichání peří u většiny druhů ptáků probíhá jednou do roka, ale jsou známy druhy, u kterých probíhá výměna peří dvakrát za rok a oproti tomu velcí dravci mohou pelichat jednou za dva až tři roky. Kachnám a husám pelichá veškeré peří najednou, takže nejsou schopny zhruba po dobu jednoho měsíce letu. Různé ptačí druhy mají odlišné vzory a strategie línání. U některých ptáků dochází k pelichání od vnějších per k perům vnitřním, u jiných je tomu naopak a někteří ptáci ztrácejí veškeré peří najednou. První způsob nebo také centripetální pelichání jako termín pro pelichání rejdovacích per je možno vidět např. u bažantovitých. Druhé, nebo také centrifugální pelichání, je možné vidět např. u rejdovacích per datlů a šoupálků, přičemž střední pár peří vypadává jako poslední, aby bylo těmto druhům umožněno šplhání po stromech. Základní vzor způsobu pelichání je možné sledovat na vrabci, kdy jako první je nahrazováno vnější peří, poté peří vnitřní a u ocasních per se postupuje ze středu ke krajům.
Peří neroste rovnoměrně po celé kůži, ale rozrůstá se na zvláštních plochách nebo pruzích, které se nazývají pernice (latinsky pterylae). Distribuční vzorky těchto plošek nebo proužků jsou využívány v taxonomii a systematice. Před hnízděním, u samic většiny ptačích druhů, vzniká v místě styku jejich těla s vejci takzvaná hnízdní nažina, tedy místo bez peří. Ta bývá buď zcela holá, či jenom řídce porostlá prachovým peřím a je hodně prokrvená, aby byl při sezení na vejcích usnadněn transfer tepla.
Biologie a chování
Mnoho druhů ptáků podniká každoročně dlouhé migrační cesty a velké množství ptáků absolvuje také krátké nepravidelné tahy. Ptáci jsou sociální živočichové a komunikují pomocí vizuálních signálů, voláním a zpěvem, účastní se sociální činnosti včetně společného lovu, rozmnožování, shromažďování a napadání predátorů. Ptáci jsou převážně společensky monogamní s tím, že u některých druhů se setkáváme s krátkodobým nebo řidčeji i dlouhodobým životem v párech; jiné druhy jsou polygamní nebo polyandrické. Ptáci podobně jako obojživelníci a plazi snášejí vejce, která obvykle ukládají do hnízd, sedí na vejcích a zahřívají je až do doby vylíhnutí mláďat. Tělesná teplota ptáků se pohybuje mezi 38 až 42 stupni Celsia.[69] Výzkum ukazuje, že většina ptačích druhů v různých geografických prostředích a ekosystémech volí své vzorce chování a převažující tělesný postoj v závislosti na možnostech efektivního udržení svojí stálé tělesné teploty.[70]
Chování ptáků je řízeno instinkty. Nejvíce ptáků je aktivních ve dne, ale mnohé druhy ptáků, jako např. sovy či lelci, jsou aktivní v noci. Někteří ptáci mohou využívat i soumrak a existují i ptáci žijící na mořském pobřeží, kteří vyrážejí na lov potravy v době vhodného přílivu, lhostejno jestli ve dne, nebo v noci.
Inteligence
U mnoha ptáků (zejména papoušků a krkavcovitých) byla pozorována schopnost řešení problémů, tvorby nástrojů pro obstarávání potravy a mnoho dalších schopností, vyžadujících poměrně vysokou inteligenci a abstrakci. U některých ptáků je pravděpodobně inteligence tak vysoká, že ji překonává již pouze inteligence vyšších primátů.[71] Nové studie potvrzují, že ptáci jsou schopni stejně složitých myšlenkových pochodů, jako vyspělí savci.[72] Vývoj ptačího mozku má množství specifických vzorců, které se neobjevují při vývoji savčího mozku.[73]
Potrava

Ptáci se živí různou potravou, zahrnující nektar, ovoce, rostliny, semena, zdechliny, drobné bezobratlé živočichy i obratlovce včetně ptáků a savců. Protože ptáci nemají zuby, je jejich zažívací soustava speciálně přizpůsobena k přijímání celých, nepřežvýkaných kousků potravy. Bezzubý ptačí zobák se postupným vývojem formoval z ozubených čelistí ptačích předků – maniraptorních teropodních dinosaurů.[74]
Ptáci používají různé strategie vyhledávání potravy. Pro některé druhy je charakteristický sběr či lov bezobratlých včetně hmyzu, pro jiné sběr semen (primárně semenožraví ptáci). Pěvci při lovu letícího hmyzu často využívají taktiku výpadu z větve. Ptáci živící se nektarem, jako jsou kolibříci, papoušci lori, strdimilovití, kystráčkovití a někteří další, mají usnadněn sběr potravy díky jazýčku přizpůsobenému do podoby kartáčku; v mnoha případech mají zobák uzpůsobený pro určitý typ květin. Sondování zobákem při hledání bezobratlých používají kiviové a bahňáci s dlouhými zobáky; v případě bahňáků jsou délka zobáků a metody krmení spojeny s „potravní nikou“ (vše, co na daném území může danému organismu sloužit jako potrava). Stíhání střemhlavým letem je vlastní sokolům a jestřábům ve vzduchu, ve vodě podobný styl lovu používají např. potáplice, poláci a tučňáci. Lov skokem do vody provádí terejovití, ledňáčkovití a rybáci. Tři druhy buřňáků, plameňáci a některé kachny získávají potravu procezováním. Husy a kachny především spásají trávu. Některé druhy provozují kleptoparasitismus čili kradou potravu ostatním ptákům – fregatky, racci a chaluhy. Kleptoparasitismus není hlavním zdrojem obživy žádného druhu ptáků, ale je doplňkovým zdrojem k potravě získávané lovem; (studie fregatek páskovaných kradoucích potravu terejům maskovým odhadují, že fregatky tak mohou získat maximálně 40 % potravy, kterou potřebují, ale v průměru je toto číslo ještě nižší – činí jen 5 %). Konečně někteří ptáci, jako jsou supové a rackové, jsou mrchožrouti. Někteří ptáci mají při získávání potravy několik strategií nebo se živí různou potravou, zatímco jiní jsou potravní specialisté nebo využívají jedinou strategii při získávání potravy.
Let
Schopnost letu je charakteristická pro většinu ptáků a odlišuje je tak od ostatních skupin obratlovců s výjimkou netopýrů, kaloňů a vyhynulých pterosaurů.[75] Tento pro převažující množství druhů hlavní prostředek pohybu slouží při rozmnožování, lovu potravy a útěku před predátory. Ptáci jsou k letu přizpůsobeni různě; mají například lehkou kostru, dva velké prsní svaly zajišťující pohyb křídel dolů (tvoří 16 % hmotnosti ptáka), svaly podklíční zvedající křídla a přední končetiny přeměněné v křídla. Především velikost a tvar křídla určuje u jednotlivých druhů typ letu, který využívají především. Mnoho ptáků kombinuje tzv. veslovací let s méně náročným klouzavým letem nebo plachtěním. Okolo 60 druhů existujících ptačích druhů není letu schopno a podobně nebyli schopni letu i někteří vyhynulí ptáci. Tyto druhy neschopné letu je možno často nalézt na izolovaných souostrovích, pravděpodobně následkem nedostatku pozemních predátorů a omezených zdrojů, jejichž důsledkem je ztráta schopností letu. Tučňáci, kteří také nejsou schopni letu, používají křídla pro veslování při potápění pod vodou, podobně jako alky, buřňáci či skorci.
Základním způsobem pohybu ptáků, až na určité výjimky, je aktivní let. Ptáci nejčastěji používají mávavý let, kdy rytmicky zvedají a spouštějí křídla, přičemž rychlost a výšku letu regulují počtem pohybů a nakloněním křídel. Dalším způsobem letu zejména u velkých ptáků je plachtění, kdy se křídla prakticky nepohybují a ptáci využívají jen nosné síly vzdušných proudů. Další formou letu je tzv. třepotavý let, při němž se rychlým máváním křídel pták zastaví ve vzduchu na určitém místě (poštolka, rybák obecný). Zcela odlišným typem letu i z energetického hlediska je vířivý let kolibříků, umožňovaný zvýšenou frekvencí pohybu křídel.
Rychlost letu ptáků jen zřídka překonává hranici 100 km/h, přičemž k nejrychlejším patří husy, kachny, rorýsi aj. Drobní pěvci létají zpravidla rychlostí 30–50 km·h−1, poštovní holubi dosahují rychlosti až 80 km·h−1. Dobří letci jsou však schopni za určitých podmínek (např. pronásledování kořisti) krátkodobě vyvinout rychlost až 350 km·h−1. Nejvyšší změřená rychlost sokola stěhovavého při střemhlavém letu činila 389 km/h.[76]
Migrace
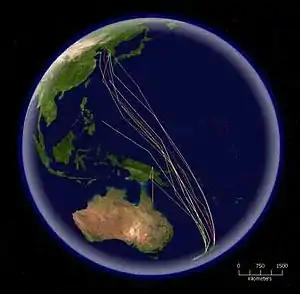
Mnohé ptačí druhy migrují, aby využily globální rozdíly sezónních teplot k optimalizaci dostupnosti zdrojů potravy a hnízdních lokalit. Tyto migrace jsou u různých skupin proměnlivé. Mnoho vnitrozemských, pobřežních či vodních ptáků zahajuje každoročně dalekou migraci, obvykle v závislosti na měnící se délce denního světla, stejně jako na povětrnostních podmínkách. Migrace jsou charakterizovány hnízdním obdobím v lokalitách mírného pásu nebo arktické či antarktické oblasti a tzv. zimovišti v oblastech tropů nebo na opačné straně polokoule. Dříve než zahájí migraci, zvýší ptáci podstatně své tukové zásoby a redukují velikost některých svých orgánů. Migrace je energeticky velice náročná, zvláště pokud ptáci překonávají bez doplňování potravy pouště nebo oceány. Vnitrozemští ptáci absolvují lety dlouhé až 2500 km a bahňáci až 4000 km. Břehouš rudý absolvuje bez zastávky tah o délce až 10 200 km. Mořští ptáci také absolvují dlouhé migrační cesty, nejdelší cestu podniká např. buřňák temný, který hnízdí na Novém Zélandu a v Chile, zatímco severské léto tráví krmením v Severním Pacifiku poblíž Japonska, Aljašky a Kalifornie, což znamená okružní cestu o délce přibližně 64 000 km. Jiní mořští ptáci se po vyhnízdění rozptylují, toulají po širokém okolí, ale nemají pevně zažité stěhovací cesty. Albatrosi, hnízdící v jižních mořích, často absolvují cirkumpolární cesty mezi hnízdními obdobími.
Ptáci provádějí i jiné způsoby migrace. Některé druhy se pouštějí do kratších cest, jen pokud je toho zapotřebí, aby se vyhnuly špatnému počasí, nebo kvůli získání potravy. Mezi ně patří i tzv. invazní druhy, které se mohou v některých letech vyskytovat zcela běžně a v jiných letech se naopak vůbec neobjevují. Tento typ migrace je obvykle spojen s dostupností potravy. Mezi takovéto migrující druhy jsou zařazovány např. severské pěnkavy, arktické sovy a brkoslavové. Ptáci také mohou cestovat na kratší vzdálenosti přesahující jejich teritoriální území s jednotlivci migrujícími na větší vzdálenosti; jiní provádějí pouze částečnou migraci, tzn. že migruje část populace, obvykle samice a subdominantní samci. Částečná migrace může tvořit velké procento migračního chování ptáků v některých regionech; v Austrálii průzkumy zjistily, že 44 % studovaných nezpěvavých ptáků a 32 % pěvců byli částečně stěhovaví. Výšková migrace je příkladem stěhování ptáků na krátké vzdálenosti, kdy období páření a výchovy mláďat probíhá ve větších nadmořských výškách, a když pominou příznivé podmínky, stěhují se do nižší polohy. Nejčastějším impulsem migrace jsou teplotní změny a obvykle je také zahájena ve chvíli, kdy se teritorium začne stávat nehostinným také v důsledku nedostatku potravy. Některé druhy mohou také kočovat, nemají žádné pevně vymezené teritorium a přesunují se v závislosti na počasí a dostupnosti potravy. Papoušci např. žijí rozptýleně, kočovně nebo pořádají nepravidelné a malé migrační cesty.
Schopnost ptáků vrátit se přes obrovské vzdálenosti přesně do určité lokality je již dlouho známa; při experimentu provedeném roku 1950 s buřňákem severním se jedinec vypuštěný v Bostonu vrátil do své kolonie na ostrovu Skomer u Walesu během 13 dnů; překonal vzdálenost 5150 km. Ptáci užívají k navigaci během migrace různé metody. Při denní migraci se k navigaci využívá slunce, v noci stelární kompas. Ptáci, kteří se orientují podle slunce, využívají pro určení směru tahu své vnitřní hodiny, pomocí nichž jsou schopni pudově určit odklon slunce a tím i světové strany. Orientace podle stelárního kompasu závisí na poloze souhvězdí okolo Polárky. Ta je u některých druhů posílena schopností magnetorecepce vnímat zemský geomagnetismus díky specializovaným fotoreceptorům.[77]
Komunikace
Ptáci komunikují hlavně používáním vizuálních a zvukových signálů. Tyto signály mohou být mezidruhové nebo vnitrodruhové.

Vizuální komunikace ptáků má více funkcí a je dána peřím a chováním. Peří může být používáno pro ocenění a potvrzení sociální dominance, zobrazuje připravenost k páření a dokonce může vyjadřovat hrozbu, jako je tomu např. u slunatce, který tak napodobuje větší dravce. Takovéto zobrazení na peří je užíváno k odrazení potenciálních predátorů, jako jsou např. krahujci, a ochraně mláďat. Variace v peří také umožňují identifikaci jedinců, zvláště mezi druhy.
Vizuální komunikace zahrnuje také rituální předvádění, signalizuje útok nebo podřízenost, slouží k formování párových vazeb. Toto ritualizované chování je tvořeno nesignalizovanými akcemi jako urovnánvání peří, časově rozlišené pozice pér, klování aj. Nejvíce propracované ukázky se objevují během námluv, jako např. při tanci v toku u albatrosů, kde úspěšné vytvoření celoživotního párového pouta vyžaduje, aby partneři praktikovali tento jedinečný tanec, a u rajek, kde úspěch samců při námluvách závisí na peří a kvalitě jeho předvádění. Samci ptáků mohou svou zdatnost demonstrovat také např. stavbou hnízda; samice snovačů, jako např. snovač asijský, si vybírají partnery podle kvality hnízda, zatímco samičky lemčíků přitahují konstrukce tzv. altánků, které samci staví a zdobí různými barevnými objekty.
Kromě vizuální komunikace jsou ptáci proslulí svými zvukovými schopnostmi. Volání a u některých druhů i zpěv jsou hlavními zvukovými komunikačními prostředky. Někteří ptáci využívají i mechanických zvuků, což může být rozeznění per pomocí proudu vzduchu (např. samci bekasíny otavní), teritoriální bubnování (datli) či používání nástrojů k tlučení (kakadu arový. Ptačí volání a písně mohou být složité. Zvuk bývá vytvářen v syrinxu umístěném na průdušnici v místě rozdvojení na průdušky, jehož obě strany některé druhy umí ovládat odděleně, čímž je možný v jednom čase vznik dvou různých písní najednou.[78][79]
Volání jsou používána pro různé cíle, několik jich může být spojeno do jedné písně. Jsou využívána při hledání partnera, jindy k jeho vábení, pomáhá k identifikaci potenciálních partnerů nebo ve společných skupinách (často v kombinaci s vizuální komunikací). Mohou předávat informace o kvalitě samců a pomoci při výběru samic. Hlas je používán i k vymezení a udržování teritoria. Volání může být používáno také k rozpoznávání jedinců, pomáhá rodičům nalézt mláďata v koloniích nebo dospělým shledat se s partnery na počátku období rozmnožování. Často se volání týká také varování dalších ptáků před potenciálními predátory; volání tohoto typu mohou být detailní a mohou vyjadřovat specifickou informaci o povaze hrozby.
Shlukování

Zatímco někteří ptáci jsou teritoriální nebo žijí v malých rodinných skupinách, jiní ptáci vytvářejí velká neteritoriální hejna. Výhody seskupování do hejn jsou různé a hejna mohou být tvořena výslovně pro specifické účely. Shlukování má také své nevýhody, zvláště společensky níže postavení ptáci, kteří jsou v područí více dominantních ptáků, musí obětovat krmnou efektivitu v hejnu za účelem zisku jiných výhod. Hlavními výhodami je větší bezpečí v hejnu a zvýšená krmná efektivita. Ochrana proti dravcům je zvláště důležitá v uzavřených lokalitách, jako je např. les, kde predátoři často útočí ze zálohy a včasné varování díky pozorování více jedinců tak nabývá na důležitosti. To vede i k vytváření druhově smíšených hejn. Mnohodruhová hejna se obvykle skládají z malých skupin více druhů; nevýhodou je konkurence při vyhledávání potravy. Ptáci se sdružují i s jinými živočichy; mořští ptáci lovící ryby vrháním se do moře z výšky se spojují s delfíny a tuňáky, kteří rybí hejna vyhánějí vzhůru k hladině; vzájemný vztah se také vyvinul např. mezi mangustou jižní a zoborožci: zoborožec vyhledává mangustu kvůli společnému hrabání a vzájemnému varování před predátory. Podobně je známé partnerství medozvěstek s medojedy, paviány, jezevci nebo lidmi při vyhledávání včelích příbytků, kdy je k nim dovede a po vybrání medu si sesbírá svůj díl (vosk, včely, larvy).
Odpočinek a hřadování
Vysoká rychlost metabolismu ptáků během aktivní části dne je doplňována odpočinkem ve zbylé části dne. Spící ptáci často používají typ spánku známý jako ostražitý spánek, při němž jsou periody odpočinku střídány s rychlým otevíráním očí, což zvyšuje citlivost ptáků na vyrušení a umožňuje jim rychlý únik před případnou hrozbou. Přestože experimentálně nebylo nikdy potvrzeno, že rorýsi mohou spát za letu, existují náznaky, že rorýsi využívají určitých druhů spánku, jež mohou probíhat dokonce za letu.
Ptáci nemají potní žlázy. Proto se ochlazují skrýváním ve stínu, stáním ve vodě, zrychleným dýcháním s otevřeným zobákem, zvětšováním plochy těla roztahováním křídel, vibrováním hrdelního laloku nebo močením na nohy (ochlazování vypařující se tekutinou).
Mnoho spících ptáků ohýbá ve spánku krk nad záda a zastrkává hlavu do peří na zádech, jiní skryjí zobáky v peří na hrudi. Mnoho ptáků také odpočívá pouze na jedné noze a někteří z nich druhou nohu schovávají do peří, zvláště za chladného počasí. Běžný je také společný hřad ptáků, kterým se snižuje ztráta tělesného tepla a zmenšují rizika spojená s napadením predátory. Hřadovací místa jsou často vybírána vzhledem k termoregulaci a bezpečnosti.
Stromoví ptáci využívají šlachy svalu zadní končetiny, které jim umožňují bez neustálého stahování prstů se pevně zachytit na větvi, a tím se na ní udržet ve spánku. Mnoho ptáků pohybujících se po zemi, jako křepelky a bažanti, hřaduje na stromech. Někteří papoušci rodu Loriculus hřadují zavěšeni hlavou dolů. Někteří kolibříci každou noc upadají do stavu strnulosti, čímž se snižuje rychlost metabolismu. Tato fyziologická adaptace je známa u okolo sta dalších ptačích druhů včetně lelčíků, lelků a lasoletů. Jeden z druhů, lelek americký, dokonce vstupuje do stavu hibernace.
Rozmnožování
Sociální systémy

Většina (95 %) druhů ptáků jsou společensky monogamní; i když polygynie (2 %) a polyandrie (<1 %), polygamie, polygynandrie (kdy samice vytváří pár s několika samci a samec vytváří pár s několika samicemi) a smíšené systémy se vyskytují rovněž. Některé druhy mohou, podle okolností, využívat více než jeden systém. U monogamních druhů tvoří samci a samice páry na několik hnízdních období; v některých případech párové pouto může přetrvat mnoho let nebo dokonce celý život páru.
Výhodou monogamního vztahu pro ptáky je oboustranná péče rodičů o mláďata. U většiny zvířat je vzácností otcovská péče o mláďata, ale u ptáků je to docela běžné; ve skutečnosti je tato skutečnost rozsáhlejší u ptáků než u jakékoliv jiné třídy obratlovců. U ptáků je možno samčí péči o mláďata považovat za důležitou nebo rozhodující pro tělesnou zdatnost samic; u některých druhů nejsou samice schopny mláďata bez pomoci samců úspěšně odchovat. Polygamní systémy pramení ze schopnosti samic vychovávat mláďata bez pomoci samců. Dělba práce v sezení na vejcích, obraně hnízda, krmení mláďat a obraně území se vyskytuje někdy u monogamních párů.
Zatímco sociální monogamie je u ptáků zcela běžná, nevěra ve formě dodatečného párového páření se běžně vyskytuje u mnoha sociálně monogamních druhů. Tato nevěra může na sebe brát formu nuceného páření (nebo znásilňování) u kachen a dalších kachnovitých nebo je běžná u dominantních samců či samic s podřízenými partnery. Tím je pro samice a jejich potomky zajištěna možnost lepší genové výbavy nebo je tímto způsobem eliminována případná neplodnost partnerů. Samci některých druhů provozují mimopartnerské páření za účelem zvýšení pravděpodobnosti zabezpečení svého potomstva.
K rozmnožování patří také určité formy námluv, které jsou nejčastěji prováděny samcem. Nejčastěji jsou velmi jednoduché a obvykle zahrnují nějaký druh písně. Některá vystoupení mohou být docela propracovaná, jsou k tomu používány takové rozmanité metody jako bubnování křídly nebo ocasem, tancování, zásnubní lety nebo skupinové toky v závislosti na druzích. Samice si nejčastěji vybírají samce, i když např. u polyandrických lyskonohů si samečkové vybírají pestře zbarvené samičky. Námluvy krmením, cukrováním a čechráním peří se obvykle provádějí již mezi partnery, nejčastěji ptáky spárovanými a pářícími se.
Teritoria, hnízdění a inkubace
Mnoho ptáků brání v období páření své území před jinými druhy. Ochrana teritoria je důležitá z hlediska shánění potravy pro mláďata. Druhy ptáků, které nejsou schopny účinně bránit své teritorium, jako např. mořští ptáci nebo rorýsi, se sdružují do hnízdních kolonií; to je výhodné z hlediska ochrany proti . Hnízdící jedinci v koloniích pak ochraňují malé hnízdní plochy a soutěžení o tato místa, jak mezidruhové, tak uvnitř vlastního druhu, může pak nabývat velmi intenzivní podoby.

Všichni ptáci snášejí amniotická vejce s tvrdými skořápkami většinou z uhličitanu vápenatého. Barva vajec se řídí řadou faktorů: ta, která jsou snášena do děr nebo doupat, jsou většinou bílá nebo bledá, zatímco vejce v otevřených hnízdech, jako např. u dlouhokřídlých, bývají obvykle maskovaná. Existují samozřejmě výjimky z tohoto pravidla, např. většina zemních lelků má vejce bílá a maskování je zajišťováno peřím ptáků. Velikost a tvar vajec závisí na mnoha faktorech, včetně vývojových, fylogenetických a environmentálních.[80] Některé druhy, které jsou oběťmi hnízdního parazitismu prováděného např. některými kukačkami, mění barvy svých vajec, aby snížily pravděpodobnost snesení kukaččího vejce do svého hnízda, protože kukaččí samičky se snaží snést své vejce do hnízda hostitele s podobně zabarvenými vejci.
Vejce jsou obvykle uložena v hnízdě, které může být velmi propracované, jak je tomu u snovačů či vlhovců, nebo na druhou stranu velmi primitivní jako u některých albatrosů, kdy se hnízda skládají pouze z hromádky seškrábané ze zobáku, do níž kladou vejce. Některé druhy si nestaví žádné hnízdo, na útesech hnízdící alkouni úzkozobí kladou svá vejce na holou skálu a vejce tučňáka císařského jsou ukládána na nohy rodičů a svrchu jsou překryta kožním záhybem na břiše; tento způsob převládá u druhů hnízdících na zemi, kde nově vylíhlá mláďata jsou předčasně vyspělá. Většina ptačích druhů si staví propracovaná hnízda ve tvaru poháru, kupole, talíře, rámu nebo doupěte. Většina hnízd je stavěna tak, aby byla co nejméně nápadná, skrytá a uzavřená a tak chráněna před predátory, zatímco otevřená hnízda jsou stavěna ptáky v hnízdních koloniích nebo druhy schopnými účinné obrany svého hnízda. Hnízda jsou často stavěna na rostlinách, některé druhy si specificky vybírají rostliny jako řebříček obecný, který má látky, redukující hnízdní parazity jako jsou roztoči, čímž se zvyšuje šance mláďat na přežití. Hnízda jsou také často vystlána peřím, aby se zamezilo úniku tepla.
Sezení na vejcích, kterým je udržována optimální teplota pro vývoj mláďat, obvykle začíná s nakladením prvního vejce, proto se potomci líhnou ve stejném sledu, jak byla jednotlivá vejce snášena. U monogamních druhů se obvykle rodiče v sezení na vejcích střídají; u polygamních druhů plní veškeré rodičovské povinnosti jen jeden z rodičů. Teplo přechází u většiny druhů od rodičů k vejcím přes tzv. hnízdní nažinu, což je oblast holé kůže na břichu nebo hrudi na vejcích sedících ptáků. Sezení na vejcích může být energeticky náročný proces, např. dospělí albatrosi ztratí za den sezení až 83 gramů své tělesné váhy. Někteří ptáci, jako např. taboni, využívají jako teplo jiné zdroje než je sezení na vejcích. Patří mezi ně tlející rostliny nebo vulkanické zdroje tepla. Sezení na vejcích trvá od 10 dnů (u druhů jako datli, kukačky a pěvci) do 80 dnů (albatrosovití a kiviové).
Rodičovská péče a opeření

Po vylíhnutí mohou být mláďata podle druhu bezmocná až samostatná. Bezmocná mláďata jsou známa jako altriciální a jsou po narození zpravidla malá, holá a často slepá; mláďata po vylíhnutí pohyblivá a krytá prachovým peřím jsou prekociální, mláďata mohou být také poloprekociální nebo poloaltriciální. Altriciální mláďata potřebují pomoc při termoregulaci a vyžadují tak delší dobu k vysezení než mláďata prekociální.
Délka a povaha rodičovské péče je různá u jednotlivých řádů a druhů. Extrémem je např. rodičovská péče tabonů, která končí líhnutím; čerstvě vylíhlá kuřata se sama bez pomoci rodičů vyhrabou z hnízdní kupy listí a ihned jsou zcela soběstačná. Opačný extrém lze najít u některých mořských ptáků s prodlouženoi dobou péče o mláďata. Rodiči nejdéle se starajícími o své potomky jsou fregatky páskované, jejichž mláďata setrvávají ve hnízdě šest měsíců do opeření a poté jsou ještě dalších čtrnáct měsíců krmena.
U některých druhů pečují o mláďata oba rodiče, u jiných pak pouze jeden z nich. U některých druhů naopak při výchově mláďat pomáhají páru i další členové druhu. Tito pomocníci jsou obvykle blízcí příbuzní rodičů, většinou mláďata z předchozích snůšek. Společná výchova mláďat je obzvlášť běžná u krkavcovitých, ale byla pozorována i u dalších druhů jako jsou např. pokřovník zelený, luňák červený nebo flétňák australský.

Ve chvíli, kdy jsou mláďata plně opeřena, se rodiče některých druhů s mláďaty rozcházejí, zatímco u jiných druhů je vztah mezi starým ptákem a mládětem ještě dlouhou dobu zachován. Mláďata některých druhů opouštějí hnízdo hned po vylíhnutí, jak je tomu např. u alkounů, kdy mláďata následují v noci po vylíhnutí své volající rodiče na moře, čímž se vyhýbají nebezpečí, které na ně číhá v podobě pozemských predátorů. Některé jiné druhy, zvláště kachny, opouštějí své hnízdo v raném věku. U většiny druhů mláďata opouštějí hnízdo hned nebo krátce poté, co získají schopnost letu. Zároveň se také po opeření mění péče rodičů o mláďata; u albatrosů mláďata po odchodu z hnízda již nepřijímají od rodičů žádnou další pomoc, jiné druhy pokračují v určité doplňkové formě krmení mláďat. Mláďata také mohou spolu se svými rodiči absolvovat svou první migrační cestu.
Hnízdní parazitismus
Ačkoliv hnízdní parazitismus provozují i hmyz a ryby, nejvíce je rozšířen u ptáků. Principem je kladení vajec do hnízd jiných druhů. Tato vejce jsou často svými adoptivními rodiči přijata a vysezena i za cenu zničení jejich vlastních mláďat. Existují dvě skupiny ptáků provozujících hnízdní parasitismus: druhy, které nejsou schopny sami vysedět svá vejce a pro jejich vysezení musí nalézt vhodné hnízdo jiného ptačího druhu, a jiné, které jsou schopny mláďata za normálních podmínek vychovávat samy, ale příležitostně provozují i hnízdní parasitismus za účelem zvýšení svého reprodukčního výkonu. Okolo sta ptačích druhů, zahrnujících medozvěstkovité, vlhovcovité, astrildovité a kachnice, jsou běžnými hnízdními parazity, i když nejznámější jsou kukačkovití. Některá mláďata hnízdních parazitů mohou zničit celou snůšku hostitele buď vyhazováním vajec z hnízda, nebo zabíjením ostatních vylíhlých ptáčat; tím si zajišťují pro sebe všechnu potravu donášenou adoptivními rodiči.
Vztah člověka k ptákům
Do roku 1600 vyhubil člověk více než sto druhů ptáků a od roku 1600 přibližně 120 až 130 druhů. Kvůli člověku hrozí vyhynutí okolo 1200 ptačím druhům; ty jsou v současnosti chráněny.
Člověku poskytují bezprostřední užitek domácí ptáci a lovné druhy, tzv. pernatá zvěř.
Důležití jsou ptáci také díky potravě. Přispívají k šíření rostlin, protože se živí jejich semeny nebo jsou opylovači. Hmyzožravé druhy, sovy a draví ptáci s denní aktivitou jsou přirozenými bioregulátory polních a lesních škůdců. Druhy živící se bezobrtalými se živí i roztoči a hmyzem přenášejícím nemoci. Řada ptáků, a nejen dravých, se živí drobnými hlodavci, kteří mohou být hostiteli, rezervoáry a přenašeči patogenů přenosných na člověka.
Ptáci jsou také objektem chovatelství, lidé chovají okrasné ptactvo, dravé ptáky aj. a účastní se chovatelských akcí. Některé druhy, například pěvce nebo papoušky chovají lidé v domácnosti, pro potěšení. Zvláštním způsobem chovu ptáků, resp. lovu pomocí vycvičených dravců je sokolnictví.
Sčítání ptáků v ČR
Pozorování a sčítání ptáků na krmítku a v jeho okolí je program občanské vědy určený pro nejširší veřejnost. Program se nazývá Ptačí hodinka. Probíhá pod hlavičkou České společnosti ornitologické (ČSO) a přestože jsou výsledky součástí vědeckého výzkumu, může se zapojit každý zájemce o přírodu, protože předchozích zkušeností není třeba. Ptáky po dobu jedné hodiny sčítají jednotlivci nebo třeba rodiny, školní třídy, kroužky, družiny. Mohou sčítat jednou, dvakrát i třikrát, protože sčítání probíhá v pátek, v sobotu a v neděli, vždy v hodinu, kterou si sami zvolí – přirozeně, že je vhodnější sčítat v době, kdy jsou ptáci aktivní. Rozhodně to je zajímavější. Po dobu sčítání je třeba zůstat na vybraném místě (na zahradě, na balkoně, v parku, u lesa, doma u okna...); za chodu sčítat nelze.[81]
- První sčítání se konalo 4. až 6. ledna 2019 a zapojilo se do něj 14 333 dobrovolníků. Sečetlo se 278 232 ptáků.
- V roce 2020 (10. až 12. ledna) se do sčítání zapojilo 21 725 dobrovolníků, kteří sečetli celkem 436 301 ptáků.
- V roce 2021 (8. až 10. ledna) probíhalo sčítání od 8. do 10. ledna a sčítalo 27 123 dobrovolníků. Celkem se sečetlo 538 607 ptáků.
- V roce 2022 (7. až 9. ledna) se sčítání účastnilo 35 270 dobrovolníků, kteří odeslali 24 365 hodinových sčítání. Sečetlo se a bylo zaznamenáno 707 133 ptáků. Podrobné výsledky lze nalézt na webu https://ptacihodinka.birdlife.cz/vysledky Další ročník sčítání je naplánován na 6. až 8. ledna 2023.[82]
Podobně sčítají ptáky stovky tisíc lidí v USA, Velké Británii, Irsku, Nizozemsku, Rakousku, Německu a v dalších zemích.
Odkazy
Reference
- Kathleen O’Connor, Daniel J. Field and Corwin Sullivan (2021). Editorial: Early Avian Evolution. Frontiers in Earth Science. 9: 730214. doi: https://doi.org/10.3389/feart.2021.730214
- Hudec, K. et al. (2003). Soustava a české názvosloví ptáků světa, Muzeum Komenského Přerov, 462 s.
- Catherine Sheard, Montague H. C. Neate-Clegg, Nico Alioravainen, Samuel E. I. Jones, Claire Vincent, Hannah E. A. MacGregor, Tom P. Bregman, Santiago Claramunt & Joseph A. Tobias (2020). Ecological drivers of global gradients in avian dispersal inferred from wing morphology. Nature Communications, 11, Article number: 2463. doi: https://doi.org/10.1038/s41467-020-16313-6
- Caley, K. J. (2007). „Fossil birds“. Pp. 10–55 in del Hoyo, J., Elliott, A. and Christie, D. (eds.), Handbook of the Birds of the World, volume 12, Picathartes to Tits and Chickadees. Lynx Edicions, Barcelona.
- KUHL, Heiner; FRANKL-VILCHES, Carolina; BAKKER, Antje; MAYR, Gerald; NIKOLAUS, Gerhard; BOERNO, Stefan T; KLAGES, Sven. An Unbiased Molecular Approach Using 3′-UTRs Resolves the Avian Family-Level Tree of Life. S. 108–127. Molecular Biology and Evolution [online]. 2021-01-04 [cit. 2021-05-06]. Roč. 38, čís. 1, s. 108–127. DOI 10.1093/molbev/msaa191. (anglicky)
- Evan T. Saitta, Maximilian T. Stockdale, Nicholas R. Longrich, Vincent Bonhomme, Michael J. Benton, Innes C. Cuthill & Peter J. Makovicky (2020). An effect size statistical framework for investigating sexual dimorphism in non-avian dinosaurs and other extinct taxa. Biological Journal of the Linnean Society, blaa105. doi: https://doi.org/10.1093/biolinnean/blaa105
- https://phys.org/news/2021-05-avocet-zebra-finch-big-billion.html - From Avocet to Zebra Finch: big data study finds more than 50 billion birds in the world
- Oliver W. M. Rauhut & Christian Foth (2020). The Origin of Birds: Current Consensus, Controversy, and the Occurrence of Feathers. In: Foth C., Rauhut O. (eds). The Evolution of Feathers: 27-45. doi: https://doi.org/10.1007/978-3-030-27223-4_3
- https://www.nationalgeographic.com/magazine/2018/05/dinosaurs-survivors-birds-fossils/?beta=true
- https://hradec.rozhlas.cz/vladimir-socha-vsichni-dnesni-ptaci-jsou-prezivajici-dinosauri-ti-tak-vlastne-8151797
- https://dinosaurusblog.com/2019/09/05/ziji-dinosauri-vsude-kolem-nas/
- Cau A. (2018). The assembly of the avian body plan: a 160-million-year long process. Bollettino della Società Paleontologica Italiana 57: 1-25. doi:10.4435/BSPI.2018.01
- Dana J. Rashid; et al. (2018). Avian tail ontogeny, pygostyle formation, and interpretation of juvenile Mesozoic specimens. Scientific Reports 8. Article number: 9014 (2018). doi: https://doi.org/10.1038/s41598-018-27336-x
- Gegenbaur, C. (1863). „Vergleichend-anatomische Bemerkungen über das Fusskelet der Vögel“. Archiv far Anatomie und Physiologie wiss Medicin 1863:450–472
- https://veda.instory.cz/1229-dinosauri-stale-mezi-nami-nekteri-vedci-jiz-dlouhodobe-mluvi-o-tom-ze-vlastne-vubec-nevyhynuli.html
- Rozhovor o příbuznosti dinosaurů a ptáků s V. Sochou, portál Avokádo (1. 10. 2018)
- Yilun Yu, Chi Zhang, and Xing Xu (2021). Deep time diversity and the early radiations of birds. Proceedings of the National Academy of Sciences. 118(10): e2019865118. doi: https://doi.org/10.1073/pnas.2019865118
- Min Wang, Graeme T. Lloyd, Chi Zhang and Zhonghe Zhou (2021). The patterns and modes of the evolution of disparity in Mesozoic birds. Proceedings of the Royal Society B: Biological Sciences. 288(1944): 20203105. doi: https://doi.org/10.1098/rspb.2020.3105
- Gauthier, J.A. and de Queiroz,K. (2001). „Feathered dinosaurs, flying dinosaurs, crown dinosaurs, and the name 'Aves'“. Pp. 7–41 in Gauthier, J.A. and Gall, L.F. (eds.), New perspectives on the origin and early evolution of birds: Proceedings of the International Symposium in Honor of John H. Ostrom. New Haven, Connecticut, USA: Peabody Museum of Natural History, Yale University.
- Brown, J.W., Rest, J.S., Garcia-Moreno, J., Sorenson, M.D. and Mindell, D.P. (2008). „Strong mitochondrial DNA support for a Cretaceous origin of modern avian lineages“. BMC Biology 6:6.
- Chiappe, L.M. (1992) „Enantiornithine (Aves) tarsometatarsi and the avian affinities of the Late Cretaceous Avisauridae“. Journal of Vertebrate Paleontology. 12 (3): 344–350.
- Bakker, R. T. and Galton, P. M. (1974). „Dinosaur monophyly and a new class of vertebrates“. Nature. 248 (5444): 168–172.
- Maria E. McNamara; et al. (2018). Fossilized skin reveals coevolution with feathers and metabolism in feathered dinosaurs and early birds. Nature Communications 9. Article number: 2072. doi: 10.1038/s41467-018-04443-x
- Neil Brocklehurst & Daniel J. Field (2021). Macroevolutionary dynamics of dentition in Mesozoic birds reveal no long-term selection towards tooth loss. iScience. 102243. doi: https://doi.org/10.1016/j.isci.2021.102243
- Piotr Bajdek & Tomasz Sulej (2018). Hypothesis: Avian flight originated in arboreal archosaurs gliding on membranous wings. PeerJ Preprints 6:e27213v1. doi: https://doi.org/10.7287/peerj.preprints.27213v1
- Ashley M. Heers, Jeffery W. Rankin and John R. Hutchinson (2018). Building a Bird: Musculoskeletal Modeling and Simulation of Wing-Assisted Incline Running During Avian Ontogeny. Frontiers in Bioengineering and Biotechnology 6: 140. doi: https://doi.org/10.3389/fbioe.2018.00140
- Kuznetsov, A. N. and A. A. Panyutina (2022). Where was WAIR in avian flight evolution?. Biological Journal of the Linnean Society. doi: 10.1093/biolinnean/blac019
- Francisco José Serrano; et al. (2019). The effect of long-term atmospheric changes on the macroevolution of birds. Gondwana Research 65: 86-96. doi: https://doi.org/10.1016/j.gr.2018.09.002
- Joyce C. Havstad & N. Adam Smith (2019). Fossils with Feathers and Philosophy of Science. Systematic Biology. syz010. doi: https://doi.org/10.1093/sysbio/syz010
- Field, Daniel J.; Benito, Juan; Chen, Albert; Jagt, John W. M.; Ksepka, Daniel T. (2020). "Late Cretaceous neornithine from Europe illuminates the origins of crown birds". Nature. 579 (7799): 397–401. doi: 10.1038/s41586-020-2096-0
- Patrick M. O’Connor, Alan H. Turner, Joseph R. Groenke, Ryan N. Felice, Raymond R. Rogers, David W. Krause & Lydia J. Rahantarisoa (2020). Late Cretaceous bird from Madagascar reveals unique development of beaks. Nature. doi: https://doi.org/10.1038/s41586-020-2945-x
- Daniel T. Ksepka; et al. (2020). Tempo and Pattern of Avian Brain Size Evolution. Current Biology. doi: https://doi.org/10.1016/j.cub.2020.03.060
- https://www.sciencedaily.com/releases/2020/04/200423130506.htm
- Peter J. Makovicky & Sushma Reddy (2020). Evolution: Brainier Birds. Current Biology. 30(136): r778-r780. doi: https://doi.org/10.1016/j.cub.2020.05.025
- Ryan N. Felice, Akinobu Watanabe, Andrew R. Cuff, Michael Hanson, Bhart-Anjan S. Bhullar, Emily R. Rayfield, Lawrence M. Witmer, Mark A. Norell & Anjali Goswami (2020). Decelerated dinosaur skull evolution with the origin of birds. PLoS Biology, 18(8): e3000801. doi: https://doi.org/10.1371/journal.pbio.3000801
- SOCHA, Vladimír. Žijí ptáci všude kolem nás?. OSEL.cz [online]. 6. září 2019. Dostupné online. (česky)
- Xu, X., Zhou, Z. and Wang, X. (2000). "The smallest known non-avian theropod dinosaur." Nature 408 (6813): 705–708
- Wedel, M. J. (2007). „What pneumaticity tells us about ‚prosauropods‘, and vice versa“. Special Papers in Palaeontology 77: 207–222.
- Seymour, R.S., Bennett-Stamper, C.L., Johnston, S.D., Carrier, D.R. & Grigg, G.C. (2004). „Evidence for endothermic ancestors of crocodiles at the stem of archosaur evolution“. Physiological and Biochemical Zoology 77 (6): 1051–1067.
- Xu, X. and Guo, Y. (2009). „The origin and early evolution of feathers: insights from recent paleontological and neontological data“. Vertebrata PalAsiatica 47 (4): 311–329.
- Mayr, G., Pohl, B. and Peters, D.S. (2005). "A well-preserved Archaeopteryx specimen with theropod features". Science 310(5753): 1483–1486.
- Senter, P. (2006). "Scapular orientation in theropods and basal birds, and the origin of flapping flight". Acta Palaeontologica Polonica 51 (2): 305–313.
- Zhou Ya-Chun, Corwin Sullivan & Zhang Fu-Cheng (2019). Negligible effect of tooth reduction on body mass in Mesozoic birds. Vertebrata PalAsiatica 57 (1): 38-50. doi: 10.19615/j.cnki.1000-3118.180307
- Jeremy J. Klingler (2020). The evolution of the pectoral extrinsic appendicular and infrahyoid musculature in theropods and its functional and behavioral importance. Journal of Anatomy. doi: https://doi.org/10.1111/joa.13256
- Nebreda, S. M.; et al. (2020). Disparity and macroevolutionary transformation of the maniraptoran manus. In Pittman, M. and X. Xu (editors), Pennaraptoran theropod dinosaurs: past progress and new frontiers: 183-203. New York: Bulletin of the American Museum of Natural History.
- Fain, M.G. and Houde, P. (2004). „Parallel radiations in the primary clades of birds“. Evolution 58 (11): 2558–2573.
- Daniel Smith-Paredes; et al. (2018). Dinosaur ossification centres in embryonic birds uncover developmental evolution of the skull. Nature Ecology & Evolution (2018). doi: https://doi.org/10.1038/s41559-018-0713-1
- Luis Ossa‐Fuentes, Sergio Soto‐Acuña, Paula Bona, Michel Sallaberry & Alexander O. Vargas (2020). Developmental evolution of the distal ankle in the dinosaur–bird transition. Journal of Experimental Biology Part B: Molecular and Developmental Evolution. doi: https://doi.org/10.1002/jez.b.23022
- https://avokado-online.cz/dinosauri-a-ptaci-vladimir-socha-rozhovor-22/
- Cracraft, J. L., Barker, F. K., Braun, M., Harshman, J., Dyke, G. J., Feinstein, J., Stanley, S., Cibois, A., Schikler, P., Beresford, P., García-Moreno, J., Sorenson, M. D., Yuri, T. & Mindell, D. P. (2004). „Phylogenetic relationships among modern birds (Neornithes): toward an avian tree of life“. Pp. 468–489 in Cracraft, J. L. & Donoghue, M. (eds.), Assembling the Tree of Life. Oxford University Press, New York.
- Wetmore, A. (1960). „A classification for the birds of the world“. Smithsonian Miscellaneous Collection 139: 1–37.
- SIBLEY, Charles; ALQUIST, Jon E. Phylogeny and Classification of the Birds. New Haven, Connecticut, US: Yale University Press, 1990. Dostupné online. ISBN 9780300040852. (anglicky)
- Pheasants, partridges, francolins [online]. IOC World Bird List v12.1 [cit. 2022-02-08]. Dostupné online. (anglicky)
- České názvosloví ptáků světa [online]. Názvoslovná komise ČSO, 2021 [cit. 2022-02-12]. Dostupné online.
- BRAUN, Edward L.; KIMBALL, Rebecca T. Data Types and the Phylogeny of Neoaves. S. 1–22. Birds [online]. 2021-01-05 [cit. 2022-02-12]. Roč. 2, čís. 1, s. 1–22. Dostupné online. DOI 10.3390/birds2010001. (anglicky)
- HACKETT, Shannon J. et al. Evolutionary history a phylogenomic study of birds reveals their evolutionary history. Science. 2008, roč. 320, s. 1763-1768. Dostupné v archivu pořízeném dne 2013-08-19.
- BOYD, John. NEORNITHES: 46 Orders [online]. 2007 [cit. 2017-12-30]. Dostupné online. (anglicky)
- Nikita V. Zelenkov, Alexander V. Lavrov, Dmitry B. Startsev, Innessa A. Vislobokova & Alexey V. Lopatin (2019). A giant early Pleistocene bird from eastern Europe: unexpected component of terrestrial faunas at the time of early Homo arrival. Journal of Vertebrate Paleontology Article: e1605521. doi: https://doi.org/10.1080/02724634.2019.1605521
- SOCHA, Vladimír. Nejmenší ze všech teropodů. OSEL.cz [online]. 13. prosince 2018. Dostupné online. (česky)
- https://blogs.scientificamerican.com/laelaps/new-titleholder-for-the-largest-bird-of-all-time/
- https://www.sciencedaily.com/releases/2018/09/180926082706.htm
- http://www.osel.cz/10143-velkolepy-argentinsky-ptak.html
- Xing, Lida; O’Connor, Jingmai K.; Schmitz, Lars; Chiappe, Luis M.; McKellar, Ryan C.; Yi, Qiru; Li, Gang (11 March 2020). "Hummingbird-sized dinosaur from the Cretaceous period of Myanmar". Nature. 579 (7798): 245–249. doi:10.1038/s41586-020-2068-4
- http://www.osel.cz/10139-nejvetsi-zastupce-ptaci-rise.html
- http://www.osel.cz/10322-nejvetsi-ptaci-zabijak.html
- Ping Wu, Yung-Chih Lai, Randall Widelitz & Cheng-Ming Chuong (2018). Comprehensive molecular and cellular studies suggest avian scutate scales are secondarily derived from feathers, and more distant from reptilian scales. Scientific Reports 8, Article number: 16766 (2018). doi: https://doi.org/10.1038/s41598-018-35176-y
- Daňková, Kristýna (2017). Čichové schopnosti ptáků. bakalářská práce - Přírodovědecká fakulta Masarykovy univerzity, Brno
- Monica A. Daley & Aleksandra Birn-Jeffery (2018). Scaling of avian bipedal locomotion reveals independent effects of body mass and leg posture on gait. Journal of Experimental Biology, 221: jeb152538. doi: 10.1242/jeb.152538
- https://kof.zcu.cz/vusc/pg/termo09/termodynamics/prilohy/t7.htm
- Gabrielle Pavlovic, Michael A. Weston & Matthew R. E. Symonds (2018). Morphology and geography predict the use of heat conservation behaviours across birds. Functional Ecology. doi: https://doi.org/10.1111/1365-2435.13233
- Luigi Baciadonna, Francesca M. Cornero, Nathan J. Emery & Nicola S. Clayton (2020). Convergent evolution of complex cognition: Insights from the field of avian cognition into the study of self-awareness. Learning & Behavior. doi: https://doi.org/10.3758/s13420-020-00434-5
- Suzana Herculano-Houzel (2020). Birds do have a brain cortex--and think. Science. 369(6511): 1567-1568. doi: 10.1126/science.abe0536
- Akinobu Watanabe, Amy M. Balanoff, Paul M. Gignac, M. Eugenia L Gold & Mark A. Norell (2021). Novel neuroanatomical integration and scaling define avian brain shape evolution and development. bioRxiv. 2021.04.20.440700. doi: https://doi.org/10.1101/2021.04.20.440700
- Tzu-Ruei Yang & P. Martin Sander (2018). The origin of the bird's beak: new insights from dinosaur incubation periods. Biology Letters 14 5 20180090. doi: 10.1098/rsbl.2018.0090
- Geoffrey Ruaux, Sophie Lumineau and Emmanuel de Margerie (2020). The development of flight behaviours in birds. Proceedings of the Royal Society B: Biological Sciences, 287(1929): 20200668. doi: https://doi.org/10.1098/rspb.2020.0668
- Článek o rychlostním rekordu sokola stěhovavého na webu Osel.cz (česky)
- https://phys.org/news/2019-09-birds-migrate-night.html - Why do birds migrate at night?
- Tobias Riede , Scott L. Thomson, Ingo R. Titze & Franz Goller (2019). The evolution of the syrinx: An acoustic theory. PLoS Biology 17(2): e2006507. doi: https://doi.org/10.1371/journal.pbio.2006507
- Michael B. Habib (2019). New perspectives on the origins of the unique vocal tract of birds. PLoS Biology 17(3): e3000184. doi: https://doi.org/10.1371/journal.pbio.3000184
- Oksana V. Shatkovska, Maria Ghazali, Ivan S. Mytiai & Natalia Druz (2018). Size and shape correlation of birds' pelvis and egg: Impact of developmental mode, habitat, and phylogeny. Journal of Morphology. doi: https://doi.org/10.1002/jmor.20888
- Jak se zapojit – podrobněji. In: Česká společnost ornitologická [online]. [16. 2. 2022] [cit. 26. 3. 2022]. Dostupné z: https://ptacihodinka.birdlife.cz/jak-se-zapojit-podrobneji/#/observe
- Ptačí hodinka. In: Česká společnost ornitologická [online]. [16. 2. 2022] [cit. 26. 3. 2022]. Dostupné z: https://ptacihodinka.birdlife.cz/vysledky-tabulka/?period=ALL&place=ALL#/observe
Literatura
- Česky
- ACKERMAN, Jennifer. Ptačí způsoby: nový pohled na to, jak ptáci mluví, pracují, hrají si, vychovávají mláďata a přemýšlejí. Přeložila Alena KLVAŇOVÁ. Brno: Kazda, 2021. 376 s. ISBN 978-80-7670-045-1.
- ANDĚRA, Miloš a SOVÁK, Jan. Atlas fauny České republiky. Praha: Academia, 2018. 664 s. Atlas. ISBN 978-80-200-2756-6.
- BIRKHEAD, Tim. Ptačí smysl: jaké to je být ptákem. Přeložila Alena KLVAŇOVÁ. Brno: Kazda, 2020. 272 s. ISBN 978-80-88316-98-5.
- BURNIE, David et al. Ptáci: obrazová encyklopedie ptáků celého světa. V Praze: Knižní klub, 2008. 512 s. ISBN 978-80-242-2235-6.
- FUCHS, Roman et al. Atlas hnízdního rozšíření ptáků Prahy: 1985–1989 (aktualizace 2000–2002). Ilustroval Jan HOŠEK. Praha: Česká společnost ornitologická v nakl. Consult, 2002. 319 s. 1 ortofotomapa + 1 CD-ROM. ISBN 80-902132-5-1.
- HOFMANN, Helga. Ptáci z naší zahrady. Přeložil Vladimír Motyčka. Praha: Svojtka, 2016. 256 s. Průvodce přírodou. ISBN 978-80-256-1724-3.
- HUDEC, Karel. Ptáci v českém životě a kultuře. Praha: Academia, 2017. 452 s. ISBN 978-80-200-2628-6.
- CHVÁTAL, Marek, ed. Ptačí oblasti České republiky = Special protection areas of the Czech Republic. Praha: Aventinum, 2009. 88 s. ISBN 978-80-87051-53-5.
- KELLER, Verena et al. European breeding bird atlas. 2, Distribution, abundance and change. First edition. Barcelona: European bird census Council & Lynx editions, 2020. 967 s. ISBN 978-84-16728-38-1.
- KLVAŇOVÁ, Alena et al. Kam za ptáky v České republice. Praha: Grada, 2016. 263 s. ISBN 978-80-247-5778-0.
- KLVAŇOVÁ, Alena et al. Ptáci lidských sídel: příručka. Praha: Česká společnost ornitologická, 2020. 64 s. ISBN 978-80-87572-34-4.
- KOMÁREK, Stanislav. Ptáci v Čechách v letech 1360–1890, aneb, Tajemství rytíře von Sacher-Masocha: das Vorkommen der Vögel in Böhmen 1360–1890: (mit dt. Zusammenfassung des tschechischen Textes). Praha: Academia, 2007. 205 s. ISBN 978-80-200-1493-1.
- STORCH, David. Fylogeneze ptáků: vyřešeno? Vesmír. 2015, roč. 94, č. 10, s. 556–563.
- SVENSSON, Lars. Ptáci Evropy, severní Afriky a Blízkého východu. [3., přeprac. vyd., chybně uvedeno 2. vyd.] Přeložil Robert DOLEŽAL. Plzeň: Ševčík, 2016. 447 s. ISBN 978-80-7291-246-9.
- ŠŤASTNÝ, Karel et al. Atlas hnízdního rozšíření ptáků v České republice: 2014–2017. Praha: Aventinum, 2021. 511 s. ISBN 978-80-7442-130-3.
- ŠŤASTNÝ, Karel. Dravci, sokoli & sovy: v ilustracích Pavla Procházky. Praha: Aventinum, 2017. 336 s. Artia. ISBN 978-80-7442-086-3.
- ŠŤASTNÝ, Karel; BEJČEK, Vladimír a HUDEC, Karel. Atlas hnízdního rozšíření ptáků v České republice: 2001–2003. Praha: Aventinum, 2009. 463 s., volná příl. ISBN 978-80-86858-88-3.
- ŠŤASTNÝ, Karel; HUDEC, Karel; BEJČEK, Vladimír et al. Ptáci. (1), Pštrosi, tinamy, tučňáci, potáplice, potápky, trubkonosí, veslonozí, brodiví, plameňáci, vrubozubí, dravci, hrabaví. Praha: Albatros, 1998. 143 s. Svět zvířat, sv. 4. ISBN 80-00-00579-4.
Edice Fauna ČR
- ŠŤASTNÝ, Karel, ed.; HUDEC, Karel, ed., et al. Ptáci = Aves. Díl I. 3., přeprac. a dopl. vyd. Praha: Academia, 2016. Fauna ČR, sv. 31. ISBN 80-200-1834-4.
- HUDEC, Karel a ŠŤASTNÝ, Karel. Ptáci = Aves. Díl II/1. 2., přeprac. a dopl. vyd. Praha: Academia, 2005. 572 s. Fauna ČR, sv. 29/1. ISBN 80-200-1114-5.
- HUDEC, Karel a ŠŤASTNÝ, Karel. Ptáci = Aves. Díl II/2. 2. přeprac. a dopl. vyd. Praha: Academia, 2005. S. 581–1203. Fauna ČR, sv. 29/2. ISBN 80-200-1114-5.
- ŠŤASTNÝ, Karel, ed. a HUDEC, Karel, ed. Ptáci = Aves. Díl III/1. 2., přeprac. a dopl. vyd. Praha: Academia, 2011. 643 s. Fauna ČR, sv. 30/1. ISBN 978-80-200-1834-2.
- HUDEC, Karel a Karel ŠŤASTNÝ, ed. Ptáci = Aves. Díl III/2. 2., přeprac. a dopl. vyd. Praha: Academia, 2011. S. 649–1189. Fauna ČR, sv. 30/2. ISBN 80-200-1113-7.
- Anglicky
- V. B. BALIGA, I. SZABO and D. L. ALTSHULER (2019). Range of motion in the avian wing is strongly associated with flight behavior and body mass. Science Advances. 5(10): eaaw6670. doi: 10.1126/sciadv.aaw6670
- Christina HEROLD, Philipp SCHLÖME, Isabelle MAFOPPA-FOMAT, Julia MEHLHORN, Katrin AMUNTS & Markus AXER (2018). The hippocampus of birds in a view of evolutionary connectomics. Cortex. doi: https://doi.org/10.1016/j.cortex.2018.09.025
- Wei-Ling CHANG et al. (2019). The Making of a Flight Feather: Bio-architectural Principles and Adaptation. Cell. 179(6): P1409-1423.E17. doi: https://doi.org/10.1016/j.cell.2019.11.008
- Kenta KAWAHATA et al. (2019). Evolution of the avian digital pattern (PDF). Scientific Reports. 9, Article number: 8560. doi: https://doi.org/10.1038/s41598-019-44913-w
- Rebecca T. KIMBALL et al. (2019). A Phylogenomic Supertree of Birds. Diversity. 11(7): 109. doi: https://doi.org/10.3390/d11070109
- KUHL, Heiner; FRANKL-VILCHES, Carolina; BAKKER, Antje; MAYR, Gerald; NIKOLAUS, Gerhard; BOERNO, Stefan T; KLAGES, Sven. An Unbiased Molecular Approach Using 3′-UTRs Resolves the Avian Family-Level Tree of Life. S. 108–127. Molecular Biology and Evolution [online]. 2021-01-04 [cit. 2021-05-06]. Roč. 38, čís. 1, s. 108–127. DOI 10.1093/molbev/msaa191. (anglicky)
- Rebecca E. O’CONNOR et al. (2018). Patterns of microchromosome organization remain highly conserved throughout avian evolution. Chromosoma. doi: https://doi.org/10.1007/s00412-018-0685-6
- Joseph A. TOBIAS, Jente OTTENBURGHS & Alex L. PIGOT (2020). Avian Diversity: Speciation, Macroevolution, and Ecological Function. Annual Review of Ecology, Evolution, and Systematics. 51. doi: https://doi.org/10.1146/annurev-ecolsys-110218-025023
Externí odkazy
 Obrázky, zvuky či videa k tématu ptáci na Wikimedia Commons
Obrázky, zvuky či videa k tématu ptáci na Wikimedia Commons  Téma Pták ve Wikicitátech
Téma Pták ve Wikicitátech Slovníkové heslo pták ve Wikislovníku
Slovníkové heslo pták ve Wikislovníku- Česká společnost ornitologická
- Ptáci na BioLibu
- Databáze pozorování ptáků birds.cz
- Atlas ptáků a klíč pro identifikaci