Buněčné jádro
Jádro (z lat. nucleus – jádro nebo oříšek, příp. řec. karyon – jádro[1]) je organela eukaryotických buněk, v níž je uložena většina genetického materiálu (DNA) buňky. Jedná se o váček obalený dvěma buněčnými membránami, který má v průměru 5–10 mikrometrů. Uvnitř se nachází chromatin, tedy DNA a různé přidružené bílkoviny, ale i další struktury (např. jadérko), kde probíhají různé enzymatické procesy související s DNA a RNA.
Jádro se vyskytuje v buňkách všech eukaryot (s několika drobnými výjimkami – např. lidská červená krvinka), tedy u všech rostlin, živočichů, hub nebo např. u prvoků. Někdy jich je dokonce v buňce více než jedno. O způsobu vzniku jádra existuje několik hypotéz a stále v tom není jasno.
Pro jeho nápadnost bylo jádro jednou z prvních struktur, které byly pozorovány pod mikroskopem, ale konkrétní biochemické a genetické děje uplatňující se v jádře jsou odhalovány až postupně. Mimo to, že jádro obsahuje DNA, má důležitou roli jako bariéra mezi místem transkripce (přepis DNA do RNA) a translace (z RNA do bílkovin).
Historie výzkumu

Jádro bylo první objevenou organelou vůbec. Autorem nejstaršího známého nákresu buňky s jádrem byl nizozemský přírodovědec Antoni van Leeuwenhoek (1632–1723). Jádro, které nazýval „lumen“, pozoroval v červených krvinkách lososa.[2] Později jádro popsal v roce 1804 Franz Bauer,[3] posléze do značných detailů Robert Brown. Poslední jmenovaný zkoumal stavbu květu orchidejí pod mikroskopem a ve vnější vrstvě květu si všiml struktury, kterou nazval areola či nucleus.[4] Jejich funkci nicméně ani on neznal. Až v roce 1838 předpověděl Matthias Schleiden, že jádro hraje roli v množení buněk: proto také pro jádro používal název cytoblast (z „cyto“–buňka, „blast“–vystavět). Věřil, že kolem „cytoblastů“ vznikají nové buňky. Odpůrcem Schleidena byl Franz Meyen, který se buněčným dělením zabýval a tvrdil, že mnohé buňky jádra mít nemusí. Později se ukázalo, že buňka vzniká vždy jen z buňky a tím pádem funkce jádra zůstala neznámá.[5]
V letech 1876–1878 publikoval Oscar Hertwig studie o průběhu oplodnění ježovek, přičemž si všiml, že během tohoto procesu dochází k fúzi jader vajíčka a spermie. Navrhl, že nový jedinec vzniká vždy z jedné oplozené buňky. Tomu se sice vzpouzel jeho současník Haeckel, nicméně později bylo to samé pozorováno u obojživelníků, měkkýšů a u rostlin. Jádro od té doby bylo považováno za centrum dědičnosti.[5] Oswald Avery, Colin MacLeod a Maclyn McCarty označili v roce 1944 ve svém slavném experimentu za nositelku genetické informace molekulu DNA.[6] V roce 1949 britský vědec H. G. Callan objevil jaderné póry.[7][8][9] Posléze bylo učiněno mnoho dalších důležitých objevů, například termín spliceozom, který označuje jaderný molekulární komplex sloužící k sestřihu RNA, byl poprvé použit v roce 1985 v laboratoři Johna Abelsona.[10][11] Podstata mnoha biochemických a molekulárních dějů v jádře však stále čeká na objevení.
Evoluční původ
Buněčné jádro je zřejmě evoluční novinka eukaryot. Velmi vzácně sice i některé bakterie (rod Gemmata) mohou mít DNA uzavřenou ve váčku, ale nejedná se o pravé eukaryotické jádro.[12] O vzniku pravého jádra eukaryot však existuje dlouhá řada hypotéz. Čtyři z nich se staly poměrně známými, nicméně žádná nepřesvědčila celou vědeckou obec.[13]
Podle jedné z nich se prostě z cytoplazmatické membrány (v místě, kde byla navázána DNA) odštěpily váčky, které následně splývaly a postupně obklopily genom.[14] Do jisté míry ve prospěch této hypotézy mluví skutečnost, že také některé planktomycetní bakterie mají jádro (Gemmata).[15]
Mnoho hypotéz o eukaryogenezi využívá endosymbióz několika primitivních buněk. Třeba podle syntrofické hypotézy ke vzniku jádra došlo takto: jakési bakterie se přiložily těsně k buněčné membráně dávné archebakterie a vzájemně mezi sebou splynuly. Archeální cytoplazmatická membrána se díky tomu mohla stát jadernou membránou nové eukaryotické buňky a cytoplazmatická membrána bakterií údajně představuje dnešní buněčnou membránu eukaryot.[16]
Další hypotézou je tzv. virová eukaryogeneze („tři viry, tři domény“). Tato velmi kontroverzní myšlenka hlásá, že vznik eukaryotické buňky se vším všudy je nutné přičítat virům. Důkazem je pro zastánce této hypotézy podoba v určitých rysech virů a eukaryot: mají lineární řetězce DNA, čepičku na mRNA a podobně. Virus mohl být pohlcen fagocytózou do buňky.[17] Podle některých autorů by předky jádra mohly být poxviry, jejichž DNA polymeráza vykazuje podobnost s totožným enzymem eukaryot.[18][19]
Konečně je možné zmínit ještě čtvrtou, tzv. „exomembránovou“ hypotézu, která byla vyslovena před několika lety. Podle ní endosymbiotické modely nejsou dostatečným vysvětlením pro vznik eukaryotického jádra. Jádro se podle této představy vyvinulo z jisté buňky, která si vyvinula druhou (vnější) cytoplazmatickou membránu. Tím pádem se ta vnitřní z membrán mohla stát jadernou membránou, zatímco její původní funkci přejala nová vnější membrána. V jaderné membráně vznikly komplikované pórovité komplexy, aby mohlo docházet k transportu různých struktur (jako jsou ribozomální podjednotky) z jádra do cytoplazmy či zpět.[20]
Parametry
Velikost a tvar
Jádro je dynamický útvar, který má různou velikost i tvar. Konkrétní rozměry jsou velmi rozmanité, průměrně se například uvádí 5–10 μm,[1][21] to jsou však pouze střední hodnoty. Jádro nervových buněk může mít pouhé 3, ale také 18 μm v průměru.[22] V typickém případě údajně jádro představuje asi 10% objemu buňky.[23] V rámci prvoků se udávají rozměry jádra mezi 1–100 μm.[24] V žabím vajíčku mohou mít jádra průměr klidně i 400 μm.[25] Co se týče rostlin, jedna studie udává, že nejmenší jádra v těle huseníčku rolního (Arabidopsis thaliana) najdeme ve svěracích buňkách průduchu, naopak suverénně největší jsou v buňkách trichomů.[26] Jádra se obecně zvětšují během svého buněčného cyklu, nejmenší jsou zpravidla po dokončení mitózy, největší na konci interfáze.[27] Je zajímavé, že po vložení jádra lidské buňky (HeLa) do vajíčka žáby, které je však mnohonásobně větší než lidská buňka, se toto jádro přizpůsobí velikosti vajíčka a zvětší svůj objem.[28] Známá je i skutečnost, že velikost jádra do jisté míry souvisí s ploidií (viz níže), tedy počtem chromozomových sad. Polyploidní jádra bývají výrazně větší.[24]
Jádra se často přizpůsobují i tvaru buňky.[25] V typickém případě jsou zřejmě jádra buněk kulovitého či mírně protáhlého tvaru. Výjimek je však spousta a například neutrofilní granulocyty mají lalokovitá, několikanásobně segmentovaná jádra,[27] která se dokonce mění s časem – nejprve jsou tyčkovitá, pak postupuje čím dál výraznější segmentace.[25] Také u rostlin se udávají nejen kulovitá, ale i protáhlá až tyčinkovitá jádra (např. v buňkách vodivých pletiv).[26] Prvoci s velkými jádry (makronukleus nálevníků) mají také tendenci mít laločnatá či segmentovaná jádra, zřejmě proto, aby lépe probíhala výměna látek mezi jádrem a cytoplazmou.[24]
Počet jader v buňce
Ve většině buněk se obyčejně vyskytuje jen jedno jádro (případně dvě těsně před rozdělením). Objevují se však výjimky, většinou u prvoků nebo hub.
Kromě těchto mnohojaderných útvarů ještě existují případy prvoků, které mají za normálního stavu dvě jádra. Rozlišují se homokaryotičtí prvoci, kteří mají více identických jader, a heterokaryotičtí prvoci, kteří mají v buňce dva jaderné typy (každý specializovaný pro jinou funkci). K homokaryotickým prvokům patří například améby Chaos a Pelomyxa, ale i rody Actinosphaerium a Opalina. Mnozí výtrusovci, jako např. kokcidie (Coccidiasina), mají alespoň vícejaderná životní stadia. Pro druhou zmíněnou skupinu, tedy heterokaryotické prvoky, je typický tzv. jaderný dualismus. Nálevníci (Ciliophora) a někteří dírkonošci (Foraminifera) mají totiž velký makronukleus (který je exprimován a neslouží k rozmnožování) a malý mikronukleus (který slouží k rozmnožování).[24] Také některé řasy (Caulerpa, Vaucheria, Botrydium) a buňky mléčnic jsou vícejaderné (polyenergidní).[29]
I u mnohobuněčných živočichů existují mnohojaderné buňky: patří k nim zejména syncytia a plazmodia. Syncytia vznikají splynutím několika jednojaderných buněk, plazmodia zase dělením jádra bez následného rozdělení buňky.[25] Například do kategorie mnohojaderných buněk patří osteoklasty, buňky kosterního svalu (rhabdomyocyty), ale i třeba buňky jaterní tkáně, chrupavek, kostní a epitelové přechodné tkáně v močovém měchýři.[25] Existují i buňky bezjaderné, takzvané neplnohodnotné, jako jsou červené krvinky savců, tedy i člověka. Také rostlinné buňky, které tvoří lýko (články sítkovic, sítkové buňky) nemají funkční jádra, ale jsou vyživovány buňkami okolními, které jádro mají.[29]
Počet sad chromozomů
Dalším parametrem, kterým je možno popsat jakékoliv jádro, je počet chromozomových sad, které se v nich nalézají. Mnoho eukaryot má haploidní jádra (pouze s jednou sadou genetické informace) – například válečovití bičíkovci (Volvocida), oxymonády (Oxymonadida), výtrusovci (Apicomplexa) či pravé hlenky (Myxogastrea).[24] Také stopkovýtrusné houby (Basidiomycota) a další zástupci říše hub mají téměř po celou dobu buněčného cyklu haploidní jádro.[30] U člověka jsou haploidní pouze pohlavní buňky, vajíčko a spermie, u rostlin všechny buňky gametofytu. Pro většinu buněk eukaryot je nicméně typické mít v jádře dvě sady genetické informace, tedy být diploidní. To platí namátkou pro mikronukley nálevníků (Ciliophora), opalinky (Opalinata), pro většinu těla cévnatých rostlin (jsou totiž z většiny tvořeny sporofytem) a pro téměř všechny buňky živočichů. Některá jádra však mohou být i polyploidní, tedy mít více než dvě sady genetické informace (např. makronukley nálevníků či celá těla některých polyploidních rostlin).[24]
Stavba
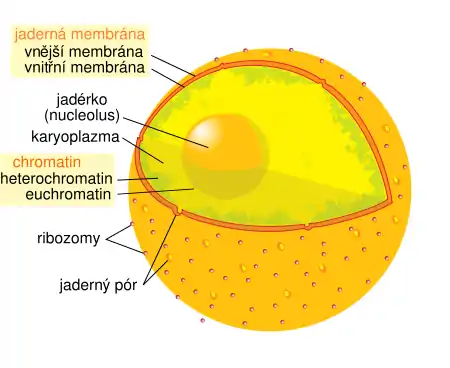
Jádro je váček obklopený dvěma fosfolipidovými membránami, tvořícími dohromady tzv. jadernou membránu (karyolemu). Ta je na vnitřní straně vyztužena systémem intermediárních filament, tzv. jadernou laminou. Uvnitř jádra se nachází karyoplazma, tedy vlastně cytoplazma s charakteristickým jaderným složením. Obsahuje především chromatin, tedy jadernou DNA a s ní asociované proteiny (histony a podobně). Kromě chromatinu lze v jádře najít ještě jadérka a další drobné struktury.
Jaderný obal
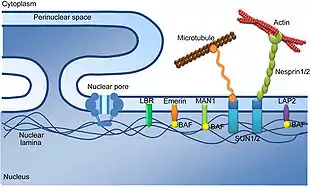
Jádro je ohraničeno dvojitým jaderným obalem („jadernou membránou“), v němž se nacházejí póry tvořené speciálními bílkovinami, které mají usnadnit a řídit transport specifických makromolekul, např. RNA. Z důvodu transportu mRNA k ribozómům a posttranslační úpravy bílkovin je jádro napojeno na drsné endoplazmatické retikulum. Obal jádra sestává ze dvou membrán a vrstvy mezi nimi:
- Vnitřní membrána, na kterou se váže chromatin (DNA a přídatné proteiny), jehož poloha se tím určuje. Také bývá přítomna vrstva filament, která zvyšuje pevnost. V typickém případě to je jaderná lamina, síť proteinů laminů silná 80–300 nm.[1] U měňavky Amoeba proteus se namísto klasické laminy nachází tzv. plástvová vrstva.[24]
- Vnější membrána připomíná membránu hrubého endoplasmatického retikula a stejně jako ona nese navázané ribozomy.[31]
- Mezimembránový prostor (perinukleární prostor, perinukleární cisterna) je ohraničen vnější a vnitřní membránou, pokračuje jako hrubé endoplazmatické retikulum. Ribozomy umístěné na vnější membráně syntetizují proteiny do tohoto prostoru. Šířka činí asi 20-40 nanometrů[31] (podle jiných zdrojů až 70 nm[1])
V jaderné membráně se nachází tzv. jaderné póry, které umožňují transport látek přes membránu. Co se týče obratlovců, jaderný pór má velikost přibližně 145 nanometrů v průměru a 80 nanometrů na délku (napříč membránou).[32] Na druhou stranu se skládají jaderné póry z „pouhých“ 30[33] (podle jiných zdrojů 50[34] V jaderném póru se odehrává prakticky veškerý membránový transport mezi cytosolem a jádrem. Tento transport je obousměrný a poměrně efektivní. Udává se, že za sekundu může přes nukleární pór projít až 1000 makromolekul.[35] Kupříkladu když buňka replikuje svoji DNA (v S-fázi), musí skrz každý z 3000 pórů projít každou minutu 100 histonů. Malé molekuly prochází pasivně, větší pomocí speciálních receptorů, které se s nimi asociují a přenášejí je.[23][33]
Chromatin
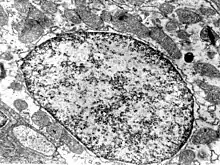
Většinu vnitřního prostoru jádra vyplňuje tzv. chromatin, tedy vlastně DNA a asociované bílkoviny (jako jsou třeba histony). Jaderná DNA, která slouží jako genetická informace pro syntézu většiny bílkovin v buňce, je tvořena několika lineárními řetězci, které se označují jako chromozomy. Zdravý člověk má v každé tělní buňce 46 takových lineárních molekul, jejichž celková délka je asi jeden metr.[25] Nejedná se však obvykle o náhodný spletenec, nýbrž o vysoce organizovaný komplex DNA a bílkovin. Na nejnižší úrovni vznikají z DNA a histonů tzv. nukleozomy, na vyšších úrovních nacházíme další typy organizace DNA. Kódující části chromatinu se nazývají euchromatin a bývají poněkud volněji smotané než heterochromatin, který tvoří nekódující DNA.
Jednotlivé chromozomy jsou obvykle pozorovány v M-fázi (při mitóze), kdy se buňka připravuje na dělení a jednotlivé chromozomy získávají svůj typický tvar (dvě chromatidy spojené v centromeře). V této době totiž je DNA tak hustě kondenzována, že jsou chromozomy viditelné i pod světelným mikroskopem. Většinu doby jsou však buňky v interfázi (období mezi děleními), kdy se chromozomy naopak rozmotávají, aby mohly být správně využívány pro běžnou činnost buňky. Je však zajímavé, že i v této době si udržují chromozomy svůj „prostor“ a příliš se navzájem nesplétají. Každá taková „komůrka“ se označuje chromozomální teritorium – zřejmě to hraje významnou epigenetickou roli v expresi genů.[36]
Jadérko
Jadérko (nucleolus) je malá vnitřní část buněčného jádra kulovitého tvaru, která obsahuje velké množství ribozomální RNA (rRNA). Vyskytuje se ve většině eukaryotických buněk. Jadérko je tvořeno satelitní DNA akrocentrických chromosomů. Jadérek bývá v typickém případě v jedné buňce jen několik, ale například v oocytech dochází k jejich namnožení do obrovských počtů, aby bylo později vajíčko schopné vyrobit dostatečné množství ribozomů.[37] Jadérko se v jádře vytváří kolem genů pro ribozomální RNA. Tyto geny jsou tak v jadérku transkribovány a následně se v jadérku zabudovávají do nově vznikajících ribozomů. Poté jsou jadernými póry transportovány ven do cytoplazmy.
Další struktury

Kromě jadérka jsou uvnitř buněčného jádra pravidelně pozorovány i další struktury, různých rozměrů a funkcí. Ač se někdy označují za organely, pomocí metody FRAP obvykle je zjištěno, že mezi nimi a karyoplazmou dochází k rychlé výměně materiálu (ostatně stejně tomu je i u jadérka). Jsou to tedy spíše lokální nahromadění molekulárních komplexů, jež spolu drží na základě slabých vazebných interakcí a zvyšují zde svou lokální koncentraci, což je vhodné pro rychlý průběh biochemických reakcí. K známějším tělískům tohoto typu patří například:[35]
- Cajalova tělíska: jsou obvykle menší než 1 mikrometr, v jádře jich bývá kolem desíti a obsahují vysokou koncentraci proteinu coilin (p80-coilin). Podílí na tvorbě snRNP, snoRNP a zrání telomerázy (její RNA komponenta je příbuzná snoRNA).[38]
- GEMs: obsahují protein SMN a hrají zřejmě důležitou roli v dozrávání snRNP komplexů.
- PIKA (OPT doména): zkratka pochází z anglického polymorphic interphase karyosomal association, je to zřejmě místo, kde dochází k vyhledávání a opravě chyb v DNA, ale jsou tam taky nahromaděny některé transkripční faktory. Během G fáze má až 5 mikrometrů.
- PML tělíska (též PODs či ND10): nachází se v jádře po 10–30 a jeho poruchy jsou dávány do souvislosti s některými typy leukémie.
- jaderné skvrny (speckles): jsou to zřejmě místa, kde dochází ke splicingu, neboť jsou v nich nahromaděny různé součásti sestřihové mašinerie. Jiné zdroje uvádí, že se zde tyto sestřihové faktor pouze skladují pro pozdější použití[35]
- paraspeckles ("tělíska v blízkosti jaderných skvrn") se vyskytují v bezprostřední blízkosti jaderných skvrn, jejich strukturní součást je dlouhá nekódující RNA NEAT1. Slouží k zachycení některých mRNA nesoucích informaci pro tvorbu proteinů využívaných pro odolání stresovým podmínkám, takové mRNA jsou bohatě modifikované RNA editací adenosin→inositol. Ve stresových podmínkách se odštípne modifikovaná oblast, na 3' konci se doplní poly(A) konec a zachycené mRNA je umožněno opustit jádro.[39]
Z různých menších, mikroskopem těžko pozorovatelných tělísek je v poslední době studován např. spliceozom, komplex, v němž probíhá splicing, tedy odstraňování intronů z pre-mRNA, [40] nebo "tělísko polycomb", což je nahloučení proteinů komplexu polycomb, který je zodpovědný za epigenetické umlčení řady genů pomocí methylace histonů a provází inaktivaci chromozomu X.[39] Otázka existence cytoskeletu v jádře je (vyjma laminů) vysoce kontroverzní, ačkoliv jisté vláknité struktury procházející skrz naskrz jádrem pozorovány byly.[41]
Funkce
Jádro je zcela zásadní součástí eukaryotických buněk, bez níž se nedá dlouhodobě přežívat. Například bezjaderné červené krvinky savců mají životnost obvykle přibližně pouhých 30 dní[25] (u člověka 120 dní). Je jasné, že hlavní funkcí jádra je uchovávat genetickou informaci, z níž je vystavěn život. Nabízí se však mimo jiné otázka, proč nestačí mít DNA v cytosolu, jako to ostatně běžně dělají bakterie a archebakterie – důvodem jsou zřejmě výhody, které přináší „kompartmentalizace“ buňky.
Genetická informace
Jádro obsahuje většinu genetické informace eukaryotických buněk. Jedná se o tzv. jadernou DNA, mimo ni existuje sice ještě mitochondriální DNA nebo plastidová DNA, ty však obvykle jsou řádově menší. V DNA se skrývají geny, tedy tzv. kódující úseky, které se v procesu transkripce nejprve musí přepsat v RNA. Tomu pomáhají různé jaderné enzymy, jako je RNA polymeráza, helikáza nebo topoisomeráza, a navíc různé transkripční faktory.[42] Vzniklá RNA se označuje jako primární transkript, je to surový materiál, který je třeba opatřit odolnými konci a sestříhat. Uvádí se tři základní kroky zahrnované pod posttranskripční modifikace: 5' guanosinová čepička, poly(A) konec a RNA sestřih (splicing). Poslední jmenovaný proces – sestřih – provádí spliceozom, enzymatická mašinérie, která vystřihává z primárních transkriptů introny. Podle toho, jak je vystřihne, mohou vznikat různé varianty též mRNA v procesu zvaném alternativní splicing.
Dalším krokem je obvykle export RNA z jádra. Ačkoliv některé typy RNA mohou pracovat v jádře, většina jich je jadernými póry posílána do cytoplazmy. To platí například pro všechny messenger RNA (mRNA). V cytoplazmě se mRNA váže na ribozomy a dochází k výrobě (translaci) polypeptidů (bílkovin).
Kompartmentalizace
Za tímto dlouhým slovem, které se však běžně používá, se skrývá zřejmě jedna z výhod eukaryotického jádra. Jaderný obal kolem jádra umožňuje, aby buňka vytvořila dva odlišné světy, cytoplazmu (vnitřek buňky) a karyoplazmu (vnitřek jádra). To je pro eukaryotickou buňku velice typické. Na obou stranách jaderného obalu tak může docházet k odlišným procesům. Pěkným příkladem je první krok glykolýzy, který je katalyzován enzymem hexokinázou. Je–li v buňce příliš fruktóza-1-fosfátu, hexokináza je přesunuta do jádra,[43] kde funguje jako represor, který zastavuje expresi (a tedy „výrobu“) enzymů nutných ke glykolýze. Celý systém je tedy negativní zpětnou vazbou.[44] Podobná je situace s transkripčními faktory (proteiny, které ovlivňují transkripci) – ty se také nalézají v cytosolu do doby, než jsou aktivovány. Ukazuje se, že nejlepší kontrolou transkripce je oddělit fyzicky transkripční faktory od DNA, aby se zamezilo byť slabé aktivaci genů ve chvíli, kdy to není žádoucí.[23]
Možná nejdůležitější je však schopnost jádra oddělit transkripci (přepis DNA v RNA) a translaci (přepis RNA v protein). Bakterie tyto dva děje provádí téměř současně, ale eukaryotická buňka musí po transkripci udělat ještě několik důležitých kroků předtím, než může mRNA být předána ribozomu. Zejména musí být proveden splicing (sestřih intronů). Kdyby k tomu nedošlo a pre-mRNA byla rovnou překládána v polypeptid, vznikaly by nefunkční bílkoviny.[45]
Jádro při buněčném dělení

Každá normální buňka musí mít plnohodnotné jádro. Zároveň však je žádoucí, aby se buňky mohly dělit. Aby se oba požadavky daly zkoordinovat, postupuje se v několika krocích. Nejprve se pouze v rámci tzv. S fáze jen namnoží obsah jádra (zejména celý genom), načež po kontrolním bodu vstoupí buňka do mitózy (M fáze). V prometafázi dojde k fosforylaci jaderné laminy (pomocí cyklin-dependentních kináz), načež se zcela rozpadne jaderná membrána. Díky tomu se mohou během metafáze navázat mikrotubuly z cytoplazmy na jednotlivé chromozomy, v anafázi dojde k rozchodu sesterských chromatid do protilehlých pólů buňky. Až v telofázi se opět obnovuje jaderná lamina i membrána, do níž se navíc znovu vkládají jaderné póry.[23]
Barvitelnost
Jádra poutala pozornost přírodovědců již od úsvitu mikroskopických technik. Někdy jsou dobře patrná pouhým okem, jindy je vhodné je barvit. Mezi klasická barviva patří zásaditý hematoxylin (fialově či hnědě zbarvená jádra), Giemsa, neutrální červeň, nilská modř nebo třeba safranin (červeně).[46][47] Takzvaná Feulgenova reakce zase využívá Shiffovo činidlo (kyselina fuchsinsiřičitá).[48] Na vzestupu je v posledních letech fluorescenční mikroskopie, která obvykle používá speciální barviva, jež se specificky váží na DNA: DAPI (diamidinofenylindol), Hoechst 33258, olivomycin, auramin-O nebo akriflavin.[49] Jindy se na jaderné proteiny navěšuje fluorescenční protein GFP.
Odkazy
Reference
V tomto článku byl použit překlad textu z článku Cell nucleus na anglické Wikipedii.
- Richard Jelínek, et al. Histologie embryologie [online]. 3. lékařská fakulta UK [cit. 2010-07-29]. Dostupné v archivu pořízeném z originálu.
- LEEUWENHOEK, A. van. Opera Omnia, seu Arcana Naturae ope exactissimorum Microscopiorum detecta, experimentis variis comprobata, Epistolis ad varios illustres viros. [s.l.]: J. Arnold et Delphis, A. Beman, Lugdinum Batavorum, 1719–1730. Citováno podle: Dieter Gerlach, Geschichte der Mikroskopie. Verlag Harry Deutsch, Frankfurt am Main, Germany, 2009. ISBN 978-3-8171-1781-9..
- HARRIS, H. The Birth of the Cell. New Haven: Yale University Press, 1999. Dostupné online.
- BROWN, Robert. On the Organs and Mode of Fecundation of Orchidex and Asclepiadea. Miscellaneous Botanical Works. 1866, roč. I, s. 511–514.
- CREMER, Thomas. Von der Zellenlehre zur Chromosomentheorie. Berlin, Heidelberg, New York, Tokyo: Springer Verlag, 1985. ISBN 3-540-13987-7. Online Version here Archivováno 11. 10. 2007 na Wayback Machine
- Avery O, MacLeod C, McCarty M. Studies on the chemical nature of the substance inducing transformation of pneumococcal types. Inductions of transformation by a desoxyribonucleic acid fraction isolated from pneumococcus type III. J Exp Med. 1944, roč. 79, čís. 2, s. 137–158. Dostupné v archivu pořízeném dne 2007-09-27. DOI 10.1084/jem.79.2.137. PMID 19871359.
- Callan, H. G., J . R . Randall, and S . G. Tomlin . 1949 . Nature (Land.) . 163 :280.
- FRANKE, W. W., U. Scheer, G. Krohne, E. D. Jarasch. The nuclear envelope and the architecture of the nuclear periphery. The Journal of Cell Biology. 1981-12-01, roč. 91, čís. 3, s. 39–50. ISSN 0021-9525.
- SOMMERVILLE, J. HAROLD GARNET CALLAN BA, DSc, FRS (Obituary) [online]. [cit. 2010-08-02]. Dostupné v archivu pořízeném dne 2006-09-25.
- BRODY, E, J Abelson. The "spliceosome": yeast pre-messenger RNA associates with a 40S complex in a splicing-dependent reaction. Science (New York, N.Y.). 1985-05-24, roč. 228, čís. 4702, s. 963–967. Dostupné v archivu pořízeném dne 2012-04-18. ISSN 0036-8075.
- GUTHRIE, Christine. From the Ribosome to the Spliceosome and Back Again. Journal of Biological Chemistry. 2010-01-01, roč. 285, čís. 1, s. 1–12. Dostupné v archivu pořízeném dne 2019-06-09. DOI 10.1074/jbc.X109.080580.
- LEE, Kuo-Chang, Rick Webb, John Fuerst. The cell cycle of the planctomycete Gemmata obscuriglobus with respect to cell compartmentalization. BMC Cell Biology. 2009-01-14, roč. 10, čís. 1, s. 4. Dostupné v archivu pořízeném dne 2009-02-09. ISSN 1471-2121. DOI 10.1186/1471-2121-10-4.
- Pennisi E. Evolutionary biology. The birth of the nucleus. Science. 2004, roč. 305, čís. 5685, s. 766–768. DOI 10.1126/science.305.5685.766. PMID 15297641.
- HAMPL, Vladimír. Protistologie - Vznik eukaryotické buňky - Eukaryogeneze [online]. natur.cuni.cz [cit. 2010-07-22]. Dostupné online.[nedostupný zdroj]
- FUERST, JA. Intracellular compartmentation in planctomycetes. Annu Rev Microbiol.. 2005, roč. 59, s. 299–328. DOI 10.1146/annurev.micro.59.030804.121258. PMID 15910279.
- MOREIRA, Lopez-Garcia. Symbiosis between methanogenic archaea and delta-proteobacteria as the origin of eukaryotes: the syntrophic hypothesis. Journal of Molecular Evolution. 1998-11, roč. 47, čís. 5, s. 517–30. Dostupné v archivu pořízeném dne 2012-01-27. ISSN 0022-2844. DOI 9797402.
- Bell PJ. (2001). "Viral eukaryogenesis: was the ancestor of the nucleus a complex DNA virus?" J Mol Biol Sep;53(3):251–256. PMID 11523012
- Takemura M. (2001). Poxviruses and the origin of the eukaryotic nucleus. J Mol Evol 52(5):419–425. PMID 11443345
- Villarreal L, DeFilippis V. A hypothesis for DNA viruses as the origin of eukaryotic replication proteins. J Virol. 2000, roč. 74, čís. 15, s. 7079–7084. Dostupné online. DOI 10.1128/JVI.74.15.7079-7084.2000. PMID 10888648.
- de Roos AD. The origin of the eukaryotic cell based on conservation of existing interfaces. Artif Life. 2006, roč. 12, čís. 4, s. 513–523.. DOI 10.1162/artl.2006.12.4.513. PMID 16953783.
- Chart of Actual Organelle Sizes [online]. [cit. 2010-07-26]. Dostupné v archivu pořízeném dne 2011-07-27.
- Archivovaná kopie. faculty.washington.edu [online]. [cit. 2010-07-26]. Dostupné v archivu pořízeném z originálu.
- ALBERTS, Bruce , et al. The Molecular Biology of the Cell. [s.l.]: Garland Science, 2002. (4th. ed). Dostupné online. ISBN 0-8153-3218-1.
- HAUSMANN, Klaus; HOLZMANN, Norbert. Protozoologie. Praha: Academia, 2003.
- PALEČEK, Jiří. Biologie buňky I. Základy mikroskopické cytologie. Praha: Karolinum, 1996.
- CHYTILOVA, EVA, JIRI MACAS, DAVID W. GALBRAITH. Green Fluorescent Protein Targeted to the Nucleus, a Transgenic Phenotype Useful for Studies in Plant Biology. Annals of Botany. 1999-06, roč. 83, čís. 6, s. 645–654. Dostupné online [cit. 2010-07-22]. ISSN 0305-7364. DOI 10.1006/anbo.1999.0866.
- WEBSTER, Micah, Keren L. Witkin, Orna Cohen-Fix. Sizing up the nucleus: nuclear shape, size and nuclear-envelope assembly. J Cell Sci. 2009-05-15, roč. 122, čís. 10, s. 1477–1486. Dostupné v archivu pořízeném dne 2010-03-16. DOI 10.1242/jcs.037333.
- JORGENSEN, Paul, Nicholas P. Edgington, Brandt L. Schneider, Ivan Rupes, Mike Tyers, Bruce Futcher. The Size of the Nucleus Increases as Yeast Cells Grow. Mol. Biol. Cell. 2007-09-01, roč. 18, čís. 9, s. 3523–3532. Dostupné v archivu pořízeném dne 2009-04-28. DOI 10.1091/mbc.E06-10-0973.
- VOTRUBOVÁ, Olga. Anatomie rostlin. Praha: Karolinum, 2001. ISBN 80-246-0367-5. (česky)
- KALINA, Tomáš; VÁŇA, Jiří. Sinice, řasy, houby, mechorosty a podobné organismy v současné biologii. Praha: Karolinum, 2005. 606 s. ISBN 80-246-1036-1.
- Oxford dictionary of biochemistry and molecular biology; revised edition. Příprava vydání R. Cammack et al. New York: Oxford university press, 2006. ISBN 0-19-852917-1.
- Tim Isgro, Lingling Miao. The Nuclear Pore Complex [online]. ks.uiuc.edu [cit. 2010-07-20]. Dostupné v archivu pořízeném z originálu.
- Anita H. Corbett. ENCYCLOPEDIA OF BIOLOGICAL CHEMISTRY, FOUR-VOLUME SET, 1-4. Příprava vydání Lennarz,W.J., Lane, M.D.. [s.l.]: [s.n.] Kapitola Nuclear Pores and Nuclear Import/Export.
- Alberts, Bruce , et al. The Molecular Biology of the Cell. [s.l.]: Garland Science, 2002. (4th. ed). ISBN 0-8153-3218-1.
- POLLARD, Thomas D; EARNSHAW, William C. Cell Biology. 2. vyd. [s.l.]: Saunders, 2007. ISBN 1416022554. S. 928.
- ZEGAŁO, Marta, Ewa Wiland, Maciej Kurpisz. Topology of chromosomes in somatic cells. Part 1. Postȩpy Higieny I Medycyny Doświadczalnej (Online). 2006, roč. 60, s. 331–342. Dostupné v archivu pořízeném dne 2012-04-18. ISSN 1732-2693.
- Thoru Pederson. Encyclopedia of Biological Chemistry - Vol 3. [s.l.]: [s.n.] Kapitola Nucleolus, Overview.
- MAO, YS.; ZHANG, B.; SPECTOR, DL. Biogenesis and function of nuclear bodies.. Trends Genet. Aug 2011, roč. 27, čís. 8, s. 295–306. DOI 10.1016/j.tig.2011.05.006. PMID 21680045.
- IP, JY.; NAKAGAWA, S. Long non-coding RNAs in nuclear bodies.. Dev Growth Differ. Jan 2012, roč. 54, čís. 1, s. 44–54. DOI 10.1111/j.1440-169X.2011.01303.x. PMID 22070123.
- ROBERT C. KING; WILLIAM D. STANSFIELD; PAMELA K. MULLIGAN. A Dictionary of Genetics, Seventh Edition. [s.l.]: Oxford University Press, 2006.
- LEWIN, Benjamin. Cells. [s.l.]: Jones & Bartlett Learning, 2007. Dostupné v archivu pořízeném dne 2012-11-14.
- NICOLINI, Claudio A. Genome Structure and Function: From Chromosomes Characterization to Genes Technology. [s.l.]: Springer, 1997. ISBN 0792345657.
- LEHNINGER, Albert L., David L. Nelson, Michael M. Cox. Lehninger principles of biochemistry. 3rd. vyd. New York: Worth Publishers, 2000. Dostupné online. ISBN 1-57259-931-6.
- Moreno F, Ahuatzi D, Riera A, Palomino CA, Herrero P. Glucose sensing through the Hxk2-dependent signalling pathway.. Biochem Soc Trans. 2005, roč. 33, čís. 1, s. 265–268. DOI 10.1042/BST0330265. PMID 15667322.
- GÖRLICH, Dirk, Ulrike Kutay. Transport between the cell nucleus and the cytoplasm. Ann. Rev. Cell Dev. Biol.. 1999, čís. 15, s. 607–660. Dostupné online. DOI 10.1042/BST0330265. PMID 10611974.
- BRYCHTOVÁ, Světlana. Histopatologický atlas. [s.l.]: Grada, 2008.
- BRUCKNER, Monica Z. Basic Cellular Staining [online]. Montana State University, Bozeman [cit. 2010-07-29]. Dostupné v archivu pořízeném z originálu.
- BERGER, Josef. BARVENÍ JÁDRA (Feulgenova reakce) [online]. [cit. 2010-07-29]. Dostupné v archivu pořízeném dne 2010-02-15.
- RAJU, N.B. How to stain and count nuclei. [online]. 2006 [cit. 2010-07-29]. Dostupné v archivu pořízeném z originálu.
Literatura
- Anglická
- BOOKS, L. L .C. Cell Nucleus: Nuclear Substructures, Chromosome, Nucleolus, Nuclear Pore, Chromatin, Nucleosol, Nucleoskeleton, Perinuclear Space, Nuclear Dots. [s.l.]: General Books, 2010. ISBN 9781157795520.
- LOMMEL, Alfons T. L. van. From cells to organs: a histology textbook and atlas. [s.l.]: Springer, 2002. ISBN 9781402072574.
- ALBERTS, Bruce, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter. Molecular Biology of the Cell. 4. vyd. [s.l.]: [s.n.] Dostupné online.
- Česká
- ALBERTS, Bruce, D. Bray, A. Johnson, J. Lewis, M. Raff, K. Roberts, P. Walter. Základy buněčné biologie, Úvod do molekulární biologie buňky. 2. vyd. [s.l.]: Espero Publishing, 2005. 740 s. ISBN 80-902906-2-0.
Externí odkazy
 Obrázky, zvuky či videa k tématu buněčné jádro na Wikimedia Commons
Obrázky, zvuky či videa k tématu buněčné jádro na Wikimedia Commons