Pavouci
Pavouci (Araneae) jsou členovci se čtyřmi páry nohou a chelicerami (klepítky), do kterých ústí vývod jedových žláz. Jsou největším řádem pavoukovců s vysokou druhovou rozmanitostí.[1] Pavouci se vyskytují na každém kontinentu kromě Antarktidy a téměř v každém prostředí kromě vzduchu a moře. K březnu 2021 bylo zaznamenáno 49 235 druhů a 128 čeledí pavouků;[2] nicméně ve vědecké komunitě panují nejasnosti v tom, jak tyto čeledi klasifikovat, což dokazuje počet více než 20 různých klasifikací, které byly navrženy od roku 1900.[3] V České republice bylo zaznamenáno 879 druhů, z toho 27 je považováno za v ČR již vyhynulé a dalších 483 se dle třetího vydání Červeného seznamu pavouků ČR nachází v různém stupni ohrožení.[4][pozn. 1]
 Diverzita pavouků | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | členovci (Arthropoda) |
| Podkmen | klepítkatci (Chelicerata) |
| Třída | pavoukovci (Arachnida) |
| Řád | pavouci (Araneae) Latreille, 1802 |
| podřády | |
| |
| Některá data mohou pocházet z datové položky. | |
Anatomicky se pavouci od ostatních členovců liší tím, že většinou mají tělo rozděleno na dvě tagmata (funkční celky), hlavohruď a zadeček, která jsou spojena malou válcovitou stopkou (pedicelem). Na rozdíl od hmyzu nemají tykadla. Až na nejprimitivnější skupinu, sklípkoše (Mesothelae), mají pavouci ze všech členovců nejvíce centralizovanou nervovou soustavu, protože všechny jejich gangliony (nervové uzliny) se nacházejí v jednom místě v hlavohrudi. Na rozdíl od většiny členovců nemají ve svých končetinách téměř žádné extenzory (natahovače) a namísto toho je natahují hydraulickým tlakem.
Zadečkové končetiny pavouků se vyvinuly ve snovací bradavky produkující pavučinová vlákna až z šesti druhů snovacích žláz, jež se nacházejí v zadečku. Pavučiny se mohou lišit velikostí, tvarem a množstvím použitého lepkavého vlákna. Zdá se, že kruhové pavučiny jsou jedny ze starších druhů pavučin. Pavouci vytvářející pavučinové vaky jsou rozšířenější a rozmanitější než křižákovití (kteří vytvářejí právě kruhové pavučiny). Pavoukům podobní pavoukovci s orgány produkujícími pavučinová vlákna se objevili v devonu někdy před 386 miliony lety, ale těmto živočichům chyběly snovací bradavky. Praví pavouci byli nalezeni v horninách z karbonu, přičemž pocházejí z období před 318 až 299 miliony lety a jsou velmi podobní nejprimitivnějšímu žijícímu podřádu, sklípkošům. Nejvýznamnější skupiny moderních pavouků, sklípkani (Mygalomorphae) a dvouplicní (Araneomorphae), se objevily v triasu před 200 miliony lety.
V roce 2008 byla popsána potravní specializace druhu Bagheera kiplingi,[5] ale všechny ostatní druhy jsou predátoři. Živí se převážně hmyzem a ostatními pavouky, i když několik velkých druhů loví i drobné savce, ptáky a plazy. K lovení kořisti pavouci využívají různé strategie: chytají ji do lepkavých pavučin; vrhají na ni lepkavá vlákna; napodobují ji, aby je neodhalila; nebo ji uženou. Většina z nich kořist odhaluje pomocí vibrací, ale aktivní lovci mají dobrý zrak. Lovci z rodu Portia vykazují znaky inteligentního chování při výběru taktiky lovu a ve schopnosti vytvářet nové. Střeva pavouků jsou příliš úzká na to, aby mohla přijímat pevnou potravu, a proto ji rozmělňují trávicími enzymy a drtí ji makadly (pedipalpy), protože nemají pravé čelisti.
Samečci na sebe upozorňují různými namlouvacími rituály, aby je samičky nesežraly. Samečci většiny druhů přežívají několik páření a umírají z jiných příčin, než že by je sežrala samice. Samičky z pavučinových vláken vytvářejí kokony na vajíčka, z nichž každý může obsahovat až stovky vajíček. Samičky mnoha druhů se starají o svá mláďata, například tak, že je nosí s sebou či se s nimi dělí o potravu. Malá část druhů je sociální a vytváří společné pavučiny, které mohou sloužit několika málo až 50 000 jedinců. Sociální chování sahá od opatrné tolerance až ke společnému lovení a sdílení potravy. Ačkoliv se většina pavouků dožívá maximálně dvou let, sklípkani se v zajetí mohou dožít až 25 let.
I když jed některých druhů je pro člověka nebezpečný, vědci zkoumají potenciální využití pavoučího jedu v lékařství a v zemědělství (jako pesticid neznečišťující prostředí). Pavoučí vlákno představuje lepší kombinaci lehkosti, síly a pružnosti než umělé materiály, a vědci zkoušeli vložit geny produkující tato vlákna do DNA různých savců a rostlin, aby zjistili, zda tyto organismy mohou vlákna vytvářet. Kvůli svému rozmanitému chování se pavouci stali v umění a mytologii symbolem trpělivosti, krutosti a kreativity.
Popis
Stavba těla
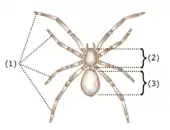
(1) čtyři páry nohou
(2) hlavohruď
(3) zadeček
Pavouci jsou klepítkatci a tudíž členovci.[6] Jako všichni členovci mají článkovité tělo s párovými končetinami, pokryté kutikulou z chitinu a bílkovin; a hlavu složenou z několika částí, které splývají v průběhu embryonálního vývoje.[7] Jakožto klepítkatci mají tělo složené ze dvou tagmat neboli souborů článků, které mají podobnou funkci: přední část se nazývá hlavohruď neboli cephalothorax a skládá se z článků, které u hmyzu vytvářejí dvě různá tagmata, hlavu a hruď; zadní část se nazývá zadeček neboli abdomen.[6] U pavouků jsou hlavohruď a zadeček spojeny malou válcovitou stopkou, pedicelem.[8] Průběh splývání částí, z kterých vzniká hlava klepítkatců, je mezi členovci jedinečný: část, která obvykle vytváří nejpřednější část hlavy, u nich zaniká v brzkém stádiu vývoje, takže ve výsledku nemají tykadla, která jsou pro většinu členovců typická. Jedinými končetinami nacházejícími se před ústy je pár chelicer (klepítek); klepítkatci nemají žádné „čelisti“.[7][9] První končetiny za ústy se nazývají makadla a u různých skupin klepítkatců slouží různým účelům.[6]
Spolu se štíry a dalšími skupinami patří pavouci mezi pavoukovce.[9] Chelicery štírů mají tři části a slouží k přijímání potravy.[10] Chelicery pavouků mají dvě části a jsou zakončeny drápky, které jsou většinou jedovaté a když je pavouci zrovna nepoužívají, nosí je ohnuté. Horní části mívají husté ochlupení tvořící „síto“, které nepropouští pevné kusy potravy, protože pavouci mohou přijímat pouze tekutou potravu.[8] Makadla štírů většinou tvoří velká klepeta, kterými štíři loví potravu,[10] zatímco pavouci mají relativně malá makadla, jejichž konce ovšem také slouží ke krmení; navíc samci mají konce zvětšené a využívají je k přenosu spermatu.[8]
Stopka, kterou jsou spojeny hlavohruď a zadeček, umožňuje zadečku nezávislý pohyb při vytváření pavučinových vláken. Horní strana hlavohrudi je pokryta konvexním karapaxem a spodní strana je kryta dvěma plochými destičkami. Zadeček je měkký a má vejcovitý tvar. Pouze u sklípkošů je jeho horní strana členitá, jinak je tvořen jedním celkem.[8]
Oběhová a dýchací soustava
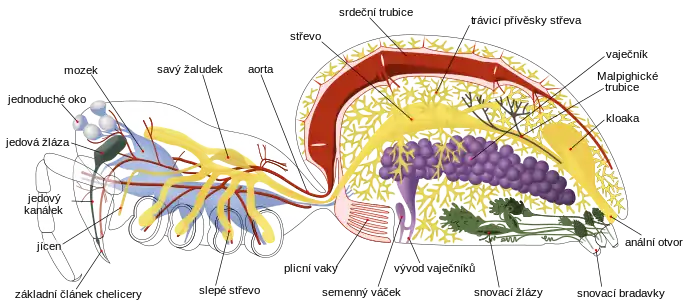
Stejně jako ostatní členovci mají coelom (pravou tělní dutinu), který se u nich vyskytuje pouze v malých oblastech v okolí rozmnožovací a vylučovací soustavy. Jeho místo zabírá převážně dutina probíhající téměř celým tělem (tzv. mixocoel), kterou proudí krvomíza - hemolymfa . Srdce je tvořeno trubicí v horní části těla a několika polopropustnými chlopněmi, které umožňují vstup hemolymfy z mixocoelu do srdce.[11] U pavouků ale zaujímá pouze horní část zadečku a hemolymfa je vylučována do mixocoelu jednou tepnou, která ústí do zadní části zadečku, a dalšími rozvětvujícími se tepnami, které procházejí stopkou a ústí do několika částí hlavohrudi. Proto mají pavouci otevřenou oběhovou soustavu.[8] Hemolymfa mnoha pavouků s plicními vaky obsahuje respirační pigment (dýchací barvivo) hemocyanin, který umožňuje efektivnější přenos kyslíku.[9]
U pavouků se vyvinuly různé druhy dýchacích soustav založené na plicních vacích, vzdušnicích (trachejích) či na kombinaci těchto dvou orgánů. Sklípkani a sklípkoši mají dva páry plicních vaků naplněných hemolymfou s otvory na spodní straně zadečku, kterými se k nim dostává vzduch a dochází k přenosu kyslíku. To platí i pro některé vývojově starší druhy z řádu dvouplicní, například z čeledi Hypochilidae, ale u ostatních zástupců tohoto řádu je zachován pouze přední pár plicních vaků a zadní pár je částečně či úplně přetvořen ve vzdušnice, skrze které je kyslík přenášen do hemolymfy či přímo do tkání a orgánů.[8] Dýchací soustava založená na vzdušnicích se vyvinula pravděpodobně u předků menšího vzrůstu za účelem odolávání desikaci (vysoušení).[9] Vzdušnice byly původně propojeny s okolím skrze pár otvorů zvaných spiracula, ale u většiny pavouků tento pár splynul v jedno spiraculum umístěné uprostřed a posunul se dozadu ke snovacím bradavkám.[8] Pavouci se vzdušnicemi mají většinou vyšší metabolický výdej a lépe v těle udržují vodu.[12]
Trávicí a vylučovací soustava

Na rozdíl od ostatních klepítkatců jsou poslední články pavoučích chelicer přetvořeny v drápky, jež velká většina pavouků používá ke vstřikování jedu do své kořisti. Jed je vytvářen jedovými žlázami v kořenech chelicer.[8] Pavouci z čeledi pakřižákovití (Uloboridae) a zástupci rodu Holarchaea o jedové žlázy přišli a svou kořist usmrcují pevným zabalením a agresivními trávicími enzymy.[13] Stejně jako většina pavoukovců[9] mají pavouci úzká střeva, která jsou schopna přijímat pouze potravu v tekutém stavu. K filtrování pevné stravy slouží dvě sady filtrů.[8] Pavouci používají dva různé druhy externího trávení. Někteří z nich napouštějí svou kořist trávicími enzymy a poté vysávají její rozpuštěné tkáně, až z ní zůstane pouze prázdná slupka. Jiné druhy drtí svou kořist na kaši pomocí chelicer a makadel, zatímco ji napouštějí enzymy; u těchto druhů chelicery a makadla vytvářejí dutinu, která drží zpracovávanou potravu.[8]
Žaludek v hlavohrudi slouží jako čerpadlo, které posouvá potravu hlouběji do trávicí soustavy. Pavouci mají několik slepých střev, ve kterých se z potravy vstřebávají živiny; většina z nich se nachází v zadečku, v němž největší prostor zaujímá právě trávicí soustava, ale několik se jich nachází i v hlavohrudi.[8]
Většina pavouků přetváří dusíkaté odpadní látky v kyselinu močovou, kterou pak vylučují v suchém stavu. Malpighické trubice tyto odpadní látky extrahují z hemolymfy v mixocoelu a vypouští je do kloaky, odkud jsou vyloučeny řitním otvorem.[8] Vytváření kyseliny močové a její vylučování přes Malpighické trubice slouží k udržování vody v těle a tento proces se vyvinul nezávisle u několika linií členovců, kteří jsou schopni žít daleko od vody;[14] například trubice hmyzu a pavoukovců se vyvinuly z naprosto odlišných částí embrya.[9] Někteří primitivní pavouci, jako například druhy podřádů sklípkoši a sklípkani, si ale zachovali nefridie („malé ledviny“),[8] které spotřebují velké množství vody k vylučování dusíkatých odpadních látek ve formě amoniaku.[14]
Centrální nervový systém
V základní podobě se centrální nervová soustava členovců skládá z páru nervových trubic podbíhajících pod střevem a párových ganglií, která slouží jako místní řídící centra ve všech článcích; a mozku, který vznikl splynutím ganglií hlavových článků nacházejících se před a za ústy, takže tento shluk obklopuje jícen.[15] Kromě primitivních sklípkošů mají pavouci nerovovou soustavu více centralizovanou: všechna ganglia za jícnem splynula do sebe, takže v hlavohrudi se nachází velké množství nervových tkání a v zadečku nejsou žádná ganglia;[8][9][15] u sklípkošů zůstávají ganglia zadečku a zadní části hlavohrudi oddělena.[12]
Oči

Většina pavouků má vepředu nahoře na hlavohrudi čtyři páry očí, jejichž rozmístění se liší čeleď od čeledi.[8] Čelní pár je tvořen z takového druhu jednoduchých očí (očí s pouze jednou čočkou), které jsou u většiny členovců schopny rozeznávat pouze směr, z něhož na ně dopadává světlo. U pavouků jsou tyto oči ale schopny vytvářet obrazy.[16][17] U ostatních očí se předpokládá, že se vyvinuly ze složených očí klepítkatců, ale už se neskládají z jednotlivých oddělených oček, která jsou pro složené oči typická. Na rozdíl od hlavních očí tyto vedlejší oči u mnoha pavouků vnímají světlo odražené od tapeta lucida a slíďákovité lze ve tmě rozeznat podle toho, že jim oči „svítí“. Na druhou stranu například vedlejší oči skákavkovitých tapeta nemají.[8] Zraková ostrost některých skákavkovitých přesahuje až desetinásobně zrakovou ostrost šídel, která mají mezi hmyzem zdaleka nejlepší zrak; lidé mají pouze asi pětkrát ostřejší zrak než skákavkovití. Dosahují toho pomocí čoček poskládaných na způsob teleobjektivu, čtyřvrstvé sítnice a schopnosti otáčet oči a sjednotit obrazy z různých fází snímání okolí. Nevýhodou procesu sjednocování obrazů je jeho pomalost.[18] Někteří pavouci mají menší počet očí, nejčastěji šest (např. čeleď šestiočkovití, ale i jen čtyři nebo dvě. Jeskynní druhy mívají oči zakrnělé.
Ostatní smysly
Stejně jako u ostatních členovců by pavoučí kutikula normálně bránila průniku informací z okolního světa. Pavouci a jiní členovci ji ale mají prostoupenou důmyslnou sadou senzorů. Různá dotyková čidla, například štětiny, reagují na síly různých velikostí od silného dotyku po slabé závany větru. Chemická čidla poskytují ekvivalent chuti a čichu, často skrze štětiny.[16] Pavouci mají také v kloubech končetin speciální orgány, které reagují na vibrace. U pavouků, kteří vytvářejí pavučiny, jsou tyto mechanické a chemické senzory důležitější než oči, které jsou naopak nejdůležitějším smyslovým orgánem pro aktivně lovící pavouky.[8]
Pavouci, stejně jako většina členovců, nemají čidla pro vnímání stability a zrychlení, a to, kde je nahoře a kde dole, rozpoznávají očima. Proprioreceptory členovců, které vnímají napětí svalů a ohyb kloubů, jsou důkladně prozkoumány. Na druhou stranu se toho příliš neví o tom, jaké další vnitřní smyslové orgány pavouci a členovci obecně mají.[16]
Pohyb
Každá z osmi nohou sestává ze sedmi částí. Část nejblíže k tělu se nazývá kyčel (coxa); další částí je příkyčlí (trochanter), které funguje jako pant pro další část, stehno (femur); následuje koleno (patella), které slouží jako pant pro holeň (tibia); další je nárt (metatarsus), který spojuje holeň s chodidlem (tarsus); chodidlo je v závislosti na čeledi zakončeno dvěma nebo třemi drápky. Všichni členovci ohýbají své končetiny pomocí svalů připevněných na vnitřní stranu vnější kostry; k jejich natahování využívají pavouci a několik dalších skupin hydraulický tlak, což je způsob zděděný po předcích členovců.[19] Jediné extenzory (natahovače) v tělech pavouků jsou umístěny ve třech kyčelních kloubech (nacházejících se mezi kyčlí a příkyčlí).[20] Následkem toho pavouk s probodnutou hlavohrudí nemůže natahovat nohy a nohy mrtvých pavouků se kroutí.[8] Pavouci jsou schopni za účelem natažení nohou vyvinout tlak až osmkrát vyšší než v klidovém stavu[21] a skákavkovití dokáží vyskočit do výše až padesátinásobku délky svého těla tím, že zvýší krevní tlak ve třetím či čtvrtém páru nohou.[8] Ačkoliv větší pavouci také používají k natahování nohou hydrauliku, na rozdíl od skákavkovitých ke skokům využívají flexory (ohybače).[20]
Většina aktivně lovících pavouků má mezi drápky na konci nohou husté chomáče chloupků. Tyto chomáče neboli skopuly obsahují chloupky, jejichž konce jsou rozvětveny až na tisíce konečků a umožňují pavoukům lézt po vertikálně položeném skle a vzhůru nohama po stropě. Tato přilnavost je zřejmě umožněna díky kontaktu s velmi tenkými vrstvami vody na površích.[8] Pavouci při chůzi či při běhu nechávají na povrchu nejméně čtyři nohy, stejně jako ostatní pavoukovci.[22]
Vytváření pavučinových vláken

Zadeček nemá žádné končetiny kromě těch, z kterých se vyvinuly krátké pohyblivé snovací bradavky produkující pavučinová vlákna. Bývá jich jeden až čtyři páry, přičemž nejčastěji jsou to tři páry. Každá snovací bradavka má několik vývodů (spigotů), z nichž každý je propojen s jednou žlázou. Existuje alespoň šest druhů snovacích žláz, z nichž každá vytváří jiný druh vláken.[8] Sklípkani mají snovací žlázy také v nohou.[23]
Pavučinové vlákno je složeno převážně z bílkoviny podobné té, která se nachází v hmyzích hedvábných vláknech. Původně se jedná o kapalinu, která tvrdne ne kvůli kontaktu se vzduchem, ale protože proces vylučování mění vnitřní strukturu bílkoviny.[24] Má podobnou pevnost v tahu jako nylon a přírodní materiály jako chitin, kolagen a celulóza, ale je mnohem elastičtější, což znamená, že ho lze více natahovat, aniž by se přetrhlo či ztratilo tvar.[8]
Někteří pavouci mají sítko (cribellum), což je speciální orgán s až 40 000 spigoty, z nichž každý vytváří jedno malé vlákno. Vlákna jsou rozčesávána hřebínkem (calamistrum) a výsledné vlákno je velmi účinné při zachycování ochlupených hmyzích končetin. Původní pavouci měli cribella, která produkovala první vlákna schopná chytat hmyz, a až poté se vyvinuly žlázy produkující lepkavé kapičky. Většina novodobých skupin pavouků již cribellum ztratila.[8]
I druhy, které nevytvářejí pavučiny, využívají pavučinová vlákna několika různými způsoby: jako obal pro samčí pohlavní buňky a pro oplodněná vajíčka; jako „záchranný provaz“; pro budování hnízd; a mláďata některých druhů jako „padák“.[8]
Rozmnožování a životní cyklus

Pavouci se rozmnožují pohlavně a oplodnění probíhá vnitřně, ale nepřímo, což znamená, že samec nevnáší sperma do samičího těla svým pohlavním ústrojím. Na rozdíl od mnoha suchozemských členovců[25] samci pavouků nevytvářejí spermatofory, ale vytvářejí malé pavučiny, do kterých ejakulují, a poté sperma přenesou do objektů podobných injekční stříkačce na konci makadel. Když samec ve svém okolí zaznamená přítomnost samice, zjistí, zda se jedná o samici stejného druhu a zda je připravena na páření; například u druhů, které vytvářejí pavučiny či „záchranné provazy“, rozpoznává samec druh a pohlaví pomocí „čichu“.[8]


Malí samci často využívají složité namlouvací rituály k tomu, aby je velké samice nesežraly před oplodněním, ale u některých druhů jsou samci tak malí, že by se samicím stejně nevyplatilo je jíst. U pavouků vytvářejících pavučiny je hlavní součástí rituálu důmyslný systém vibrací, zatímco u aktivně lovících pavouků hrají velkou roli doteky samičího těla, které dokážou samici „zhypnotizovat“. Pro skákavkovité, kteří mají dobrý zrak, jsou důležité různé posunky a tance. Při úspěšných námluvách samec vnese spermie ze svých makadel do samičího pohlavního otvoru skrze pohlavní destičku (epigyne) na spodní straně jejího zadečku. Samičí rozmnožovací soustava může sestávat pouze z jednoduchých trubic či může obsahovat i schránky, ve kterých samice skladuje spermie do doby vhodnější pro oplodnění.[8]
Samcům z rodu Tidarren před dospěním upadává jedno makadlo. U těchto pavouků představují makadla 20 % hmotnosti samčího těla, a když se jednoho z nich zbaví, zvýší se jim pohyblivost. U jemenského druhu Tidarren argo zbývající makadlo utrhává samice. To poté asi čtyři hodiny zůstane zachycené v její pohlavní destičce a zřejmě dále funguje. Mezitím samice samce sežere.[26] Ve více než 60 % případech samice australské snovačky Hasseltovy zabije a sežere samce ve chvíli, kdy do jejího pohlavního otvoru vloží své druhé makadlo; samci jí v tom dokonce pomáhají tím, že se snaží nabodnout na její drápky. Většina samců tohoto druhu totiž nikdy nedostane možnost se pářit a ti, kterým se to podaří, se snaží navýšit počet mláďat tím, že zajistí, aby byla samice dobře krmena.[27] Ale samci většiny druhů přežijí několik páření a umírají z jiných příčin než usmrcením samicí. Někteří dokonce nějakou dobu žijí v pavučinách svých partnerek.[28]
Samice kladou až 3000 vajíček do jednoho či více kokonů z pavučinových vláken,[8] které si udržují víceméně konstantní vlhkost vzduchu.[28] Samice některých druhů poté zemřou, ale samice jiných druhů kokony chrání tak, že je uchytí ke své pavučině, schovají v hnízdě, nosí v chelicerách nebo přichytí ke snovacím bradavkám.[8]
Mláďata tráví celé své larvální stadium uvnitř vajíčka a když se vylíhnou, jsou podobná dospělým jedincům, jen jsou menší. Někteří pavouci o svá mláďata pečují, například samice slíďákovitých nosí kokon s vajíčky připevněný na zadečku[8] a samice některých druhů reagují na „škemrání“ svých mláďat tak, že jim dávají svou kořist či dokonce vyvrhují potravu.[28]
Stejně jako ostatní členovci se při růstu musí svlékat, protože jejich kutikula se nedokáže roztáhnout.[29] Samci některých druhů se páří s čerstvě vysvlečenými samicemi, které jsou příliš slabé na to, aby pro ně byly nebezpečné.[28] Většina pavouků žije pouze jeden až dva roky, ale někteří sklípkani mohou v zajetí přežít více než 20 let.[8][30]

Velikost
Pavouci dorůstají různorodých velikostí. Délka těla těch nejmenších, mezi které patří například Patu digua z Kolumbie, nepřesahuje 0,37 milimetrů. Tělo největších sklípkanů dorůstá až do délky 90 milimetrů a rozpětí nohou mají až 250 milimetrů.[31]
Chování
Lov kořisti

Nejznámější metodou lovu kořisti je její chycení do lepkavé pavučiny. Různá umístění pavučiny umožňují různým druhům pavouků v jedné oblasti chytat odlišné druhy hmyzu, například horizontálně umístěné sítě zachycují hmyz, který vzlétá z rostlin pod nimi, zatímco vertikálně umístěné sítě zachycují hmyz v horizontálním letu. Pavouci, kteří budují pavučiny, mají špatný zrak, ale jsou velmi citliví na vibrace.[8]
Samičky vodního pavouka vodoucha stříbřitého (Argyroneta aquatica) stavějí pod vodou „zvon“ z pavučin, který plní vzduchem a využívají ho k trávení potravy, svlékání, páření a kladení vajíček. Pavouci tohoto druhu tráví ve zvonech téměř celý svůj život a vylézají jenom za účelem chycení kořisti, která se zvonu nebo k němu přilehlých vláken dotkla.[32] Několik druhů využívá hladiny jezer a rybníků podobně jako pavučiny tak, že na nich rozpoznává vibrace, které způsobuje hmyz spadlý do vody a snažící se dostat ven.[8]
Pavouci, kteří loví vrháním vláken, vytvářejí pouze malé pavučiny, kterými následně hýbají. Zástupci rodu pakřižák (Hyptiotes) a čeledi křižáčkovití (Theridiosomatidae) své pavučiny natahují, a když se jich kořist dotkne, tak je uvolní. Jedinci z čeledi vrhačovití (Deinopidae) vytvářejí ještě menší pavučiny, které drží mezi prvními dvěma páry nohou. Když zahlédnou potenciální kořist, pavučinu roztáhnou, vyrazí s ní vpřed a kořist chytí. Plocha pavučiny se tímto roztažením může zvětšit až desetkrát. Několik druhů vrhačovitých využívá dvě různé techniky lovu: útočí buď nahoru na létající hmyz (který detekuje pomocí vibrací) nebo dolu na hmyz pohybující se po zemi (který zpozoruje). Převážnou část potravy většiny vrhačovitých tvoří pozemní hmyz, ale byla pozorována i populace druhu Deinopis subrufa, která se živila hlavně létajícími tiplicemi (Tipulidae).[33]
Dospělé samice bolasových pavouků vytvářejí „pavučiny“ sestávající z jediného vlákna, které střeží. Loví pomocí „lasa“, což je pavučinové vlákno zakončené velkou lepkavou koulí. Vylučují látky připomínající můří feromony a poté na můry vrhají svá lasa. Ačkoli asi v polovině případů minou, za noc jsou schopny ulovit hmyz zhruba o stejné hmotnosti jako pavouci stejné velikosti, kteří vytvářejí pavučiny. Když se jim zhruba do půl hodiny nepodaří ulovit žádnou potravu, sežerou laso, chvíli si odpočinou a poté vytvoří nové.[34][35] Mláďata a dospělí samci jsou mnohem menší a lasa nevytvářejí. Místo toho vylučují feromony, kterými lákají koutulovité (Psychodidae), a tyto molům podobné mušky chytají předními páry nohou.[36]
.jpg.webp)
Sklípkošovití (Liphistiidae), sklípákovití (Ctenizidae), a mnoho sklípkanů přepadává kořist ze svého doupěte, které je často kryto poklopem a/nebo obklopeno systémem pavučinových vláken, které pavouky upozorňují na přítomnost potenciální kořisti.[12] Několik druhů, které se živí včelami, dokáže nastavit odrazovost ultrafialového záření od svého těla tak, aby výsledek byl podobný rostlinám, mezi kterými se ukrývají. (Včely vidí ultrafialové záření.)[37] Slíďákovití, skákavkovití, běžníkovití a lovčíci (Dolomedes) svou kořist honí a spoléhají se převážně na svůj zrak.[8]

Někteří pavouci z rodu Portia loví ostatní pavouky způsobem, ve kterém lze spatřit náznaky inteligentního chování.[18] Laboratorní výzkumy ukázaly, že instinktivní způsob lovu je pro tyto pavouky jen počátečním bodem a svou lovící taktiku pilují metodou pokus a omyl.[38]

Pavouci napodobující mravence se od ostatních pavouků liší v několika věcech: mají užší zadeček a umělý „pas“ v hlavohrudi, aby napodobili tři oddělená tagmata mravenčích těl; prvním párem nohou napodobují tykadla, protože je nemají a protože mravenci mají jen tři páry nohou; okolo jednoho páru očí mají velké barevné skvrny, aby zakryli fakt, že (většinou) mají osm jednoduchých očí a mravenci mají dvě složené oči; a pokrývají své tělo chloupky odrážejícími světlo, aby tak napodobili lesklé tělo mravenců. U některých druhů samec a samička napodobují odlišné druhy mravencovitých, protože samičky jsou většinou mnohem větší než samci. Imitátoři také uzpůsobují své chování tak, aby připomínalo chování jejich mravenčích protějšků: například napodobují jejich způsob pohybu a vyhýbají se skákání. Napodobování mravenců slouží u mnoha pavouků a dalších členovců jako ochrana před predátory, kteří k lovu využívají zrak, jako jsou například ptáci, ještěrky a pavouci. Někteří z nich se ale živí mravenci či živočichy, které mravenci opečovávají, což jsou například mšice. Pavouk rodu Amyciaea v klidu nevypadá jako mravenec rodu Oecophylla, ale loví tak, že napodobuje chování umírajícího pavouka a láká tak mravenčí dělnice.[39]
Jiné způsoby opatřování potravy
Pavouci jsou většinou draví živočichové, ale Bagheera kiplingi z čeledi skákavkovití přijímá více než 90 % své potravy ve formě rostlinného materiálu, který produkují akácie v rámci vzájemně prospěšného vztahu s mravenci.[40]
Mláďata některých druhů z čeledí šplhavkovití (Anyphaenidae), hlavounovití (Corinnidae), zápředníkovití (Clubionidae), běžníkovití (Thomisidae) a skákavkovití (Salticidae) se živí rostlinným nektarem. I v laboratorních pokusech dávají tito pavouci přednost sladkým roztokům před vodou, z čehož lze usuzovat, že vyhledávají živiny. Nektar kromě cukru obsahuje aminokyseliny, lipidy, vitamíny a minerály a z různých studií vyplývá, že některé druhy pavouků žijí déle, když ho mají k dispozici. Živení se nektarem odstraňuje riziko spojené s lovením potravy a potřebu vytváření jedu a trávicích enzymů.[41]
Některé druhy se živí mrtvými členovci (mrchožroutství), pavoučími vlákny nebo svou svléknutou kutikulou. Někdy žerou i pyl zachycený v pavučinách a z pozorování vyplývá, že mláďata mají větší šanci přežití, pokud mají možnost ho žrát. V zajetí se pavouci některých druhů živí také banány, marmeládou, mlékem, žloutky či klobásami.[41]
Nepřátelé

Pavouci mají rozličné predátory, pro mnohé z nich představují důležitý či dokonce jediný zdroj potravy. Mnohé druhy ptáků intenzivně loví pavouky, kteří mohou tvořit i tři čtvrtiny celkového objemu jejich kořisti. Rejsci, žáby, ještěrky, někteří hadi a jiní menší obratlovci rovněž zařazují pavouky do svého jídelníčku. Důležitými predátory respektive parazitoidy pavouků jsou zástupci hmyzího řádu blanokřídlých. Jde například o některé lumky z čeledi Ichneumonidae, kutilky (například rod Sceliphron) a především hrabalky (Pompilidae). Využívají pavouky jako zásobárnu živin pro své larvy. Nepřáteli pavouků mohou být i jiní pavouci. Ostníci se například specializují na lov síťových pavouků, skákavky rodu Portia loví širokou škálu jiných pavouků. Pavouky dále napadají a paraziticky využívají některé houby (Gibellula), hlístice, kulatěnky, pakudlanky jižní a další.[42]
Obrana

Existují silné důkazy podporující myšlenku, že zbarvení pavouků slouží jako maskování, které je chrání před hlavními predátory v podobě ptáků a parazitoidních vos (kutilek, hrabalek), kteří mají dobré barevné vidění. Mnoho druhů pavouků je zbarveno tak, aby zapadlo do okolí, a někteří mají rušivé zbarvení, které predátorům znemožňuje rozeznat jejich obrysy. U několika málo druhů, mezi které patří například Theridion grallator, se vyskytuje mnoho různých typů zbarvení, což má za účel zmást predátory, kteří tak mohou mít problémy tyto druhy rozeznat. Většina pavouků není dostatečně nebezpečná či nepoživatelná na to, aby se u nich vyskytovalo varovné zbarvení. Několik druhů se silným jedem či velkými chelicerami má ale skvrny varovných barev, které ukazují, když se cítí ohroženi.[37][43]
Mnoho zástupců čeledi Theraphosidae má na zadečku dráždivé chloupky, které svýma nohama hází na predátory. Tyto chloupky mají na konci háčky, které dokážou silně dráždit, i když podle všeho neobsahují žádný jed.[44] Několik pavouků se chrání před vosami tak, že do pavučiny zaplétají zvláště odolná vlákna, s jejichž protrhnutím mají vosy problémy, takže pavouk má čas na útěk.[45] Pavouci druhu Carparachne aureoflava žijící v Namibské poušti unikají parazitoidním vosám tak, že se převrátí na bok a skutálí se z písečných dun.[46]
Sociální pavouci
Několik druhů, které vytvářejí pavučiny, žije spolu ve velkých koloniích a vykazuje sociální chování, i když ne tak důsledné jako u sociálního hmyzu. Například Anelosimus eximius z čeledi snovačkovití (Theridiidae) vytváří kolonie čítající až 50 000 jedinců.[47] V rodu Anelosimus se vyskytuje několik sociálních druhů: všechny druhy žijící v Americe jsou sociální a druhy na Madagaskaru jsou alespoň částečně sociální.[48] U různých druhů patřících do stejné čeledi se sociální chování může vyvinout nezávisle na sobě. Například druh Theridion nigroannulatum patří do rodu, ve kterém se žádný jiný sociální druh nevyskytuje, a přitom vytváří kolonie až o několika tisících jedincích, kteří spolupracují při lovu potravy, kterou mezi sebou sdílejí.[49] Mezi další sociální pavouky patří například několik druhů rodu Philoponella (čeleď pakřižákovití), pokoutník společenský (Agelena consociata, čeleď pokoutníkovití) a Mallos gregalis (čeleď cedivečkovití).[50] Draví sociální pavouci musí chránit svou potravu před kleptoparazity a větší kolonie jsou v plnění tohoto úkolu úspěšnější.[51] Býložravý pavouk Bagheera kiplingi žije v malých koloniích, které chrání vajíčka a mláďata.[40] Dokonce i snovačky (Latrodectus), které jsou známé kanibalismem, v zajetí vytvářejí malé kolonie, ve kterých jedinci sdílí pavučiny a společně žerou.[52]
Druhy pavučin

Neexistuje žádný pevný vztah mezi klasifikací pavouků a typem pavučin, které vytvářejí: druhy ve stejném rodu mohou vytvářet velmi podobné či výrazně odlišné pavučiny. Totéž platí o chemickém složení pavoučích vláken. Vzájemně podobné techniky vytváření pavučin nejčastěji vznikají konvergencí. Většina pavouků snová nekruhové pavučiny, přičemž se předpokládá, že kruhové pavučiny jsou evolučně starší.[53]
Kruhové pavučiny

Zhruba polovina potenciální kořisti, která se zachytí do kruhové pavučiny, unikne. Pavučina má tři funkce: zachycení kořisti; pohlcení její hybnosti, aniž by se roztrhla; a zadržení kořisti. Neexistuje ideální struktura pavučiny. Větší mezery mezi vlákny například zvětšují plochu pavučiny a tím pádem zvyšují šanci na zachycení kořisti, na druhou stranu ale snižují šanci na její zadržení; menší mezery, větší lepkavé kapky a silnější vlákna zase kořist lépe zadrží, ale ta si tím spíše pavučiny všimne a vyhne se jí. Neexistují žádné pevné rozdíly mezi kruhovými pavučinami používanými přes den a těmi používanými přes noc.[53]
Místo, kde pavouci číhají, se většinou nachází přímo nad středem pavučiny, protože pavouci se dolů pohybují rychleji než nahoru. Pokud existuje nějaký směr, kterým pavouk může uniknout do bezpečí před svými predátory, většinou je vyčkávací místo pavouka vychýleno tímto směrem.[53]
Celkem běžné jsou horizontální sítě, i když jsou méně efektivní při zachycování a zadržování kořisti a více náchylné k poškození deštěm a padajícími objekty. Na druhou stranu ale mají některé výhody: nejsou tolik poškozovány větrem; kořist letící vzhůru je nevidí tak dobře kvůli slunci v pozadí; a lze je rozkmitat tak, aby chytily hmyz v pomalém horizontálním letu.[53]
Pavouci často na pavučiny přidělávají silné pásky vláken, takzvaná stabilimenta. Podle výzkumníků se v takto ozdobených pavučinách za hodinu zachytí více kořisti.[54] Jak ukázala laboratorní studie, když pavouci cítí přítomnost predátorů, vytváření stabiliment omezují.[55]
Existuje několik neobvyklých variant kruhových pavučin, z nichž se mnohé vyvinuly konvergentně. Patří mezi ně například pavučiny připevněné k vodní hladině, možná kvůli chytání hmyzu plovoucího ve vodě; pavučiny s větvičkami uprostřed, za kterými se pavouk může schovat před predátory; či pavučiny podobné žebříku, které se zdají vhodné na chytání můr. Význam mnoha variací však zůstává nejasný.[53]
V roce 1973 vzala kosmická loď Skylab 3 pavouky vytvářející kruhové sítě do kosmického prostoru za účelem otestování snovacích dovedností ve stavu beztíže. Zpočátku pavouci snovali nekvalitní pavučiny, ale brzy se novému prostředí přizpůsobili.[56]
Pavučinový vak

Pavouci z čeledi snovačkovití snovají nepravidelné, propletené trojrozměrné pavučiny, kterým se také říká pavučinové vaky. Zdá se, že existuje evoluční trend směřující k redukci použitých lepkavých vláken, což u některých druhů vyústilo až v jejich úplnou absenci. Snování pavučinových vaků je méně stereotypní než snování kruhových pavučin a může trvat až několik dnů.[57]
Ostatní druhy pavučin
Plachetnatkovití (Linyphiidae) vytvářejí horizontální hrbolaté „plachetky“, nad kterými jsou smotaná zastavovací vlákna. Když do těchto vláken narazí hmyz, spadne do plachetky, kde se zachytí v lepkavých vláknech. Pavouk na něj potom zaútočí zespoda.[58]
Evoluce
.jpg.webp)
I když je fosilní záznam pavouků považován za slabý,[59] bylo z fosilií popsáno téměř 1000 druhů.[60] Protože těla pavouků jsou křehká, většina z fosilizovaných pavouků byla zachována v jantaru.[60] Nejstarší známá fosilie členovce v jantaru pochází z rané křídy z doby před 130 miliony lety. Fosilie neslouží pouze ke zkoumání anatomie pavouků, ale v jantaru byli zachyceni také pavouci při páření, lovení zvěře, snování vláken a možná i při péči o mláďata. Zachovaly se i zkameněliny kokonů s vajíčky a pavučin se zachycenou kořistí;[61] nejstarší známý fosilní záznam pavučiny je starý 100 milionů let.[62]
Nejstarší známý pavoukovec je Palaeotarbus jerami z období siluru z doby asi před 420 miliony lety. Měl trojúhelníkovou hlavohruď, členitý zadeček, osm nohou a makadla.[63] Attercopus fimbriunguis z devonu z doby asi před 386 miliony lety má nejstarší známé spigoty vytvářející pavučinová vlákna a byl tudíž označen za pavouka.[64] Tyto spigoty se ale možná nacházely na spodní straně zadečku namísto na snovacích bradavkách, jejichž pohyblivost je důležitá při stavění pavučin. Proto Attercopus a jemu podobný permský pavoukovec Permarachne možná nebyli pravými pavouky, protože pavučinová vlákna využívali spíše na stavbu hnízd a kokonů než na vytváření pavučin.[65] Největším známým fosilním pavoukem je Mongolarachne jurassica z doby asi před 165 miliony lety, který byl objeven ve Vnitřním Mongolsku.[66] Délka jeho těla dosahuje téměř 25 mm a délka nohou asi 56,5 mm.[67]
Několik pavouků z karbonu je z primitivního podřádu sklípkoši (Mesothelae), z něhož do dnešní doby přežila pouze čeleď sklípkošovití (Liphistiidae).[64] Ačkoliv v období permu před 299 až 251 miliony lety probíhala rychlá diverzifikace létajícího hmyzu, zachovalo se z tohoto období velmi málo fosilií pavouků.[64]
Dvě hlavní skupiny moderních pavouků, sklípkani (Mygalomorphae) a dvouplicní (Araneomorphae), se poprvé objevily v triasu před více než 200 miliony lety. Někteří z pavouků z tohoto období se zdají být z čeledi sklípkancovití (Hexathelidae) a jejich snovací bradavky byly zřejmě uzpůsobené k vytváření pavučin ve tvaru trychtýřů. Dvouplicní zahrnují velkou většinu současných pavouků, mezi než patří pavouci snovající kruhové sítě. Z jury a křídy se zachovalo velké množství fosilizovaných pavouků včetně zástupců mnoha současných čeledí.[64]
Taxonomie
Pavouci jsou rozděleni do dvou podřádů, sklípkoši a Opisthothelae, přičemž Opisthothelae zahrnuje dva infrařády: sklípkani a dvouplicní.[pozn. 2] Je známo přes 40 000 žijících druhů pavouků, které arachnologové rozřazují do zhruba 128 čeledí a zhruba 4200 rodů.[2][68]
| Rozmanitost[2][68] | Vlastnosti | ||||||
| Podřád | Druhy | Rody | Čeledi | Členitá horní strana zadečku[69] | Ganglia v zadečku !! Snovací bradavky[69] !! Směr pohybu chelicer[8] | ||
|---|---|---|---|---|---|---|---|
| sklípkoši | 116 | 8 | 1 | ano | ano | čtyři páry, u některých druhů je jeden pár spojený, pod středem zadečku | dolů a dopředu |
| sklípkani | 2900 | 350 | 20 | ne, pouze některé fosilie | ne | jeden, dva nebo tři páry na konci zadečku | |
| dvouplicní | 44 000 | 3700 | 96 | ze stran do středu jako klepeta | |||
Sklípkoši

Jediní žijící zástupci z podřádu sklípkoši jsou z čeledi sklípkošovití a vyskytují se pouze v jihovýchodní Asii, Čínské lidové republice a Japonsku.[68] Většina sklípkošů si vytváří doupata s poklopem lemovaná pavoučími vlákny, i když někteří zástupci rodu Liphistius stavějí zakryté pavučinové trubice s druhým poklopem, kterým můžou uniknout v případě nouze. Zástupci rodu Liphistius natahují vlákna mimo tunely, aby lépe dokázali zaznamenat blížící se kořist, zatímco zástupci rodu Heptathela se spoléhají pouze na vnitřní senzory vibrací a vlákna nenatahují.[71] Pavouci z rodu Heptathela nemají žádné jedové žlázy, i když v drápcích mají jejich vývody.[72]
Sklípkani

Sklípkani, kteří se poprvé objevili v triasu,[64] jsou většinou mohutní a chlupatí a mají velké silné chelicery a drápky.[68] Většina z nich tráví většinu života v doupatech, ale někteří snovají pavučiny. Na rozdíl od dvouplicních ale nedokáží produkovat takový druh pavoučího vlákna, který se okamžitě lepí na další vlákna či na jiné povrchy, a toto jim vytváření pavučiny ztěžuje.
Kromě členovců se živí také žábami, ještěrkami a šneky.[73]
Dvouplicní

Dvouplicní zahrnují asi 90 % pavoučích druhů a patří mezi ně například křižákovití, slíďákovití a skákavkovití,[68] jakožto i jediný známý býložravý pavouk, Bagheera kiplingi.[40] Vyznačují se drápky orientovanými proti sobě, na rozdíl od sklípkanů, jejichž drápky jsou téměř rovnoběžné.[74]
Pavouci a lidé
Pavoučí kousnutí
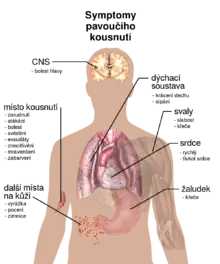
Většina pavouků napadá lidi jen při sebeobraně a jen málo kousnutí způsobuje horší následky než bodnutí komára či včely.[76] Většina těch se silnějším jedem, jako jsou například snovačky (mezi které patří i známá snovačka jedovatá) a pavouci z rodu Loxosceles, je plachá a kousá pouze tehdy, když se cítí ohrožená.[77][78] Sklípkanec jedovatý má agresivní obrannou taktiku a jeho jed způsobil úmrtí minimálně 13 lidí.[79] Z 20. století existuje asi 100 věrohodných nahlášených případů úmrtí na pavoučí kousnutí.[80]
Užitek pro lidi

Vaření sklípkani představují pro Kambodžany a pro indiány kmene Piaroa z jižní Venezuely pochoutku.[81][82]
Pavoučí jedy by mohly být šetrnější variantou k běžným pesticidům, protože jsou smrtelné pro hmyz, ale většina z nich je neškodná pro obratlovce. Vhodným zdrojem by mohli být australští sklípkanci jedovatí, protože většina hmyzích druhů na jejich jed nemá vybudovanou imunitu a sklípkancům jedovatým se daří v zajetí a jed z nich lze lehce dostat. Jednotlivé škůdce by možná bylo možné zaměřit pomocí genetického inženýrství tak, že by se geny zodpovědné za tvorbu pavoučích toxinů implantovaly do virů, které napadají například populace černopásky bavlníkové.[83]
Pavoučí jed má potenciální využití v lékařství například při léčbě srdeční arytmie,[84] Alzheimerovy choroby,[85] mozkové mrtvice[86] a erektilní dysfunkce.[87]
Protože pavoučí hedvábí je zároveň lehké a pevné, probíhají pokusy o jeho umělou syntézu. Geny pro tvorbu proteinů pavoučích vláken jsou také pomocí metod genetického inženýrství přenášeny do genomu dalších potenciálních producentů, například některých rostlin, bakterií, kvasinek či koz.[88][89]
Arachnofobie
Arachnofobie je specifická fobie – abnormální strach z pavouků nebo čehokoliv připomínajícího pavouky, jako jsou například pavučiny a pavoučí tvary. Je jednou z nejběžnějších specifických fobií[90][91] a některé statistiky uvádějí, že se projevuje u 50 % žen a 10 % mužů.[92] Může se jednat o přehnanou formu instinktivní reakce, která pomohla prvním lidem k přežití,[93] či o kulturní jev běžný hlavně v evropských společnostech.[94]
Pavouci v kultuře

Pavouci byli předmětem zájmu příběhů a mytologií různých kultur.[95] Kvůli své technice lovu spočívající ve vytváření sítí a čekání na kořist symbolizují trpělivost, na druhou stranu kvůli jedovatým (a někdy smrtelným) kousnutím symbolizují krutost.[96] Snování pavučin také zapříčinilo spojení pavouků s mýty o stvoření, protože pavouci jako by si vytvářeli vlastní světy.[97]
Močikové ze starodávného Peru vzývali přírodu a často ve svém umění zobrazovali pavouky.[98]
Odkazy
Poznámky
- Rozhovor s jedním z tvůrců červeného seznamu o příčinách jeho aktualizace a diskuse proč někteří pavouci z české přírody mizí je volně dostupný na stránkách Českého rozhlasu: (pořad Planetárium, 12. 9. 2015)[nedostupný zdroj].
- Skupina Opisthothelae je někdy považována za nezařazený klad a někdy za podřád pavouků. V tom druhém případě se sklípkani a dvouplicní považují za infrařády, jinak za podřády.
Reference
V tomto článku byl použit překlad textu z článku Spider na anglické Wikipedii.
- SEBASTIAN, P. A.; PETER, K. V. Spiders of India. India: Universities Press, 2009. 614 s. Dostupné online. ISBN 978-81-7371-641-6. S. 2. (anglicky)
- PLATNICK, Norman I. The World Spider Catalog, version 13.0 [online]. American Museum of Natural History, 2012 [cit. 2021-03-16]. Dostupné online. (anglicky)
- FOELIX, Rainer F. Biology of Spiders. New York: Oxford University Press, 1996. Dostupné online. ISBN 0-19-509593-6. S. 3. (anglicky)
- ŘEZÁČ, Milan; KŮRKA, Antonín; RŮŽIČKA, Vlastimil, HENEBERG Petr:. Red List of Czech spiders: 3rd edition, adjusted according to evidence-based national conservation priorities.. S. 645–666. Biologia [online]. 2015. Roč. 70, čís. 5, s. 645–666. Dostupné online. DOI 10.1515/biolog-2015-0079. (EN)
- MEEHAN, Christopher J.; OLSON, Eric J.; REUDINK, Matthew W., KYSER, T. Kurt; CURRY, Robert L. Herbivory in a spider through exploitation of an ant–plant mutualism. Current Biology. 2009, roč. 19, čís. 19, s. R892–3. DOI 10.1016/j.cub.2009.08.049. PMID 19825348. (anglicky)
- Ruppert, 554–555
- Ruppert, 518–522
- Ruppert, 571–584
- Ruppert, 559–564
- Ruppert, 565–569
- Ruppert, 527–528
- CODDINGTON, J. A.; LEVI, H. W. Systematics and Evolution of Spiders (Araneae). Annual Review of Ecology, Evolution, and Systematics. 1991, roč. 22, s. 565–592. Dostupné online. DOI 10.1146/annurev.es.22.110191.003025. (anglicky)
- CASAS, Jérôme. Spider Physiology and Behaviour: Physiology. [s.l.]: Academic Press, 2011. 239 s. Dostupné online. ISBN 9780123876683. S. 3–4. (anglicky) Google-Books-ID: RCs0uQLZqWMC.
- Ruppert, 529–530
- Ruppert, 531–532
- Ruppert, 532–537
- Ruppert, 578–580
- HARLAND, D. P.; JACKSON, R. R. "Eight-legged cats" and how they see — a review of recent research on jumping spiders (Araneae: Salticidae). Cimbebasia. 2000, roč. 16, s. 231–240. Dostupné v archivu pořízeném dne 2006-09-28. (anglicky) Archivováno 28. 9. 2006 na Wayback Machine
- BARNES, R. S. K., CALOW, P., OLIVE, P., GOLDING, D., SPICER, J. The Invertebrates: A Synthesis. Oxford: Blackwell Publishing, 2001. Dostupné online. ISBN 0-632-04761-5. Kapitola Invertebrates with Legs: the Arthropods and Similar Groups, s. 168. (anglicky)
- WEIHMANN, Tom, GÜNTHER, Michael; BLICKHAN, Reinhard. Hydraulic Leg Extension Is Not Necessarily the Main Drive in Large Spiders. The Journal of Experimental Biology. 2012-02-15, roč. 215, čís. 4, s. 578–583. Dostupné online [cit. 2013-04-27]. ISSN 1477–9145 0022-0949, 1477–9145. DOI 10.1242/jeb.054585. (anglicky)
- PARRY, D. A., BROWN, R. H. J. The Hydraulic Mechanism of the Spider Leg. Journal of Experimental Biology. 1959, roč. 36, čís. 2, s. 423–433. Dostupné online [cit. 2013-04-27]. (anglicky)
- Ruppert, 325–349
- Tarantulas Shoot Silk from Their Feet. Science Daily [online]. 2011-05-17 [cit. 2013-04-28]. Dostupné online. (anglicky)
- VOLLRATH, F., KNIGHT, D. P. Liquid crystalline spinning of spider silk. Nature. 2001, roč. 410, čís. 6828, s. 541–548. DOI 10.1038/35069000. PMID 11279484. (anglicky)
- Ruppert, 537–539
- KNOFLACH, B., van HARTEN, A. Tidarren argo sp. nov (Araneae: Theridiidae) and its exceptional copulatory behaviour: emasculation, male palpal organ as a mating plug and sexual cannibalism. Journal of Zoology. 2001, roč. 254, čís. 4, s. 449–459. DOI 10.1017/S0952836901000954. (anglicky)
- ANDRADE, Maydianne C. B. Risky mate search and male self-sacrifice in redback spiders. Behavioral Ecology. 2003, roč. 14, čís. 4, s. 531–538. DOI 10.1093/beheco/arg015. (anglicky)
- FOELIX, R. F. Biology of Spiders. New York: Oxford University Press US, 1996. Dostupné online. ISBN 0-19-509594-4. Kapitola Reproduction, s. 176–212. (anglicky)
- Ruppert, 523–524
- FOELIX, Rainer F. Biology of Spiders. New York: Oxford University Press, 1996. Dostupné online. ISBN 0-674-07431-9. S. 232–233. (anglicky)
- LEVI, Herbert W.; LEVI, Lorna R. Spiders and their Kin. New York: Golden Press, 2001. Dostupné online. ISBN 1582381569. S. 20 a 44. (anglicky)
- SCHÜTZ, D.; TABORSKY, M. Adaptations to an aquatic life may be responsible for the reversed sexual size dimorphism in the water spider, Argyroneta aquatica. Evolutionary Ecology Research. 2003, roč. 5, čís. 1, s. 105–117. Dostupné v archivu pořízeném dne 2008-12-16. (anglicky)
- CODDINGTON, J.; SOBREVILA, C. Web manipulation and two stereotyped attack behaviors in the ogre-faced spider Deinopis spinosus Marx (Araneae, Deinopidae). Journal of Arachnology. 1987, roč. 15, s. 213–225. Dostupné online [cit. 2008-10-11]. (anglicky)
- EBERHARD, W. G. Aggressive Chemical Mimicry by a Bolas Spider. Science. 1977, roč. 198, čís. 4322, s. 1173–1175. Dostupné online [cit. 2008-10-10]. DOI 10.1126/science.198.4322.1173. (anglicky)[nedostupný zdroj]
- EBERHARD, W. G. The Natural History and Behavior of the Bolas Spider, Mastophora dizzydeani sp. n. (Araneae). Psyche. 1980, roč. 87, čís. 3–4, s. 143–170. Dostupné online [cit. 2008-10-10]. DOI 10.1155/1980/81062. (anglicky)
- YEARGAN, K. V.; QUATE, L. W. Adult male bolas spiders retain juvenile hunting tactics. Oecologia. 1997, roč. 112, čís. 4, s. 572–576. Dostupné online. DOI 10.1007/s004420050347. (anglicky)
- OXFORD, G. S.; GILLESPIE, R. G. Evolution and Ecology of Spider Coloration. Annual Review of Entomology. 1998, roč. 43, s. 619–643. Dostupné online. DOI 10.1146/annurev.ento.43.1.619. PMID 15012400. (anglicky)
- WILCOX, S.; JACKSON, R. The Cognitive Animal: Empirical and Theoretical Perspectives on Animal Cognition. Příprava vydání Bekoff, M., Allen, C. a Burghardt, G. M.. Cambridge: MIT Press, 2002. Dostupné online. ISBN 0-262-52322-1. Kapitola Jumping Spider Tricksters, s. 27–34. (anglicky)
- MCLVER, J. D.; STONEDAHL, G. Myrmecomorphy: Morphological and Behavioral Mimicry of Ants. Annual Review of Entomology. 1993, roč. 38, s. 351–377. DOI 10.1146/annurev.en.38.010193.002031.
- MEEHAN, C. J., OLSON, E. J., CURRY, R. L. Exploitation of the Pseudomyrmex–Acacia mutualism by a predominantly vegetarian jumping spider (Bagheera kiplingi) [online]. 93rd ESA Annual Meeting, 2008-08-21 [cit. 2008-10-10]. Dostupné v archivu pořízeném dne 2019-12-01. (anglicky)
- JACKSON, R. R., POLLARD, Simon D., NELSON, Ximena J., EDWARDS, G. B., BARRION, Alberto T. Jumping spiders (Araneae: Salticidae) that feed on nectar. Journal of Zoology. 2001, roč. 255, s. 25–29. Dostupné online. DOI 10.1017/S095283690100108X. (anglicky)
- KŮRKA, Antonín, a kol. Pavouci České republiky. Praha: Academia, 2015. S. 55–58.
- Different smiles, single species [online]. University of California Museum of Paleontology [cit. 2008-10-10]. Dostupné online. (anglicky)
- COOKE, J. A. L., ROTH, V. D., MILLER, F. H. The urticating hairs of theraphosid spiders. American Museum novitates. American Museum of Natural History. Čís. 2498. Dostupné online [cit. 2008-10-11]. (anglicky)
- BLACKLEDGE, T. A.; WENZEL, J. W. Silk Mediated Defense by an Orb Web Spider against Predatory Mud-dauber Wasps. Behaviour. 2001, roč. 138, čís. 2, s. 155–171. DOI 10.1163/15685390151074357. (anglicky)
- ARMSTRONG, S. Fog, wind and heat — life in the Namib desert. New Scientist. 1990-07-14. Dostupné online [cit. 2008-10-11]. (anglicky)
- VOLLRATH, F. Eusociality and extraordinary sex ratios in the spider Anelosimus eximius (Araneae: Theridiidae). Behavioral Ecology and Sociobiology. 1986, roč. 18, čís. 4, s. 283–287. Dostupné online. DOI 10.1007/BF00300005. (anglicky)
- AGNARSSON, I.; KUNTNER, M. Madagascar: an unexpected hotspot of social Anelosimus spider diversity (Araneae: Theridiidae). Systematic Entomology. 2005, roč. 30, čís. 4, s. 575–592. DOI 10.1111/j.1365-3113.2005.00289.x. (anglicky)
- A New Independently Derived Social Spider with Explosive Colony Proliferation and a Female Size Dimorphism. Biotropica. 2006, roč. 38, čís. 6, s. 743–753. Dostupné online. DOI 10.1111/j.1744-7429.2006.00202.x. (anglicky)
- MATSUMOTO, T. Cooperative prey capture in the communal web spider, Philoponella raffray (Araneae, Uloboridae). Journal of Arachnology. 1998, roč. 26, s. 392–396. Dostupné online [cit. 2008-10-11]. (anglicky)
- CANGIALOSI, K. R. Social spider defense against kleptoparasitism. Behavioral Ecology and Sociobiology. 1990, roč. 27, čís. 1. DOI 10.1007/BF00183313. (anglicky)
- BERTANI, R., FUKUSHIMA, C. S., MARTINS, R. Sociable widow spiders? Evidence of subsociality in LatrodectusWalckenaer, 1805 (Araneae, Theridiidae). Journal of Ethology. 2008, roč. 26, čís. 2, s. 299. DOI 10.1007/s10164-007-0082-8. (anglicky)
- EBERHARD, W. G. Function and Phylogeny of Spider Webs. Annual Reviews in Ecology and Systematics. 1990, roč. 21, s. 341–372. Dostupné online [cit. 2008-10-15]. DOI 10.1146/annurev.es.21.110190.002013. (anglicky)
- HERBERSTEIN, M. E. Australian Journal of Zoology. Australian Journal of Zoology. 2000, roč. 48, čís. 2, s. 217–223. DOI 10.1071/ZO00007. (anglicky)
- LI, D.; LEE, W. S. Predator-induced plasticity in web-building behaviour. Animal Behaviour. 2004, roč. 67, čís. 2, s. 309–318. DOI 10.1016/j.anbehav.2003.06.011. (anglicky)
- THOMSON, Peggy; PARK, Edwards. Odd Tales from the Smithsonian [online]. [cit. 2008-07-21]. Dostupné online. (anglicky)
- AGNARSSON, I. Morphological phylogeny of cobweb spiders and their relatives (Araneae, Araneoidea, Theridiidae). Zoological Journal of the Linnean Society. 2004, roč. 141, čís. 4, s. 447–626. DOI 10.1111/j.1096-3642.2004.00120.x. (anglicky)
- SCHÜTT, K. Drapetisca socialis (Araneae: Linyphiidae): Web reduction — ethological and morphological adaptations. European Journal of Entomology. 1995, roč. 92, s. 553–563. Dostupné online [cit. 2008-10-19]. (anglicky)
- SELDEN, P. A., ANDERSON, H. M., ANDERSON, J. M. A review of the fossil record of spiders (Araneae) with special reference to Africa, and description of a new specimen from the Triassic Molteno Formation of South Africa. African Invertebrates. 2009, roč. 50, čís. 1, s. 105–116. Dostupné online. DOI 10.5733/afin.050.0103. (anglicky)
- DUNLOP, Jason A., PENNEY, David, TETLIE, Erik, ANDERSON, Lyall I. How many species of fossil arachnids are there?. The Journal of Arachnology. 2008, roč. 36, čís. 2, s. 267–272. Dostupné online. DOI 10.1636/CH07-89.1. (anglicky)
- PENNEY, D.; SELDEN, P. A. Spinning with the dinosaurs: the fossil record of spiders. Geology Today. 2007, roč. 23, čís. 6, s. 231–237. DOI 10.1111/j.1365-2451.2007.00641.x. (anglicky)
- HECHT, H. Oldest spider web found in amber [online]. New Scientist [cit. 2008-10-15]. Dostupné online. (anglicky)
- DUNLOP, J. A. A trigonotarbid arachnid from the Upper Silurian of Shropshire. Palaeontology. 1996, roč. 39, čís. 3, s. 605–614. Dostupné v archivu pořízeném dne 2008-12-16. (anglicky)
- VOLLRATH, F.; SELDEN, P. A. The Role of Behavior in the Evolution of Spiders, Silks, and Webs. Annual Review of Ecology, Evolution, and Systematics. 2007, roč. 38, s. 819–846. Dostupné v archivu pořízeném dne 2008-12-09. DOI 10.1146/annurev.ecolsys.37.091305.110221. (anglicky)
- SELDEN, P. A.; SHEAR, W. A. Fossil evidence for the origin of spider spinnerets. PNAS. Prosinec 2008, roč. 105, čís. 52, s. 20781–5. DOI 10.1073/pnas.0809174106. (anglicky)
- SELDEN, P. A., ChungKun Shih a Dong Ren. A golden orb-weaver spider(Araneae: Nephilidae: Nephila) from the Middle Jurassic of China. Biology Letters. 2011, roč. 7, čís. 5, s. 775–8. DOI 10.1098/rsbl.2011.0228. (anglicky)
- SELDEN, P. A., ChungKun Shih a Dong Ren. A giant spider from the Jurassic of China reveals greater diversity of the orbicularian stem group. Naturwissenschaften. 2013, roč. 100, čís. 12, s. 1171–81. DOI 10.1007/s00114-013-1121-7. (anglicky)
- CODDINGTON, J. A. Spiders of North America: an identification manual. Příprava vydání Ubick, D., Paquin, P., Cushing, P. E. a Roth, V. Poughkeepsie: American Arachnological Society, 2005. Dostupné online. ISBN 0-9771439-0-2. Kapitola Phylogeny and Classification of Spiders, s. 18–24. (anglicky)
- LEROY, J.; LEROY, A. Spiders of Southern Africa. Kapské Město: Struik, 2003. Dostupné online. ISBN 1-86872-944-3. Kapitola How spiders function, s. 15–21. (anglicky)
- ONO, H. New and Remarkable Spiders of the Families Liphistiidae, Argyronetidae, Pisauridae, Theridiidae and Araneidae (Arachnida) from Japan. Bulletin of the National Science Museum (of Japan), Series A. 2002, čís. 1, s. 51–60. Dostupné online. (anlicky)
- COYLE, F. A. Spiders—webs, behavior, and evolution. Stanford: Stanford University Press, 1986. Dostupné online. ISBN 0-8047-1203-4. Kapitola The Role of Silk in Prey Capture, s. 272–273. (anglicky)
- FORSTER, R. R.; PLATNICK, N. I. A review of the archaeid spiders and their relatives, with notes on the limits of the superfamily Palpimanoidea (Arachnida, Araneae). Bulletin of the American Museum of Natural History. 1984, roč. 178, s. 1–106. Dostupné online [cit. 2008-10-13]. (anglicky)
- Natural history of Mygalomorphae [online]. Agricultural Research Council of New Zealand [cit. 2008-10-13]. Dostupné v archivu pořízeném dne 2008-12-26. (anglicky)
- FOELIX, Rainer F. Biology of Spiders. New York: Oxford University Press, 2011. Dostupné online. ISBN 0-19-973482-8. S. 4–5. (anglicky)
- BROUHARD, Rod. Spider Bite: Symptoms, Pictures and Treatment [online]. About.com [cit. 2013-05-15]. Dostupné online. (anglicky)
- Spiders [online]. Illinois Department of Public Health [cit. 2008-10-11]. Dostupné online. (anglicky)
- VETTER, R. S.; BARGER, D. K. An infestation of 2,055 brown recluse spiders (Araneae: Sicariidae) and no envenomations in a Kansas home: implications for bite diagnoses in nonendemic areas. Journal of Medical Entomology. 2002, roč. 39, čís. 6, s. 948–51. DOI 10.1603/0022-2585-39.6.948. PMID 12495200. (anglicky)
- HANNUM, C.; MILLER, D. M. Widow Spiders [online]. Department of Entomology, Virginia Tech [cit. 2008-10-11]. Dostupné v archivu pořízeném dne 2008-10-18. (anglicky)
- Funnel web spiders [online]. Australian Venom Research Unit [cit. 2008-10-11]. Dostupné online.
- DIAZ, J. H. The Global Epidemiology, Syndromic Classification, Management, and Prevention of Spider Bites. American Journal of Tropical Medicine and Hygiene. 2004-08-01, roč. 71, čís. 2, s. 239–250. Dostupné online. PMID 15306718. (anglicky)
- RAY, N. Lonely Planet Cambodia. Melbourne: Lonely Planet Publications, 2002. ISBN 1-74059-111-9. S. 308. (anglicky)
- WEIL, C. Fierce Food. New York: Plume, 2006. Dostupné v archivu pořízeném dne 2011-05-11. ISBN 0-452-28700-6. (anglicky)
- Spider Venom Could Yield Eco-Friendly Insecticides [online]. National Science Foundation (USA) [cit. 2008-10-11]. Dostupné online. (anglicky)
- NOVAK, K. Spider venom helps hearts keep their rhythm. Nature Medicine. 2001, roč. 7, čís. 155, s. 155. DOI 10.1038/84588. PMID 11175840. (anglicky)
- LEWIS, R. J.; GARCIA, M. L. Therapeutic potential of venom peptides. Nature Reviews Drug Discovery. 2003, roč. 2, čís. 10, s. 790–802. Dostupné v archivu pořízeném dne 2008-12-16. DOI 10.1038/nrd1197. PMID 14526382. (anglicky) Archivováno 16. 12. 2008 na Wayback Machine
- BOGIN, O. Venom Peptides and their Mimetics as Potential Drugs. Modulator. Jaro 2005, čís. 19. Dostupné v archivu pořízeném dne 2008-12-09. (anglicky) Archivováno 9. 12. 2008 na Wayback Machine
- ANDRADE, E., VILLANOVA, F.; BORRA, P.; LEITE, K.; TRONCONE, L.; CORTEZ, I.; MESSINA, L.; PARANHOS, M.; CLARO, J. Penile erection induced in vivo by a purified toxin from the Brazilian spider Phoneutria nigriventer. British Journal of Urology International. 2008, roč. 102, čís. 7, s. 835–7. DOI 10.1111/j.1464-410X.2008.07762.x. PMID 18537953. (anglicky)
- HINMAN, M. B., JONES, J. A., LEWIS, R. W. Synthetic spider silk: a modular fiber. Trends in Biotechnology. 2000, roč. 18, čís. 9, s. 374–9. Dostupné v archivu pořízeném dne 2008-12-16. DOI 10.1016/S0167-7799(00)01481-5. PMID 10942961. (anglicky) Archivováno 16. 12. 2008 na Wayback Machine
- MENASSA, R., ZHU, H., KARATZAS, C. N., LAZARIS, A., RICHMAN, A., BRANDLE, J. Spider dragline silk proteins in transgenic tobacco leaves: accumulation and field production. Plant Biotechnology Journal. 2004, roč. 2, čís. 5, s. 431–8. DOI 10.1111/j.1467-7652.2004.00087.x. PMID 17168889. (anglicky)
- A Common Phobia [online]. phobias-help.com [cit. 2009-08-02]. Dostupné v archivu pořízeném dne 2009-08-02. (anglicky)
- FRITSCHER, Lisa. Spider Fears or Arachnophobia [online]. About.com, 2009-06-03 [cit. 2009-08-02]. Dostupné v archivu pořízeném dne 2009-06-19. (anglicky)
- The 10 Most Common Phobias — Did You Know? [online]. 10 Most Common Phobias [cit. 2009-08-02]. Dostupné online. (anglicky)
- FRIEDENBERG, J.; SILVERMAN, G. Cognitive Science: An Introduction to the Study of Mind. Thousand Oaks: SAGE, 2005. Dostupné online. ISBN 1-4129-2568-1. S. 244–245. (anglicky)
- DAVEY, G. C. L. The "Disgusting" Spider: The Role of Disease and Illness in the Perpetuation of Fear of Spiders. Society and Animals. 1994, roč. 2, čís. 1, s. 17–25. DOI 10.1163/156853094X00045. (anglicky)
- DE VOS, Gail. Tales, Rumors, and Gossip: Exploring Contemporary Folk Literature in Grades 7–12. Englewood: Libraries Unlimited, 1996. Dostupné online. ISBN 1-56308-190-3. S. 186. (anglicky)
- GARAI, Jana. The Book of Symbols. New York: Simon & Schuster, 1973. Dostupné online. ISBN 0-671-21773-9. (anglicky)
- DE LAGUNA, Frederica. American Anthropology: Papers from the American Anthropologist. Lincoln: University of Nebraska Press, 2002. Dostupné online. ISBN 0-8032-8280-X. S. 455. (anglicky)
- BERRIN, Kathleen. The Spirit of Ancient Peru: Treasures from the Museo Arqueologico Rafael Larco Herrera. New York: Thames & Hudson, 1997. (anglicky)
Literatura
- BUCHAR, Jan; KŮRKA, Antonín. Naši pavouci. Praha: Academia, 2001. 162 s.
- FOELIX, Rainer. Biology of Spiders. 2. vyd. New York: Oxford University Press, 1996. 330 s. Dostupné online.
- KŮRKA, Antonín, a kol. Pavouci České republiky. 1. vyd. Praha: Academia, 2015. 621 s. (Atlasy). ISBN 978-80-200-2384-1.
- MACEK, Rudolf. Pavouci - webové stránky se soupisem a fotografiemi pavouků České republiky [online]. 2006. Dostupné online.
- MARTINOVÁ, Zdena. Pavouk superpredátor: největší lovec ze všech. ABC mladých techniků a přírodovědců [online]. 2017, č. 9. 5. června 2017 [cit. 5. 10. 2017]. ISSN 0322-9580. Dostupné z: https://www.abicko.cz/clanek/precti-si-priroda/21581/pavouk-superpredator-nejvetsi-lovec-ze-vsech.html
- RUPPERT, E. E.; FOX, R. S.; BARNES, R. D. Invertebrate Zoology. Fort Worth: Brooks / Cole, 2004. (7). Dostupné online. ISBN 0-03-025982-7. (anglicky)
Externí odkazy
 Obrázky, zvuky či videa k tématu pavouci na Wikimedia Commons
Obrázky, zvuky či videa k tématu pavouci na Wikimedia Commons - Galerie pavouci na Wikimedia Commons
 Slovníkové heslo pavouk ve Wikislovníku
Slovníkové heslo pavouk ve Wikislovníku Encyklopedické heslo Pavouci v Ottově slovníku naučném ve Wikizdrojích
Encyklopedické heslo Pavouci v Ottově slovníku naučném ve Wikizdrojích Taxon Araneae ve Wikidruzích
Taxon Araneae ve Wikidruzích Téma Pavouk ve Wikicitátech
Téma Pavouk ve Wikicitátech- Aktuální představa o fylogenetickém stromu pavouků (dle Garrison NL, Rodriguez J, Agnarsson I, Coddington JA, Griswold CE, Hamilton CA, Hedin M, Kocot KM, Ledford JM, Bond JE. (2016) Spider phylogenomics: untangling the Spider Tree of Life. PeerJ 4:e1719 )