Ploutvonožci
Ploutvonožci (Pinnipedia) jsou široce rozšířená skupina masožravých savců, kteří jsou uzpůsobení životu ve vodě. Moderní taxonomie je považuje za nezařazený klad v řádu šelem (Carnivora) a podřádu psotvární (Caniformia). K ploutvonožcům patří tři současné čeledi: tuleňovití (Phocidae), mrožovití (Odobenidae; s jediným žijícím zástupcem mrožem ledním) a lachtanovití (Otariidae). Počet druhů ploutvonožců se pohybuje kolem 33–36 v závislosti na tom, zda jsou některé taxony uznány za samostatný druh či nikoliv. Je známo více než 50 fosilních druhů. Ploutvonožci se oddělili od ostatních psotvárných asi před 50 miliony lety, k nejstarším známým prapředkům patří Enaliarctos a Puijila.
 Z levého horního rohu (ve směru hodinových ručiček): lachtan Forsterův, rypouš sloní (mládě), lachtan ušatý, mrož lední, tuleň kuželozubý | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Podkmen | obratlovci (Vertebrata) |
| Třída | savci (Mammalia) |
| Nadřád | placentálové (Placentalia) |
| Řád | šelmy (Carnivora) |
| Podřád | psotvární (Caniformia) |
| (nekategorizováno) | ploutvonožci (Pinnipedia) |
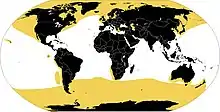 Areál běžného výskytu ploutvonožců (žlutě) Areál běžného výskytu ploutvonožců (žlutě)
| |
| Recentní nadčeledi a čeledi | |
| |
| Synonyma | |
Phocoidea, Illiger, 1811 | |
| Některá data mohou pocházet z datové položky. | |
Ploutvonožci mají hydrodynamická těla s končetinami přeměněnými v ploutve. Mrož a lachtanovití dokáží podsunout své zadní ploutve pod tělo, což jim pomáhá při pohybu po souši. K pohybu ve vodě používají hlavně své přední ploutve. Tuleňovití nemohou své zadní ploutve podsunout pod tělo, takže jejich pohyb na souši je nemotorný a pomalý; ve vodě jsou však obratnější než lachtani. Tuleni používají k pohybu ve vodě hlavně své zadní ploutve a střídavé pohyby zadní části těla. Zatímco lachtani mají ušní boltce, u tuleňů a mrožů boltce chybí. Velikost a váha ploutvonožců se pohybuje od cca 1,1 m a 55 kg (tuleň bajkalský) až po cca 6 m a 4 tuny (rypouš sloní). Rypouš sloní je nejen největší ploutvonožec, ale i největší zástupce řádu šelem na světě. Smysly ploutvonožců jsou dobře vyvinuty. Jejich sluch a zrak je výborně přizpůsoben vodnímu i vzdušnému prostředí a jejich vibrisy jim umožňují vnímat ve vodě i nepatrné vibrace, což jim pomáhá hlavně při pronásledování kořisti a patrně i v navigaci. Všichni ploutvonožci se umí potápět, některé druhy i na dobu přesahující hodinu a do hloubek přes jeden kilometr (rypouš sloní byl dokonce zaznamenán při ponoru přesahujícím 2 kilometry; pod vodou dokáže strávit až 2 hodiny). Všichni ploutvonožci disponují vrstvou podkožního tuku, která slouží jako termoizolace. Většina druhů má tělo pokryté hustou srstí, jen mroži mají na povrchu těla holou kůži s velmi řídkým ochlupením. Mroži jako jediní z ploutvonožců disponují dlouhými kly.
I když jsou ploutvonožci široce rozšíření, většina druhů žije v chladných vodách jižní a severní polokoule. Většinu času tráví ve vodě a na souš přichází pouze za účelem páření, kvůli vrhům, výchově mláďat, línání či při útěku před predátory jako jsou žraloci a kosatky. Žerou převážně bezobratlé živočichy, ryby, hlavonožce, některé agresivnější druhy jako tuleň leopardí loví i tučňáky či jiné ploutvonožce. Mroži se specializují na sbírání mlžů z oceánského dna. Většina ploutvonožců je polygynní, nicméně míra polygynie závisí na druhu. Samci ploutvonožců, kteří se páří na souši, mívají větší počet samic než druhy, které se páří na ledu. Největší počet samic na jednoho samce se vyskytuje u rypoušů, kde se samice shlukují v početných harémech. Reprodukční strategie samců se liší podle druhu a zahrnují lekový systém, obranu konkrétních samic před ostatními samci nebo obranu konkrétních teritorií před soky téhož druhu. Mláďata se rodí v jarních a letních měsících a jejich výchovu zajišťují pouze samice. Samice některých druhů se během laktace postí a jejich mláďata se osamostatňují poměrně brzy (již v rámci dnů), zatímco samice jiných druhů podnikají během laktace výpravy na moře za potravou a osamostatnění jejich mláďat tak trvá déle (až několik týdnů). Mrožice kojí i vychovávají svá mláďata na moři. Ploutvonožci dokáží vydávat řadu zvuků od hlasitého štěkotu (lachtani) přes zvuky připomínající gong (mroži) až po táhlé komplikované zvuky doplněné o ultrazvukovou frekvenci (tuleni Weddellovi).
Ploutvonožci byli loveni lidmi již od doby kamenné. Zvláště arktické národy dodnes ploutvonožce loví pro jejich maso, tuk i srst. V 18. a 19. století byla řada druhů komerčně lovena na pokraj vyhynutí. Tuleň karibský a lachtan japonský byli zcela vyhubeni. Ve 20. století začalo docházet k ochraně těchto mořských savců na národní i mezinárodní úrovni. Jedním z vrcholů mezinárodních ochranářských snah byla Úmluva o zachování antarktických tuleňů z roku 1972. Některé domorodé arktické národy (zejména na území Kanady) s lovením ploutvonožců pokračují i ve 21. století. K hlavním novodobým hrozbám pro tyto šelmy vedle lovu patří lidské aktivity jako jsou komerční rybolov, znečištění oceánů, těžební aktivity a současné klimatické změny.
Vědecká systematika
Taxonomie

K prvním popisům ploutvonožců došlo již v první první polovině 18. století. První vědecké popisy druhů byly uveřejněny v roce 1758 v 10. vydání Systema naturae švédského přírodopisce Carla Linného, který popsal mj. lachtana medvědího (Callorhinus ursinus), mrože ledního (Odobenus rosmarus), rypouše sloního (Mirounga leonina) či tuleně obecného (Phoca vitulina).[1] Linné nejdříve zařadil popsané druhy ploutvonožců do rodu Phoca a řádu Ferae, který v jeho pojetí zahrnoval masožravé šelmy[2] (v moderním pojetí jsou do této skupiny řazeni vedle šelem ještě luskouni). V pozdějších letech (1766) patrně pod vlivem francouzského přírodovědce Mathurina Jacquese Brissona však Linné přeřadil mrože do skupiny mořských krav Trichechus (kapustňák) a řádu Bruta, který podle Linného zahrnoval slony, lenochody a mravenečníky. V následujících desetiletích byli mroži i další ploutvonožci spojováni vedle zmíněných skupin živočichů hlavně se sirénami. Johann Friedrich Blumenbach mezi lety cca 1788–1825 řadil mrože a sirény do rodu Trichechus, který v jeho taxonomii společně s ptakopysky (Ornithorhynchus) utvářel řád Palmata.[3]
Prvním, kdo rozpoznal ploutvonožce jako samostatnou taxonomickou jednotku, byl německý přírodovědec Johann Karl Wilhelm Illiger, který v roce 1811 přiřadil jméno Pinnipedia nově vytvořenému řádu i čeledi těchto mořských savců.[4] Vědecké jméno Pinnipedia je odvozeno z latinského pinna (ploutev) a pes (noha), genitiv pedis.[5] Podobně i český název „ploutvonožci“ má základ ve slovech „ploutev“ a „noha“. Illigerův přínos spočíval zejména v tom, že rozdělil ploutvonožce a sirény do samostatných skupin (do té doby se obě tyto skupiny často řadily k sobě). Illigerovo zařazení ploutvonožců nicméně nebylo akceptováno všemi taxonomy a taxonomické debaty o zařazení ploutvonožců se vedou i ve 21. století.[3]
Americký zoolog Joel Asaph Allen v roce 1880 publikoval knihu Historie severoamerických ploutvonožců (celý název v originále zní History of North American pinnipeds, a monograph of the walruses, sea-lions, sea-bears and seals of North America), ve které provedl revizi dosavadního poznání těchto šelem. Allen rozebral biologické znaky čeledí a rodů, podrobně popsal severoamerické ploutvonožce a sestavil přehled všech ostatních známých ploutvonožců. Allen nahlížel na ploutvonožce jako na podřád skupiny Ferae.[3]
_(14568900397).jpg.webp)
Podobné přeřazování pokračovalo v průběhu konce 19. a 20. století. Skupina Pinnipedia byla klasifikována jako řád savců, podřád řádu šelem či jednoduše se název používal jako lidové označení jedné či více čeledí mořských šelem z arktických či antarktických oblastí.[6][7] Ve velké taxonomické revizi savců zpracované americkým paleontologem Georgem Gaylordem Simpsonem z roku 1945 se řád šelem dělil do tří skupin, a sice podřádů Creodonta, Fissipedia (suchozemské šelmy) a Pinnipedia (ploutvonožci).[8]
V roce 1989 tým vědců v čele s americkou paleontoložkou Annalisou Bertou navrhl vytvoření blíže nezařazeného kladu Pinnipedimorpha, který by zahrnoval i fosilní rod Enaliarctos.[9] Berta později přišla s tím, že sesterskou skupinou Enaliarctos je klad Pinnipediformes, který se dále dělí na Pteronarctos a Pinnipedia.[10]
Společnost pro mořskou mammalogii (The Society for Marine Mammalogy) hodnotí Pinnipedia k roku 2021 jako „nezařazenou taxonomickou jednotku“.[1] K tomu se přiklání i většina moderních fylogenetických studií z 21. století, které o ploutvonožcích píší jako o „skupině“ či „kladu“ bez přiřazení konkrétní taxonomické kategorie.[11][12] Prakticky všechny moderní fylogenetické studie potvrzují monofyletický původ skupiny Pinnipedia.[13][11][12][14]
Klad Pinnipedia je řazen do řádu šelem (Carnivora) a podřádu psotvární (Caniformia).[15] Řada autorů řadí Pinnipedia do infrařádu Arctoidea, kam se vedle ploutvonožců řadí ještě Ursoidea a Musteloidea.[14][12][13] Klad Pinnipedia (tzn. moderní ploutvonožci) zahrnuje tři čeledi: Otariidae (lachtanovité), Odobenidae (mrožovité) a Phocidae (tuleňovité). Lachtanovití a mrožovití tvoří nadčeleď Otarioidea[16] (která je nicméně patrně parafyletická[17]) a tuleňovití tvoří samostatnou nadčeleď Phocoidea.[18]
V roce 2012 zoologové Annalisa Berta a Morgan Churchill na základě morfologických studií a výsledků genetických výzkumů vytvořili fylogenetický strom ploutvonožců. Podle autorů existuje 33 recentních druhů ploutvonožců a 29 poddruhů (nicméně 5 z těchto poddruhů postrádá dostatečné podklady k tomu, aby mohly být jednoznačně identifikovány za poddruhy).[19] Berta a Churchill dále navrhli, aby rod Arctocephalus zahrnoval pouze lachtana jihoafrického a ostatní lachtani tohoto rodu se přesunuli do nově vytvořeného rodu Arctophoca.[19] I přes řadu rozsáhlých genetických studií se přesné fylogenetické vztahy ploutvonožců ve vědecké komunitě nadále diskutují.[20] Lze se setkat i s větším množstvím rozeznávaných druhů (např. Berta, Sumich a Kovacs v roce 2006 rozeznávali 34–36 druhů,[21] IUCN k roku 2021 rozeznává 36 druhů včetně dvou vyhynulých.[22]
K roku 2003 bylo popsáno více než 50 fosilních druhů ploutvonožců.[23]
Seznam druhů
Počet rozeznávaných druhů ploutvonožců se pohybuje kolem cca 33–36. Problematický je hlavně rod Arctocephalus a jeho rozdělení do druhů a poddruhů, což je předmětem neutuchajících vědeckých debat.[19][24] Níže je uveden seznam 35 druhů uváděných na webu Společnosti pro mořskou mammalogii:[1]
- čeleď Phocidae (tuleňovití)
- podčeleď Monachinae
- tribus Monachini
- rod Monachus
- tuleň středomořský (Monachus monachus)
- rod Neomonachus
- tuleň havajský (Neomonachus schauinslandi)
- tuleň karibský (Neomonachus tropicalis)
- tribus Miroungini
- rod Mirounga
- rypouš severní (Mirounga angustirostris)
- rypouš sloní (Mirounga leonina)
- tribus Lobodontini
- rod Ommatophoca
- tuleň Rossův (Ommatophoca rossi)
- rod Lobodon
- tuleň krabožravý (Lobodon carcinophagus)
- rod Hydrurga
- tuleň leopardí (Hydrurga leptonyx)
- rod Leptonychotes
- tuleň Weddellův (Leptonychotes weddellii)
- počeleď Phocinae
- tribus Cystophorini
- rod Cystophora
- čepcol hřebenatý (Cystophora cristata)
- tribus Erignathini
- rod Erignathus
- tuleň vousatý (Erignathus barbatus)
- tribus Phocini
- rod Phoca
- tuleň obecný (Phoca vitulina)
- tuleň pacifický (Phoca largha)
- rod Pusa
- tuleň kroužkovaný (Pusa hispida)
- tuleň bajkalský (Pusa sibirica)
- tuleň kaspický (Pusa caspica)
- rod Pagophilus
- tuleň grónský (Pagophilus groenlandicus)
- rod Histriophoca
- tuleň pruhovaný (Histriophoca fasciata)
- rod Halichoerus
- tuleň kuželozubý (Halichoerus grypus)
- čeleď Otariidae (lachtanovití)
- rod Arctocephalus
- lachtan jihoamerický (Arctocephalus australis)
- lachtan Forsterův (Arctocephalus forsteri)
- lachtan galapážský (Arctocephalus galapagoensis)
- lachtan antarktický (Arctocephalus gazella)
- lachtan ostrovní (Arctocephalus philippii)
- lachtan jihoafrický (Arctocephalus pusillus)
- lachtan jižní (Arctocephalus tropicalis)
- rod Callorhinus
- lachtan medvědí (Callorhinus ursinus)
- rod Eumetopias
- lachtan ušatý (Eumetopias jubatus)
- rod Neophoca
- lachtan šedý (Neophoca cinerea)
- rod Otaria
- lachtan hřivnatý (Otaria flavescens)
- rod Phocarctos
- lachtan novozélandský (Phocarctos hookeri)
- rod Zalophus
- lachtan tmavý (Zalophus californianus)
- lachtan japonský (Zalophus japonicus)
- lachtan mořský (Zalophus wollebaeki)
- čeleď Odobenidae (mrožovití)
- rod Odobenus
- mrož lední (Odobenus rosmarus)
Charakteristika recentních čeledí
Lachtanovití (Otariidae) jsou typičtí přítomností ušních boltců. Ve vodě se pohybují pomocí dobře vyvinutých předních ploutví. Zadní ploutve jsou ohebné dopředu a lachtani je při pohybu po souši podsouvají pod sebou a mají tak funkci jakýchsi „zadních noh“.[25] Tradičně se lachtanovití dělí do dvou podčeledí, a sice:
- Otariinae (v angličtině označováni jako sea lions, doslova „mořští lvi“) – pro lachtany z této podčeledi jsou typické zakulacené čenichy a krátké tlusté chlupy.[26]
- Arctocephalinae (v angličtině fur seals, ve volném překladu „osrstění lachtani“) – zástupci podčeledi Arctocephalinae bývají o trochu menší než Otariinae, mají zašpičatělé čenichy, o něco delší přední ploutve a mnohem hustší srst, která zahrnuje i huňatou podsadu s pesíky.[26]
Rozdělení lachtanovitých do těchto dvou podčeledí však nabouraly rozbory DNA. Genetická studie z roku 2001 nejdříve zaznamenala, že lachtan medvědí z podčeledi Arctocephalinae je příbuzný spíše některým zástupcům z podčeledi Otariinae než ostatním zástupcům Arctocephalinae.[27] Dále podle výsledků studie DNA z roku 2006 jsou lachtan šedý a lachtan novozélandský (zástupci Otariinae) nejblíže příbuzní lachtanům z rodu Arctocephalus (podčeleď Arctocephalinae).[28]
Mrožovití (Odobenidae) mají pouze jediného současného zástupce, kterým je mrož lední. Mrož je dobře rozeznatelný od ostatních současných ploutvonožců mohutností svého těla (většími ploutvonožci jsou jedině rypouši), takřka neosrstěnou kůží a především prodlouženými horními špičáky – kly. Stejně jako lachtanovití, i mroži dokáží otočit své zadní ploutve dopředu pro snadnější pohyb po souši. Ve vodě se při nízkých rychlostech pohybují pomocí ploutví, vyšších rychlostí dosahují vlněním páteře.[11] Stejně jako tuleni, i mroži postrádají ušní boltce.[29] Pyjová kost mrožů dosahuje délky až 63 cm, což představuje vůbec nejdelší pyjovou kost ze všech savců.[30]
Tuleňovití (Phocidae) postrádají ušní boltce a jejich ploutve směřují trvale dozadu, takže je nemohou při pohybu na souši podsouvat pod tělem. Při pohybu po souši tak působí poněkud těžkopádně a suchozemského pohybu dosahují pomocí pomalého plazení v kombinaci s drobnými přískoky. Ve vodě jsou nicméně rychlejší a obratnější než ostatní ploutvonožci; k pohybu používají střídavých pohybů zadní části těla s menším přispěním zadních ploutví, které používají hlavně pro kormidlování. Z důvodů využívání pohybů těla pro plavání je nejvíce svaloviny soustředěno kolem páteře.[25][30] Podle studie molekulární DNA z roku 2006 se dají tuleňovití rozdělit do dvou monofyletických podčeledí, a sice Monachinae (zahrnující triby Monachini, Miroungini a Lobodontini) a Phocinae (zahrnující rody Pusa, Phoca, Halichoerus, Histriophoca, Pagophilus, Erignathus a Cystophora).[28]
Evoluce

Některé starší zdroje uvádí, že ploutvonožci jsou difyletického původu, čili že pocházejí ze dvou společných předků. Původ lachtanovitých a mrožovitých býval dáván do souvislosti s medvědovitými a původ tuleňovitých býval spojován s lasicovitými.[31] Nicméně moderní morfologické a molekulární rozbory ukazují jednoznačně na monofyletický původ.[31][32][33] Otázka, zda jsou ploutvonožci příbuzní spíše medvědovitým, nebo lasicovitým, však nebyla uspokojivě zodpovězena. Zatímco některé zdroje uvádí obě skupiny,[33] řada studií se přiklání k medvědovitým[34][35][36] a jiné zase k lasicovitým.[28][37][38]
Ploutvonožci se oddělili od ostatních psotvárných asi před 50 miliony lety v době eocénu.[39] Evoluční link mezi ploutvonožci a suchozemskými savci patrně představuje Puijila darwini. Fosilie tohoto raně miocenního savce byla objevena v roce 2007 v Nunavutu, arktické oblasti Kanady, kde žil někdy před 24–20 miliony lety. Podobně jako recentní vydry, Puijila darwini měla dlouhý ocas a krátké končetiny s plovacími blánami, které naznačují počínající adaptaci na vodní prostředí, avšak stále se nejedná o ploutve jako u moderních ploutvonožců. Předpokládá se, že zvíře se pohybovalo po souši po čtyřech končetinách a pro pohyb ve vodě využívalo taktéž jen končetiny. Puijila darwini měla velké oči, což je typické i pro moderní ploutvonožce. Takřka celá kostra druhu byla nalezena v jezerním nánosu, takže se dá předpokládat, že ploutvonožci během evoluce prošli sladkovodní fází, tzn. jejich prapředci se nejdříve adaptovali z oceánu na souš a poté se přizpůsobili nejdříve na sladké a nakonec zpětně i na slané vodní prostředí. Vědci, kteří tento chybějící článek objevili, druh zařadili do stejného kladu jako Potamotherium a Enaliarctos. Ve srovnání s těmito taxony byla Puijila darwini nejméně uzpůsobena životu ve vodě.[40][41]
.JPG.webp)
Enaliarctos představuje pozdně oligocenní / raně miocenní rod šelem, jehož zástupci žili v dnešní Kalifornii před 24–22 miliony lety. Zástupci rodu Enaliarctos silně připomínají moderní ploutvonožce. Jejich těla byla dobře adaptována pro pohyb ve vodě díky ploutvím a pružné páteři. Jejich zuby byly přizpůsobeny trhání kořisti, což je typické pro suchozemské šelmy. Tyto dávné šelmy se patrně zdržovaly poblíž břehů častěji než recentní ploutvonožci. Enaliarctos používali k plavání předních i zadních končetin, avšak přední končetiny patrně používali více.[31] U jednoho z druhů tohoto rodu, Enaliarctos emlongi, byl zjištěn výrazný pohlavní dimorfismus, což napovídá, že fyzické znaky hrály v evoluci ploutvonožců důležitou roli. Druh byl patrně vysoce polygynní, což je u běžné i u moderních ploutvonožců.[42]
Příbuznějšího předka moderních ploutvonožců představuje rod Pteronarctos. Zástupci tohoto fosilního taxonu žili v americkém Oregonu někdy před 19–15 miliony lety. Podobně jako u moderních ploutvonožců, očnice šelem z rodu Pteronarctos nebyly tvarovány obličejovými kostmi jako je slzní nebo lícní kost, ale především horní čelistí.[31]

K dalším fosilním taxonům patří např. vyhynulá čeleď Desmatophocidae, jejíž zástupci žili v severním Pacifiku někdy před 23–10 miliony lety. Desmatophocidae měli protáhlé lebky, velké oči a zaoblené lícní zuby. Disponovali výrazným pohlavním dimorfismem a k pohybu ve vodě patrně využívali předních i zadních ploutví.[31]
Předkové nadčeledí Otarioidea a Phocoidea se od sebe oddělili před cca 33 miliony lety.[39] Phocidae se vydělili nejméně před 15 miliony lety (patrně ještě dříve)[31] a podle molekulárních analýz se podčeledi Monachinae a Phocinae začaly oddělovat někdy před 22 miliony let.[28] První zástupci Phocidae se přitom vyvinuli v severním Atlantiku, kam jejich předek doputoval patrně buď severní cestou přes Arktidu nebo jižní cestou přes oblast dnešní Panamské šíje.[31]
Fosilní pozůstatky rodů Monotherium (podčeleď Monachinae) a Leptophoca (podčeleď Phocinae) ještě byly nalezeny na společném území, a sice v jihovýchodní části Severní Ameriky. Někdy před 17 miliony lety došlo k vydělení rodů Erignathus a Cystophora.[31] Co se týče Phocinae, tak rody Phoca a Pusa vznikly patrně poté, co se Phocinae rozšířili až do pradávného moře Paratethys a odtud začali směřovat k arktické pánvi a dále na východ. Z oblasti Arktidy tuleňovití ploutvonožci doputovali až k jezeru Bajkal, kde se v izolaci vyvinuli do dnešních tuleňů bajkalských. V izolaci se ocitli i prapředci tuleňů kaspických, když ústup moře Paratethys dal vzniknout bezodtokovému Kaspickému moři.[31] Co se Monachinae týká, tak ve Středozemním moři se nejdříve vyvinul rod Monachus, který se rozšířil do Karibiku a poté do středních částí severního Tichomoří.[43] Dva současní zástupci rodu Mirounga (rypouš) se patrně oddělili do samostatných druhů před cca 4 miliony lety. Rozdělení rypoušů do dvou linií je dáváno do souvislosti se vznikem Panamské šíje, která rozdělila tehdejší rypouše na dvě populace;[28] je však možné že rypouši nejdříve osídlili antarktické oblasti a teprve odtud se rozšířili na sever.[44] Předci tribu Lobodontini se objevili někdy před 9 miliony lety, kdy kolonizovali oceánské oblasti kolem Antarktidy.[43]

Lachtanovití (Otariidae) a mrožovití (Odobenidae) se rozdělili někdy před 28 miliony lety.[28] Prapůvodní domovinou lachtanů je severní Tichomoří. Nejstarší datovaný fosilní nález lachtana představují pozůstatky jedince z rodu Pithanotaria, který žil v Kalifornii někdy před cca 11 miliony lety. Rod Callorhinus se vydělil před 16 miliony lety. Následovalo vydělení rodů Zalophus, Eumetopias a také rodu Otaria, který kolonizoval vody kolem Jižní Ameriky. Většina ostatních druhů lachtanů se vyvinula na jižní polokouli. Nejraněji datované fosilie mrožovitých představuje rod Prototaria z Japonska (stáří 16–14 milionů let[45]) a Proneotherium z Oregonu (před cca 20–16 milionů let).[46] Tito předci dnešního mrože měli mnohem kratší špičáky a živili se rybami, zatímco dnešní mroži se živí převážně měkkýši. Další druhy mrožů se vyvinuly ve středním a pozdním miocénu. Několik druhů mělo zvětšené horní i dolní špičáky. U rodů Valenictus a Odobenus již byly přítomny dlouhé špičáky, resp. kly.[31] Patrně před vznikem Panamské šíje, někdy před 8–5 miliony lety, se předci moderních mrožů rozšířili ze severního Tichomoří do Karibiku a dále do severního Atlantiku a poté přes Arktidu se před 1 milionem let vrátili do severního Tichomoří. Podle alternativní teorie se předci mrožů rozšířili ze severního Tichomoří do Arktidy a poté v období pleistocénu i do severního Atlantiku.[31]
Níže se nachází kladogram zobrazující hlavní vyhynulé i současné linie a rody kladu Pinnipedia. Kladogram byl v mírně upravené podobě převzat od Annalisy Berty (2009).[31]
| ← Pinnipedia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Anatomie a fyziologie
Popis


Ploutvonožci mají protáhlá hydrodynamická těla se zmenšenými (lachtanovití) či chybějícími ušními boltci (tuleňovití, mrož), zakulacenou zploštělou hlavou, flexibilním krkem a krátkým ocasem. Končetiny jsou přeměněny v ploutve.[47][48] Lebky ploutvonožců mají velké očnice, krátké čenichy a zúženou interorbitální oblast (plocha mezi očima).[49] Tito mořští savci jsou v rámci šelem jedineční v tom, že stěny očnic jsou výrazně tvarovány horní čelistí a ne kostmi obličejové části.[31] Zašpičatělé zuby jsou kuželovitého tvaru. Oproti většině ostatních šelem mají méně zubů (zvláště řezáků a zadních stoliček). Trháky úplně chybí.[50] U mrože se horní špičáky vyvinuly v dlouhé kly, což je u ploutvonožců jedinečné.[51] Prsní žlázy a genitálie se mohou zatáhnout dovnitř těla.[47]
Velikost ploutvonožců se různí. Obecně mají větší těla než ostatní šelmy.[47] Nejmenšího ploutvonožce představuje tuleň bajkalský, jehož délka se pohybuje kolem 1,1–1,4 m a váha mezi 50–130 kg.[52] Naopak největším ploutvonožcem je rypouš sloní, který může měřit až kolem 6 m a vážit přes 4 tuny.[53] Tyto rozměry dělají z rypouše sloního vůbec největšího zástupce řádu šelem.[47] Několik druhů disponuje výrazným pohlavním dimorfismem, který se projevuje hlavně odlišnou velikostí těla, která je pozitivně spjatá s mírou polygynie. Obecně se dá říci, že extrémně polygynní druhy jako rypouši jsou extrémně dimorfní (např. samec rypouše sloního může vážit i 10× více než samice[54]), zatímco samci méně polygynních druhů mají vzhledem k samicím menší velikostní poměr.[55][56] I přes tento přímo úměrný vztah mezi pohlavním dimorfismem a mírou polygynie se podle některých výzkumů rozdílná velikost mezi pohlavími vyvinula následkem odlišností v ekologii a navíc ještě před vývinem polygynie.[57][58] Samice z tribu Lobodontini jsou mírně větší než samci. Samci pohlavně dimorfních druhů často mívají sekundární pohlavní znak jako je např. chobot u rypoušů, nafukovací vak u čepcolů hřebenatých či tlusté krky a hřívy u lachtanů.[55][56]

Takřka všichni ploutvonožci mají těla pokrytá srstí. Výjimku představuje mrož, jehož kůži pokrývá pouze řídké ochlupení. I plně osrstěné druhy jako lachtani podčeledi Otariinae mají méně chlupů než většina suchozemských savců.[59] Mláďata druhů, které žijí na ledu, mívají hustší srst než dospělci. Jednotlivé chlupy na srsti mláďat (tato srst se nazývá lanugo), mají schopnost zachycovat teplo ze slunečních paprsků a udržet tak mláďata v teple.[60] Zbarvení srsti ploutvonožců využívá principu protistínu, maskovací techniky založené na postupném zesvětlování barevných odstínů srsti od tmavých na dorzální (hřbetní) straně těla ke světlejším na ventrální (břišní) straně těla. Takto se potlačí stín dopadající na spodní část těla při průsvitu paprsků skrz vodní sloupec a dojde k oslabení trojrozměrného vnímání ploutvonožců ostatními živočichy, takže jejich těla lépe splynou s okolním prostředím.[61][62] Mláďata tuleňů grónských jsou čistě bílá, což je dobře skryje v arktickém prostředí.[61] Některé druhy jako tuleni pruhovaní, kroužkovaní či leopardí mají srst posetou kontrastními vzory světlé a tmavé barvy. Všichni plně osrstění ploutvonožci línají; tuleňovití jednou za rok během několikatýdenního období, u lachtanovitých dochází k nahrazování srsti průběžně po celý rok.[63] Všichni ploutvonožci disponují vrstvou podkožního tuku, která je zvláště silná u tuleňů a mrožů.[47] Podkožní tuk má dvojí funkci: poskytuje potřebnou termoizolaci a slouží jako zásobárna energie v postních obdobích, která nastávají během páření, u některých samic během laktace a u tuleňovitých i během línání.[60] Termoizolace u lachtanů z podčeledi Arctocephalinae je zajišťována primárně jejich hustou srstí s huňatou podsadou.[64] Podkožní tuk může představovat až 50 % váhy zvířete. Mláďata se rodí pouze s tenkou vrstvou podkožního tuku, což je kompenzováno hustým lanugem.[60]
Ploutvonožci mají jednoduchý žaludek, který je stavbou podobný žaludkům suchozemských šelem. Většina druhů nemá slepé střevo. Přechod mezi tenkým a tlustým střevem je nejasný; tlusté střevo je vzhledem k tenkému velmi krátké a jen o trochu širší. Délky tenkých střev se pohybují od osminásobku délky těla (lachtan kalifornský) až po 25 násobek tělesné délky (rypouši). Délka střev je patrně důsledek adaptace na časté hloubkové potápění. Zvýšený objem zažívacího traktu totiž během ponoru slouží jako úložný prostor pro částečně natrávenou kořist.[65] Podobně jako u většiny ostatních mořských savců, ledviny jsou rozčleněny na menší laloky, které dovedou účinně absorbovat vodu a odfiltrovat z ní přebytečnou sůl.[66]
Pohyb


Ploutvonožci disponují předními i zadními ploutvemi. Lokty a kotníky nevyčnívají ven, ale jsou umístěny uvnitř těla.[67] Na rozdíl od suchozemských šelem, které někdy mají jen čtyři prsty, všichni ploutvonožci mají pět prstů. Ty jsou spojené plovací blánou.[68] Jsou pomalejšími plavci než kytovci, ve vodě dokáží vyvinout maximální rychlost 9–28 km/h v závislosti na druhu (srv. například s delfíny, kteří plavou rychlostí až cca 37 km/h). Ploutvonožci jsou ve vodě velice mrštní a hbití.[59] Někteří z lachtanů jako je lachtan kalifornský dokáží ohnout krk dozadu a hlavou se dotknout zadních ploutví.[69] U ploutvonožců se vyvinulo několik adaptací pro zmenšení odporu, kterému čelí při pohybu vodním sloupcem. Vedle hydrodynamicky tvarovaných těl to je jemná síť svalových svazků přítomných v kůži. Tato miniaturní svalová vlákna dovedou navýšit laminární proudění krve, díky čemuž mohou proklouzávat vodou snadněji. U ploutvonožců chybí hladká svalovina (musculus arrector pili) napojená na vlasové folikuly. Tato svalovina způsobuje napřímení chlupů (známé např. během „husí kůže“), takže chlupy ploutvonožců se dokáží snadno přizpůsobit pohybu plavání tak, aby kladly co nejmenší odpor.[59]
Lachtanovití při pohybu ve vodě spoléhají na své přední ploutve, které používají k plavání a potápění podobným způsobem jako mořské želvy nebo tučňáci.[70] Oproti suchozemským šelmám došlo u lachtanovitých k redukci délky předních končetin, což poskytuje mechanickou výhodu pohybovým svalům v oblasti ramen a loktů (menší páka).[67] Zadní ploutve lachtanovitých slouží především k vyvažování.[59] Naproti tomu tuleni a mroži používají pro plavání svých zadních ploutví a vlnění zadní části těla ze strany na stranu.[70] Jejich přední ploutve slouží k určování směru pohybu.[67] Některé druhy při plavání vyskakují nad vodní hladinu, což jim umožňuje pohybovat se rychleji. Lachtani z podčeledi Otariinae někdy „chytají vlny“ (tzn. pohybují se nebo skáčou ve směru pohybujících se vln), díky čemuž patrně dokáží snížit svůj energetický výdej potřebný k překonání vzdáleností.[59]
Ploutvonožci se dokáží pohybovat i na souši, avšak ne tak obratně jako suchozemští savci. Lachtanovití a mroži dokáží přetočit své zadní ploutve dopředu, takže je při pohybu po souši podsouvají pod tělem a dokáží se tedy pohybovat po všech čtyřech končetinách.[71] Předními ploutvemi přitom pohybují spíše příčně než sagitálně. Tuleňovití nedokáží otočit své zadní ploutve dopředu a při pohybu po souši se spoléhají hlavně na své hlavy a krky.[72] Pohybu dosahují rozkýváním vrchní části těla, vrtěním i malými přískoky. Přední ploutve jim slouží pouze k udržování rovnováhy, i když některé druhy tuleňů dokáží využít přední ploutve i ke krátkému popotažení. Tuleni jsou tedy na souši poměrně neobratní, i když na ledu jim to jde o něco lépe, jelikož dovedou využít jeho klouzavých vlastností. Naproti tomu lachtani dokáží i např. vylézt po schodech či „dojít“ po souši poměrně daleko od moře.[71]
Smysly
Oči ploutvonožců jsou vzhledem k velikosti těl poměrně velké. Jsou umístěny v přední části hlavy; výjimku představuje pouze mrož, u něhož jsou oči po stranách hlavy.[73][74] Důvod je patrně ten, že mrož se krmí na málo pohyblivých měkkýších ze dna moře, k čemuž nepotřebuje tak ostré vidění zajištěné oběma očima v přední část hlavy jako to mají ostatní ploutvonožci, kteří se živí mnohem pohyblivější kořistí.[73] Oči ploutvonožců prošly dvojí adaptací, a sice z vodního prostředí na souš a poté zpětně na vodní prostředí, což přitahovalo pozornost vědců již od 19. století.[75] Čočka je zakulacená, většina sítnice se nachází ve stejně vzdálenosti od středu čočky. Rohovka má plochou středovou část, takže refrakce je takřka stejná ve vodě i na souši. Duhovkové svaly i cévy jsou zesílené. Dobře vyvinutý rozvěrač zornice (musculus dilatator pupillae; sval pohánějící rozšíření zornice) zajišťuje dostatečně velký rozsah rozšíření zornice, což je potřeba ve zhoršených světelných podmínkách jako jsou hluboké vody oceánů, kam proniká jen málo světla. Zúžená zornice má většinou hruškovitý tvar. Schopnost míry otevření zornice je menší u druhů, které žijí v mělkých vodách (např. lachtan kalifornský, tuleň obecný), zatímco druhy lovící v hlubokých oceánských vodách mohou své zornice rozevřít mnohem více.[76]
Na souši ve slabém světle jsou ploutvonožci krátkozrací. Krátkozrakost se v jasném světle zmírňuje, jak zúžená zornice snižuje schopnost čočky a rohovky ohýbat světlo. Ploutvonožci mají dobře vyvinuté tapetum lucidum, odrazivou vrstvu mezi sítnicí a cévnatkou, která zesiluje dopadající světlo jeho odrazem zpět do tyčinek. To umožňuje lepší vidění v horších světelných podmínkách.[73] Rohovky druhů žijících na ledu (např. tuleň grónský) mají vysokou toleranci ultrafialového záření, které je typické pro jasné, sněhem pokryté prostředí, takže u u těchto druhů nedochází k vývinu sněžné slepoty.[77] Ploutvonožci patrně mají omezené barevné vidění, protože nemají čípky typu S (jeden ze tří druhů čípků, které zajišťují barevné vidění).[78] Mroži mají dobře vyvinuté okohybné svaly, což spolu s chybějící horní stěnou očnice umožňuje povystrčení očí a vizi jak dopředu tak dozadu.[79] Ploutvonožci na ochranu svých očí vylučují velké množství hlenu.[59] Rohovkový epitel je keratinizovaný a oční bělmo je dostatečně silné na to, aby odolalo tlakům během potápění. Podobně jako u řady jiných savců a ptáků, i u ploutvonožců je přítomná mžurka (membrana nictitans).[80]
Uši jsou uzpůsobeny pro poslouchání pod vodou, kde ploutvonožci dokáží zaznamenat frekvence až do 70 000 Hz. Na souši je sluch ploutvonožců oproti mnohým suchozemským savcům redukován. I když jsou schopni slyšet široké spektrum frekvencí (např. u lachtana medvědího to je 500–32 000 Hz, srovnej 20–20 000 Hz u lidí), jejich citlivost na vnímání vzdušných zvuků je jen nízká.[81] Studie z roku 1998 zkoumala sluch u tří ploutvonožců, a sice tuleně obecného, lachtana kalifornského a rypouše severního. Podle závěrů studie byl nejlépe adaptován na vnímání vzdušných zvuků lachtan kalifornský, tuleň obecný slyšel stejně dobře ve vodě i na vzduchu a sluch rypouše severního byl lépe adaptován na vnímání zvuků pod vodou.[82] Ačkoli ploutvonožci mají na souši celkem dobrý čich,[83] pod vodou ho kvůli zavřeným nozdrám nepoužívají.[84]
Hmatové smysly jsou dobře vyvinuty. Ploutvonožci disponují vibrisy (hmatové vousky známé též např. u koček), díky kterým dokáží účinně zaznamenávat i sebemenší vibrace ve vodě. Na každý hmatový vous je napojeno množství mechanoreceptorů a 1000–1600 nervových vláken, zhruba 10× více než u suchozemských savců.[85][86] Vibrace ve vodě vznikají např. při plavání ryby skrz vodu. Detekce těchto vibrací je klíčová při hledání potravy a zvláště v hlubokých vodách s nedostatkem světla doplňuje či dokonce nahrazuje zrak.[87] Díky vibracím dokáží ploutvonožci následovat kořist či ostatní jedince svého druhu i několik minut po jejich proplutí, a to na základě hydrodynamických stop šířených vodou. Ploutvonožci dokáží pomocí hmatových vousků dobře odhadnout velikost pronásledované kořisti a dokonce i konkrétní druh.[88][89][86] Dovedou rozeznat i pouze několikamilimetrové rozdíly v texturách a tvarech objektů.[86] U tuleňů kroužkovaných na finském jezeře Saimaa bylo prokázáno úspěšné lovení kořisti i přesto, že byli slepí. Lze předpokládat, že se při lovu spoléhali právě na své vibrisy.[90]
U ploutvonožců se rozlišují dva typy vibrisů, hladké a vlnkované (vlnky jsou okem sotva postřehnutelné; jedná se o mírnou změnu tloušťky hmatového chlupu každých několik milimetrů jeho délky). Mroži, lachtanovití a dva druhy tuleňů (tuleň středomořský, tuleň vousatý) mají hladké vibrisy, všichni ostatní tuleňovití je mají vlnkované.[91] Ploutvonožci představují jediné živočichy s vlnkovanými vibrisy.[92] Funkce těchto zvláštních vibrisů zůstává nejasná, i když vědci předložili několik teorií jako např. že vlnkovaná struktura zvyšuje citlivost vnímání vibrací, redukuje šum či snižuje dopady víření, které nastává při pohybu ploutvonožce skrze vodní sloupec.[91] Z dosavadních výzkumů se však zdá, že na citlivost vnímání má spíše vliv úhel, jaký vibrisy svírají k průtoku vody, než tvar vibrisů (přinejmenším lachtani kalifornští a tuleni obecní při pronásledování kořist mají napřímené vibrisy, takže svírají k tělu pravý úhel).[91] U lachtanovitých mohou být hmatové vousky poměrně dlouhé a např. u lachtana antarktického dosahují až 41 cm.[93] Nejvíce vibrisů mají mroži, kteří disponují 600–700 hmatovými chlupy. Vibrisy patrně hrají roli i v navigaci. Některým druhů jako je tuleň kroužkovaný patrně pomáhají během plavání pod ledem v lokaci dýchacích děr.[94]
.jpg.webp)


.jpg.webp)

Potápění

Ploutvonožci jsou zdatní potápěči. Před potopením typicky vyprázdní kolem poloviny vzduchu ze svých plic[95] a uzavřou nozdry a hrtanové chrupavky, aby tak ochránili průdušnice.[96] Dýchací cesty vedoucí do plic jsou zesíleny prstencovými chrupavkami a hladkou svalovinou. Plicní sklípky se při hlubších ponorech zcela vyprázdní.[97][98] Na rozdíl od většiny suchozemských savců jsou ploutvonožci schopni úplně vyprázdnit své plíce.[99][100] Ve středním uchu se nachází dutiny, které se během ponorů patrně naplní krví, čímž se zabraňuje ušnímu barotraumatu (fyzickému poškození tkání způsobené rozdílem tlaků).[101] Průdušnice je pružná, což umožňuje její stlačení při velkých tlacích.[95] Během hlubokých ponorů je všechen zbývající vzduch uložen v průdušinkách a průdušnici, což zabraňuje vývinu dekompresní nemoci, opojení dusíkem i toxicity kyslíku. Ploutvonožci mají vysokou toleranci kyseliny mléčné, což snižuje únavu kosterního svalstva při intenzivních fyzických aktivitách jako je potápění.[101]
Na potápění prošla adaptací i oběhová soustava. Během ponorů se ploutvonožcům rozšíří žíly, čímž se navýší jejich kapacita a mohou tak přenášet větší množství kyslíku. V jejich svalech a krvi je uloženo velké množství hemoglobinu a myoglobinu. Relativní množství krve v tělech ploutvonožců je vyšší než u většiny ostatních savců (cca 2,5× vyšší než u člověka), což opět pomáhá zvyšovat zásoby kyslíku. U hloubkových potápěčů, jako jsou rypouši, představuje krev až 20 % hmotnosti těla. Krevní barvivo zvířat dokáže dobře vázat kyslík. Ploutvonožci mají vysokou toleranci oxidu uhličitého v krvi a dokáží snížit spotřebu kyslíku po ponoření pomocí zúžení periferních cév.[68][102] Ploutvonožci pod hladinou také výrazně omezují tepovou frekvenci (bradykardie) a jejich těla koncentrují kyslík v klíčových životních orgánech, jako je srdce, centrální nervová soustava a plíce.[103][101]
Termoregulace

U ploutvonožců hraje důležitou roli schopnost udržovat vysokou tělesnou teplotu v chladném prostředí. K tomu jim napomáhá nízký poměr povrchu těla k jeho objemu, silná vrstva podkožního tuku,[104] srst a rychlý metabolismus.[105] Cévy v ploutvích jsou adaptovány na protiproudou výměnu. Žíly přenášející chladnou krev z končetin jsou umístěny v blízkosti tepen, které přinášejí teplou krev z jádra těla. Teplem z této krve se otepluje studenější krev v cévách, která pak putuje zpět směrem k srdci.[106]
Na souši, zejména během teplých slunečních dní, ploutvonožcům naopak hrozí přehřátí těl. Některé druhy se zbavují přebytečného tepla pomocí nahazování studeného, vlhkého písku na záda, aby se tak navýšila tepelná výměna. Rypouši severní pomáhají ochlazovat svá těla supěním, tuleni z tribu Monachini se na plážích snaží prohrabat k vrstvě studeného písku, na kterém pak odpočívají. Ploutvonožci někdy odpočívají v bazénkách, pokud jsou dostupné.[107] Přinejmenším u některých druhů ploutvonožců (rypouš sloní,[108] tuleň obecný, kuželozubý a grónský) dochází k odvodu tepla pomocí termálních oken umístěných na trupu a ploutvích.[107][109]
Spánek
Tyto mořské šelmy tráví na otevřeném moři i měsíce v kuse, takže ke spánku nutně dochází na otevřené vodní ploše. Ploutvonožci většinou spí „na půl mozku“, kdy jedna mozková hemisféra spí a druhá zůstává vzhůru. Takoví lachtani se položí na vodě na stranu s břichem nahoru. Jednu z předních ploutví ponoří do vody a jemně s ní pádlují, což je udržuje na hladině. Další tři ploutve vystrčí nad hladinu, patrně za účelem minimalizace ztráty tepla. Nad hladinou je vystrčený i čumák, který lachtanům umožňuje dýchat.[110] Pokud mají lachtani na výběr, spí raději na souši,[111] kde se ke spánku uchylují obě hemisféry.[112]
Někteří jiní ploutvonožci jako je rypouš severní spí pod hladinou. Položí se na záda a s břichem obráceným vzhůru pomalými krouživými pohyby připomínajícím padání listu klesají ke dnu. Rypouši takto pomalu padají i desítky minut až do hloubek kolem 400 m, kde jsou dobře chráněni před predátory. Poté se probudí, vyplavou na hladinu nabrat vzduch a proces se opakuje.[113]
Rozšíření a habitat

Ploutvonožci většinou obývají polární a subpolární oblasti, jako jsou severní a jižní Tichomoří, severní a jižní Atlantik, Severní ledový oceán a Jižní oceán. Tuleň kaspický a bajkalský obývají velké vodní plochy hluboko ve vnitrozemí (Kaspické moře a jezero Bajkal). V orientální oblasti se ploutvonožci nevyskytují vůbec.[114] Tuleni tribu Monachini (tuleni středomořští, havajští a karibští) a někteří z lachtanovitých žijí v subtropických a tropických oblastech. Ploutvonožci typicky vyžadují chladnější vody bohaté na živiny, s teplotou do 20 °C. Výjimku představují pouze tuleni tribu Monachini, kteří se vyskytují i v teplejších vodách.[114][115]
Ploutvonožci obývají širokou škálu vodních habitatů: pobřežní vody, volná moře, brakické vody či dokonce sladkovodní jezera a řeky. Většina druhů vyhledává pobřežní oblasti, i když řada z těchto druhů pak cestuje za potravou do hlubších vod daleko od břehu. Tuleň bajkalský je jediný čistě sladkovodní ploutvonožec. Sladké vody příležitostně obývá i tuleň kroužkovaný, který se občas vyskytuje ve sladkovodních jezerech v Rusku poblíž Baltského moře. Tuleni obecní občas navštěvují estuária, jezera či řeky, kde mohou pobývat až rok. I další druhy, jako lachtan kalifornský či lachtan hřivnatý, příležitostně navštěvují sladkovodní plochy.[116] Ploutvonožci se po část roku vyskytují i na souši, kde obývají širokou škálu ostrovních i kontinentálních habitatů. V mírných a tropických oblastech se vyskytují na písčitých i oblázkových plážích, skalnatých březích, mělčinách, wattech, v přílivových bazéncích a v mořských jeskyních. Některé druhy občas odpočívají na lidmi vytvořených objektech, jako jsou mola, bóje a ropné plošiny. Občas se ploutvonožci vydají i do vnitrozemí, kde odpočívají na písečných dunách a vegetaci. Příležitostně vyšplhají i na útesy.[117] Druhy žijící v polárních oblastech se vyskytují na kontinentálním ledu i ledových krách.[118]
Biologie a ekologie
Život na vodě
%252C_Point_Lobos%252C_CA%252C_US_-_Diliff.jpg.webp)
Většinu času tráví ve vodě, avšak přicházejí na břeh za účelem páření, vrhů, výchovy mláďat, línání, odpočinku, termoregulace či v případě ohrožení vodními predátory. Některé druhy migrují obrovské vzdálenosti v honbě za potravou nebo při změnách životního prostředí, které mohou nastat např. následkem vlivů El Niña nebo změn v ledové pokrývce. Rypouši zůstávají na moři po 8–10 měsíců; na souš dochází typicky pouze za účelem páření a línání, přičemž v mezičase se vydávají do lovišť vzdálených i tisíce kilometrů.[119] Rypouši severní mají vůbec nejdelší migrační trasy ze všech savců.[120] U samic rypoušů severních bylo zaznamenáno, že během jediného roku urazí cca 18 tisíc kilometrů a samci i 21 tisíc kilometrů.[120] Obecně platí, že tuleňovití migrují více než lachtanovití.[119] Pro navigaci ploutvonožci využívají geomagnetická pole, vodní i větrné proudy, polohu Slunce a Měsíce nebo chuť a teplotu vody.[121]
U ploutvonožců je běžné potápění, a to buď za účelem lovu nebo při snaze uniknout predátorům. Např. tuleň Weddellův se většinou potápí na méně než 15 minut a do hloubek kolem 400 m, ale dokáže se potopit i na mnohem déle (73 minut) a hlouběji (600 m).[122] Nejdéle a nejhlouběji ze všech ploutvonožců se potápí rypouši. Rypouš severní se běžně potápí do hloubek kolem 350–650 m na dobu kolem 20 minut, avšak dokáží se potopit až do hloubek kolem 1500 m na 77 minut.[123] Rypouš sloní byl dokonce zaznamenán při ponoru do hloubky 2 388 m[124] a pod vodou dokáže strávit až 120 minut.[125] Ponory lachtanů bývají kratší a méně hluboké. Typicky trvají kolem 5–7 minut s průměrnou hloubkou ponoru 30–45 m. Rekordmanem mezi lachtanovitými je lachtan novozélandský, který byl zaznamenán při ponoru do hloubky 460 m a pod vodou zůstal 12 minut.[122] Mroži se většinou nepotápějí příliš hluboko, jelikož potravu vyhledávají především v mělkých vodách.[126]
Doba dožití se obecně pohybuje kolem 25–30 let, i když se může výrazně lišit podle druhu. Samice většinou žijí déle.[127] K nejstarším zaznamenaným jedincům ve volné přírodě patří samice tuleně kroužkovaného, která se dožila 43 let, a samice tuleně kuželozubého, která v době smrti měla 46 roků.[128] Věk, ve kterém ploutvonožci dosahují pohlavní dospělosti, se pohybuje od 2–12 let v závislosti na druhu. Samice typicky dospívají dříve než samci.[129]
Potrava

Ploutvonožci jsou výhradně masožraví. Živí se hlavně rybami a hlavonožci, příležitostně korýši a mlži, občas i zooplanktonem, tučňáky či mořskými ptáky.[130] Většina ploutvonožců jsou potravní oportunisté a generalisté, pouze několik druhů představuje specialisty. Složení jídelníčku závisí na druhu i populaci, např. tuleni krabožraví se živí hlavně krunýřovkami, tuleni kroužkovaní pojídají převážně korýše, tuleni Rossovi a rypouši sloní se specializují na desetiramenatce a tuleni vousatí spolu s mroži pojídají převážně měkkýše a další bezobratlé živočichy zdržujících se u dna moře.[114]
Ploutvonožci loví samostatně nebo ve skupině. Samostatný lov je typický při pronásledování osamocených ryb, pomalých nebo nepohyblivých bezobratlých a během lovu teplokrevné kořisti. Druhy, které loví samostatně, se většinou vydávají za kořistí do pobřežních vod, zálivů a řek. Výjimku představuji rypouši, kteří se krmí v hlubokých oceánských vodách. Mroži se při krmení shlukují do neforemných skupin a i když se mohou vynořovat a ponořovat ve stejnou dobu, krmení probíhá samostatně. V případě dostupnosti rybích hejn nebo desetiramenatců dochází u některých druhů ploutvonožců ke kooperativnímu lovu. Zejména někteří lachtani se shlukují do velkých skupin a kořist nahánějí společným úsilím. Některé druhy jako lachtani kalifornští nebo lachtani hřivnatí občas loví v oblastech výskytu mořských ptáků a kytovců, kteří se společně stahují do oblastí bohatých na kořist.[131]

Ke konzumaci kořisti dochází obvykle přímo pod vodou. Ploutvonožci svou kořist příliš neporcují a obvykle ji celou spolknou. Větší úlovky si zatáhnou na břeh, kde je naporcují a snědí postupně.[132] Tuleni leopardí svou kořistí často násilně máchají ze strany na stranu, dokud ji neusmrtí.[133] Některé druhy, které získávají potravu filtrací vody (např. tuleň krabožravý), k tomu mají zvláště uzpůsobený chrup. Stoličky vedle hlavního hrotu mají ještě menší výstupky oddělené tenkými hlubokými rýhami. Při sevření tlamy do sebe zuby přesně zapadnou a průchodné zůstávají pouze úzké zářezy, kterými se filtruje kril, případně další malí živočichové.[134][114] Mrož je jedinečný v tom, že potravu sbírá nasáváním kořisti, kdy nejdříve nabere mlže do tlamy a poté za pomocí jazyka vysaje tělo živočicha z jeho schránky.[74] Většina ploutvonožců si shání potravu ve vodě, avšak najdou se i výjimky, jako je lachtan hřivnatý, který občas loví tučňáky i na souši.[135] Byl také zaznamenán případ lachtana novozélandského, který plenil hnízda albatrosů královských.[136] Některé druhy polykají kamínky či oblázky, a to patrně z důvodů podpoření rozmělňování potravy nebo kontrole parazitů (např. hlístic), ke které dochází jejich mechanickým rozdrcením.[137] I když jsou ploutvonožci schopni pít mořskou vodu, většinu tekutin získávají z těl své kořisti.[66]
Predátoři


Přestože ploutvonožci představují velké šelmy při vrcholu potravní pyramidy, i tak podléhají predaci. K jejich hlavním přirozeným predátorům patří hlavně kosatky. Tito kytovci loví ploutvonožce tak, že do nich narážejí hlavou, pleskají po nich ocasem a vymršťují je do vzduchu. Lov probíhá ve skupinách do 10 kosatek, výjimečně jich může být i více, případně i osamocená kosatka se může pustit do lovu ploutvonožce. Kosatky většinou cílí na mláďata, avšak mohou zaútočit i na dospělce. K dalším významným predátorům ploutvonožců patří velké druhy žraloků, jako je žralok bílý, tygří nebo mako. Útoky žraloků většinou nastávají zespodu ze zálohy. Ploutvonožcům se většinu žraločích útoků podaří odrazit, avšak ostré žraločí zuby často zanechávají na jejich tělech hluboké rány, které se časem promění ve viditelné jizvy. Lachtani většinou mívají zjizvení ve spodních částech těla, zatímco tuleni mívají jizvy spíše v částech horních.[138] Podle kousanců pozorovaných na rypouších sloních lze za predátora rypoušů považovat i žraloka Somniosus antarcticus z čeledi světlošovití (Somniosidae).[139] Na ploutvonožce cílí i někteří suchozemští predátoři, z nichž nejvýznamnější je medvěd lední, který se zaměřuje hlavně na mroží a tulení mláďata.[138] Na dospělé mrože si medvěd občas troufne také, nicméně mroži mohou svými kly medvěda i zabít, a tak se jim medvědi spíše vyhýbají.[140][141] K dalším suchozemským predátorům patří puma americká, hyena čabraková a některé druhy psovitých šelem. Stejně jako medvěd, i tito predátoři cílí hlavně na mláďata a mladší jedince.[138] U západního pobřeží Namibie se lvi pustinní naučili lovit lachtany jihoafrické.[142]

Ploutvonožci se někdy snaží chránit před predátory sdružováním do skupin.[143] Rypouši severní se vyhýbají predaci potopením do velkých hloubek, jelikož kosatky a žraloci bílí se krmí hlavně při povrchu (cca do 150 m).[113] V Antarktidě, kde se nevyskytují suchozemští predátoři, tráví ploutvonožci na zaledněných březích více času než ploutvonožci z oblastí Arktidy.[144] Antarktické druhy ploutvonožců se též zdají na břehu klidnější než jejich arktické protějšky. Arktičtí ploutvonožci téměř nikdy nekálí na ledu, patrně aby tak nepřilákali pozornost predátorů. Tuleni kroužkovaní si na obranu před predátory dokonce hloubí nory ve sněhové pokrývce, kde dochází k vrhům mláďat.[145][146]
U ploutvonožců občas nastává mezidruhová predace. Zejména tuleni leopardí jsou známí lovením mláďat ostatních ploutvonožců. Napadají zejména mláďata tuleňů krabožravých, která od listopadu do ledna tvoří důležitou součást jídelníčku tuleňů leopardích. Starší jedinci tuleňů krabožravých jsou často poseti jizvami od nezdařilých útoků tuleňů leopardích. Studie z roku 1977 zkoumala vzorek 85 tuleňů krabožravých a u 75 % z nich vědci našli zjizvení způsobené tuleni leopardími.[147] I když se mroži specializují na bezobratlé živočichy žijící při dně, příležitostně napadnou i jiné arktické ploutvonožce, které usmrtí svými dlouhými kly a poté sežerou jejich podkožní tuk a kůži. Lachtani ušatí byli pozorováni při požírání mláďat tuleňů obecných, rypoušů severních i lachtanů kalifornských. Lachtani novozélandští občas hodují na mláďatech jiných druhů lachtanů a lachtani hřivnatí se občas vydají na lov lachtanů jihoamerických.[138]
Rozmnožování
Pářící systém ploutvonožců se liší podle druhu a zahrnuje jak extrémní polygynii (jeden samec na více samic), tak sériovou monogamii (svazky na jedno pářící období, tvořené pouze jednou samicí a jedním samcem).[148] Z 33 druhů ploutvonožců se 20 rozmnožuje na souši a 13 na ledu.[149] Druhy, které se páří na souši, jsou většinou polygynní, protože samice se shromažďují ve velkých skupinách a samcům je tak umožněno se s nimi ve velkém pářit a zároveň samci dokáží samice ubránit před ostatními samci. K polygynním druhům patří rypouši, tuleni kuželozubí a většina lachtanovitých.[56] Druhy, které se rozmnožují na souši, se často páří na ostrovech, kde je méně suchozemských predátorů (na subantarktických ostrovech nejsou suchozemští predátoři vůbec). Vhodných stanovišť je pomálu a tak ty existující jsou mnohdy přecpána a zvířata jsou natěsnána těsně vedle sebe. V době páření nejdříve přicházejí na souš samci; samice připlavou až o něco později. Zatímco samice se rok od roku vracejí ke stejným břehům, samci si často k páření vybírají odlišná stanoviště od předchozího roku. U polygynních druhů se samci snaží spářit s co největším počtem samic. Samci některých druhů se v době říje dokonce po několik měsíců postí, protože kdyby si odskočili do vody na lov, je pravděpodobné, že by ztratili svou dominanci i samice.[150]
U polygynních druhů je vyvinut výrazný pohlavní dimorfismus. Samci bývají mnohem robustnější než samice, a to především v oblasti hrudníku a krku, jejich špičáky bývají delší a jejich srst bývá hustší. Všechny tyto vlastnosti přijdou samcům vhod při zápasech s ostatními samci o přízeň samic. Čím větší váhy samec dosáhne, tím déle může žít ze svých tukových zásob během půstu v období páření.[56] Robustnější velikost je výhodná i při lovu. Větší samci totiž mají lepší termoregulaci i větší množství energetických zásob, takže jsou schopni odolávat nižším teplotám některých bohatých krmišť.[57] Občas to jsou pouze ti největší z největších, kteří nakonec doplují do těch nejvzdálenějších krmišť či se potopí do hloubek bohatých na kořist, na kterou ostatní menší jedinci nedosáhnou.[151]
Ostatní ploutvonožci jako mrož a většina tuleňů tráví období říje na ledu a ke kopulacím dochází ve vodě (některé druhy, které tráví období rozmnožování na souši, však také kopulují ve vodě).[56][152][153] U těchto druhů se samice většinou neshlukují do skupin a následkem nestálosti ledu se pářící stanoviště nachází každý rok jinde, takže samci nemohou předvídat, kam přesně samice na počátku říje připlují. I z tohoto důvodu je polygynie u druhů, které tráví období říje na ledu, méně rozšířená. Výjimkou jsou pouze mrožice, které se shlukují do početných skupin, a to patrně z důvodů roztroušenosti potravních zdrojů. Ploutvonožci, kteří se rozmnožují na nepohyblivým ledu u pevniny, mají větší sklony ke shlukování než ti, kteří se množí na volně plovoucích krách.[152] Některé z druhů rozmnožujících se na ledu jsou sériově monogamní (tuleň obecný, tuleň krabožravý, čepcol hřebenatý).[154] Pohlavní dimorfismus bývá u druhů pářících se na ledu méně výrazný a u tuleňů z tribu Lobodontini jsou samice dokonce o něco větší než samci. Ze všech druhů rozmnožujících se na ledu to jsou pouze mroži a čepcoli hřebenatí, u nichž je pohlavní dimorfismus výrazný ve prospěch samců.[56][152]

K zajištění reprodukční úspěšnosti používají dospělí samci ploutvonožců několik strategií. Takoví lachtanovití zakládají teritoria, která zahrnují zdroje, na které nalákají samice. K těmto zdrojům patří dostatek stínu a malé bazénky k zchladnutí těl v parných letních dnech nebo snadný přístup k moři. Hranice mezi teritorii většinou představují přirozené přírodní prvky.[155] Některá teritoria mohou být částečně ponořená nebo dokonce úplně pod vodou.[156][157] Samci si svá teritoria bedlivě střeží proti ostatním samcům pomocí výhružných postojů a vokálních projevů. K fyzickým zápasům dochází jen zřídka.[158] Lachtani se každoročně přichází pářit na ta samá teritoria. Samci některých druhů (lachtan medvědí, lachtan ušatý) si dokáží podržet teritoria i 2–3 měsíce. U většiny druhů samci nedokáží zabránit samicím ve volném pohybu mezi teritorii ostatních samců, avšak i v tomto případě existují výjimky – samci lachtanů hřivnatých, medvědích a šedých dokáží samice udržet na svém teritoriu i proti jejich vůli. Samci některých tuleňovitých (tuleň vousatý, obecný a Weddellův) si zakládají teritoria i pod vodou v blízkosti samičích shromaždišť.[155] Taková teritoria jsou udržována hlavně vokalizací.[159] U lachtanů Weddelových se podvodní teritoria samců překrývají s dírami v ledu, kam se samice připlouvají nadechnout.[152]

Zejména u některých populací mrožů je vyvinut lekový systém páření.[155] Samci se shlukují kolem samic a snaží se na ně zapůsobit propracovanými namlouvacími pózami a barvitými hlasovými projevy.[155][160] Lekový systém se vyskytuje při nejmenším i u některých populací lachtanů jihoamerických,[161] lachtanů novozélandských a tuleňů obecných.[155][162] U některých druhů (rypouši, tuleni kuželozubí) se urostlí samci snaží uplatnit dominantní nárok na ostatní samice a zajistit si k nim výhradní přístup. Tyto samice pak brání proti ostatním samcům.[155] U rypoušů na začátku období říje dochází k vytvoření hierarchie založené na dominanci, která je často přímo úměrná mohutnosti samčích těl. Alfa samci si udržují harémy o velikosti několika desítek až nižších stovek samic, avšak u rypoušů sloních se v extrémních případech jedná i o více než tisíc samic.[163] Dominantní rypouši severní se těší nerušeným kopulacím se všemi samicemi v harému, který ovládají. Pokud se o páření pokusí některý ze subdominantních samců, alfa samci ho většinou zaženou.[164] Zatímco samci některých druhů (např. tuleni kuželozubí) si nárokují určité území v rámci skupiny samic (samice se na tomto území mohou střídat),[164] u některých jiných druhů (mroži, rypouši) si samci nárokují konkrétní skupinu samic.[155] Samci čepcolů hřebenatých, tuleňů grónských a tuleňů krabožravých následují a brání jednu až dvě kojící samice[165] a trpělivě čekají, až samice vstoupí do estrálního cyklu, aby se s nimi mohli spářit.[154][155]

Mladší a subdominantní samci ploutvonožců se také občas pokusí o kopulaci, avšak poněkud netradičními cestami. Nedospělí rypouši se občas pokusí proklouznout do harému samic, se kterými se snaží splynout zakrýváním či zatahováním svých chobotů. Často také obtěžují samice, které na konci období říje míří zpět do vody, a pokouší se s nimi spářit. U některých lachtanovitých, jako je lachtan jihoamerický či lachtan šedý, se nedospělí jedinci, kteří si nedokázali vybojovat teritorium, shlukují do skupin a občas proniknou do kolonie dominantního samce s cílem vyvolat chaos. Zmatečnou situaci se snaží využít pro zmocnění se některé z překvapených samic.[166]
Samice ploutvonožců mívají pouze omezené možnosti výběru, s jakým samcem se nakonec spáří. Lepší šanci mívají u druhů provozujících lekový systém, jako jsou některé populace mrožů. Samice rypoušů a tuleňů kuželozubých se v případě hrozby kopulace s nechtěným samcem brání vykrucováním a úprkem doprovázeným hlasitým štěkotem a pleskáním dotírajícího samce ocasem. Způsobený povyk často přiláká pozornost ostatních samců a ten nejdominantnější z nich nakonec většinou zažene subdominantního jedince pokoušejícího se o tento plíživý sex. Dominantní samec se nakonec pokusí sám spářit se samicí, které prvně ochotně přispěchal na pomoc.[160][167][168] Dominantní samice rypoušů severních zůstávají ve středu kolonie, kde mají lepší šanci se nakonec spářit s dominantním samcem, zatímco samicím při okrajích kolonie spíše hrozí, že se jich zmocní některý ze subdominantních samců.[169] Samice lachtanů ušatých se dominantním samcům dokonce aktivně podbízejí.[160]
Vrh a péče o mláďata
Takřka všechny samice ploutvonožců vstupují do estrálního cyklu krátce po vrhu a mohou tedy rodit každý rok.[170] Výjimku představují pouze mrožice, u nichž březost trvá kolem 16 měsíců a ovulace nastává až po vychování mláděte. Teoreticky tedy mrožice mohou vrhat každé dva roky, avšak zejména u starších samic je obvyklejší vrh každé 3–4 roky, případně i méně často.[171] Takřka u všech druhů probíhá odložená nidace, kdy sice dochází k oplodnění vajíčka, avšak to se neimplantuje do stěny dělohy hned, ale až za několik týdnů či měsíců. Díky odložené nidaci dochází k odložení vrhu do vhodnější doby, takže se samice stačí dostatečně vykrmit a rodí ve stejném – teplejším – období, kdy se přichází na břeh pářit.[170][172] Výjimkou jsou patrně lachtanice mořské, které žijí v prostředí s nevyzpytatelnou dostupností potravy, takže nemají potřebu svou březost optimalizovat.[173] Doba latence (období mezi oplodněním a uchycením vajíčka) trvá 1,5–5 měsíců v závislosti na druhu. Celková doba březosti (včetně období latence) trvá u většiny druhů kolem 10–12 měsíců, nejdéle trvá u mrožů (16 měsíců).[47][170] U většiny druhů se vrh odehrává v jarních a letních měsících.[174] V drtivé většině případů rodí samice pouze jedno mládě;[170] dvojčata se rodí pouze vzácně a když k tomu dojde, jejich úmrtnost je vysoká.[172] Mláďata jsou precociální, tzn. přicházejí na svět dobře vyvinuty.[170]
Na rozdíl od suchozemských savců mléko ploutvonožců neobsahuje laktózu.[175] Kojení a péče o mláďata se liší podle druhu. Některé samice tuleňovitých, jako jsou rypoušice, tulenice kuželozubé nebo samice čepcolů hřebenatých, zůstávají na souši nebo ledu po celou dobu laktace, během které se úplně postí. Např. u čepcolů to jsou jen čtyři dny, avšak u rypoušů to je kolem pěti týdnů. Mléko těchto druhů se skládá až ze 60 % z tuku, což mláďatům umožňuje rychle vyrůst. Mláďata rypoušů severních přibírají kolem 4 kg za den až do doby jejich osamostatnění. Některá mláďata těchto postních druhů se občas pokusí ukořistit mléko od ostatních kojících samic a nabrat tak váhu rychleji než jejich vrstevníci. U některých samic druhů, které se během laktace dlouhodobě postí, dochází občas k výchově nevlastních mláďat.[176][170]

U lachtanovitých a některých tuleňovitých (např. u tuleňů obecných) dochází k postění samic během laktace pouze na krátká několikadenní období, mezi nimiž se samice vydávají na moře nakrmit. Tyto lovné výpravy mohou trvat cca den až dva týdny v závislosti na dostupnosti potravy, resp. vzdálenosti krmiště. V době nepřítomnosti samic se mláďata postí.[170] Období kojení může u lachtanovitých trvat 6–11 měsíců; u lachtana galapážského může trvat dokonce až 3 roky. Mláďata těchto druhů v době osamostatnění váží méně než mláďata většiny tuleňovitých.[177] Mrožice jsou v rámci ploutvonožců jedinečné v tom, že svá mláďata krmí ve vodě.[170] Kojení mrožic probíhá plováním samice na vodní hladině ve vertikální (s hlavou nahoře) nebo horizontální poloze (v tomto případě s břichem dolů) a mládě hlavou dolů saje mléko.[178]
Mladí ploutvonožci se typicky naučí plavat sami. Zatímco některé druhy dovedou plavat bezprostředně po narození, jiným druhům trvá dny až týdny, než se poprvé odhodlají do vody. Mladí rypouši začínají plavat až několik týdnů po osamostatnění.[179] Samci ploutvonožců hrají ve výchově mláďat jen minimální úlohu.[180] Samci mrožů někdy pomáhají učit nezkušené mladé mrožíky plavat. Občas se dokonce starají o osiřelá mláďata.[181] Samci lachtanů kalifornských někdy chrání plavající mláďata před predátory.[182] Samci ploutvonožců nicméně mohou představovat pro mláďata i hrozbu. U druhů, které se páří na souši, může dojít ke zranění či usmrcení mláďat zalehnutím, a to zejména během samčích zápasů.[180] Nedospělí samci lachtanů jihoamerických občas unáší mláďata od jejich matek a zacházejí s nimi podobným způsobem jako dospělí samci zacházejí se samicemi. Toto chování jim patrně pomáhá v nabrání zkušeností, které využijí v pozdějších letech pro kontrolování samic. Pro mláďata takový únos končí v lepším případě traumatem, v horším případě zraněním či smrtí.[183]
Komunikace

Ploutvonožci mají poměrně bohatý hlasový projev zahrnující štěkání, chrochtání, vrčení, bručení, cvrlikání, supění či pískání. Většina zvuků je slyšitelná lidským uchem, avšak např. u tuleně leopardího bylo v zajetí zaznamenáno, jak pod vodou vydává zvuky ultrazvukové frekvence.[159] I tuleň Weddelův vydává zvuky v ultrazvukovém pásmu, i když význam těchto hlasových projevů zůstává nejasný.[184] U rypouše severního bylo během hlasového projevu zaznamenáno šíření infrazvukových vibrací. Lachtanovití jsou vokálnější na souši, zatímco tuleňovití se projevují hlavně ve vodě. Ploutvonožci obývající antarktické oblasti se hlasově projevují více než jejich arktičtí protějšci, a to z důvodu absence suchozemských predátorů.[159] Samčí volání mívají nižší frekvenci než samičí.[185]
Hlasové projevy ploutvonožců jsou zvláště důležité v období říje. Dominantní samci rypoušů používají svůj výrazný řev k vyjádření svého postavení i k zahánění soupeřů. Hlasitý řev rypoušů bývá zdůrazňován jejich kožním vakem na konci čenichu (zvaným též chobot).[186][187] K vokálním projevům samců lachtanovitých patří hlasitý štěkot, vrčení, řev a fňukání. Podobné zvuky umí vyprodukovat i samci mrožů, kteří nadto ještě dokáží pískat či vyprodukovat zvuk připomínající cinkání gongu, který používají k nalákání samic.[186][188]
Snad nejpropracovanější vokální projev má však tuleň Weddellův, který má oddělený repertoár pro suchozemské a vodní prostředí.[159] Jeho podvodní hlasový projev zahrnuje trylky, cvrlikání, frkání a klepání.[189] Vydávaný zvuk může trvat až 70 vteřin, což je na mořského savce dlouhá doba. Některá podvodní volání mají kolem sedmi rytmických vzorců a jsou připodobňovány k ptačímu nebo velrybímu zpěvu.[190] Výzkumná pracovnice z antarktické polární stanice McMurdo dokonce tyto netradiční zvuky přirovnala ke skupině Pink Floyd.[191] Podobná volání byla zaznamenána i u tuleňů z tribu Lobodontini[192] a u tuleňů vousatých.[193]
U některých druhů ploutvonožců byly zaznamenány geograficky odlišné „dialekty“,[194] u jiných druhů se zase vokální projevy liší jedinec od jedince.[195] Tyto individuální rozdíly jsou pravděpodobně důležité pro matky a jejich mláďata při udržování kontaktu na přecpaných plážích.[185] Samice lachtanovitých a jejich mláďata používají hlasitá kontaktní volání (provozovány zejména matkou) hlavně při návratu matky z lovecké výpravy na moři zpět na souš.[185][196] Samice rypoušů vábí svá mláďata rytmický voláním. V případě ohrožení jinými dospělci nebo když se mláďata snaží o sání mléka, tak vydávají arytmický ostrý řev.[197] Rypouší mláďata se hlasově projevují během hraní, při pobízení svých matek, aby jim dopřály mateřského mléka, nebo když se ocitnou v úzkých.[196][197]
Neverbální komunikace je u ploutvonožců spíše vzácná; častější je dokonce i u kytovců. Neverbálně komunikují např. tuleni bajkalští a obecní, kteří se při vyrušení vetřelci varovně pleskají předními ploutvemi o tělo, cvakají zuby, syčí nebo výrazně dýchají. Některé druhy se uchylují i k vizuálním postojům; např. tuleň Weddelův při hlídání svého podvodního teritoria někdy stáčí své tělo do písmene S a tuleň Rossův v případě pocitu ohrožení výstražně cení zuby a nápadně ukazuje své pruhování na hrudi.[159] Samci čepcolů hřebenatých používají své nafukovací vaky k vábení samic.[55]

Inteligence
Během pokusů s lachtany kalifornskými bylo zjištěno, že přinejmenším někteří lachtani dokáží porozumět elementární matematické symetrii, tranzitivitě a ekvivalenci.[198] Lachtani kalifornští též dokáží porozumět základní syntaxi a příkazům při učení umělé znakové řeči, nicméně sami je logicky používají pouze vzácně.[199] V roce 2011 byl u lachtana kalifornského chovaného v zajetí pozorováno, jak pokyvuje hlavou do rytmu puštěné hudby. Takové chování bylo do té doby zaznamenáno pouze u lidí, papoušků a některých ostatních druhů ptáků, kteří dokáží používat vokální mimikry (napodobování hlasem).[200] Ploutvonožci do určité míry dovedou napodobovat lidskou řeč. Tuleni obecní vychováni v zajetí si dovedli osvojit napodobování několika anglických slov a frází a dokonce i lidského smíchu.[201][202][203]
Lachtani chovaní v zajetí a v zábavních parcích jsou často cvičeni ve vykonávání určitých úkolů. K nejčastějším patří balancování míče na čenichu. Takový trik trvá lachtanovi zhruba rok, než se ho naučí. Jakmile je trik uložen v dlouhodobé paměti, lachtani ho mohou předvést i nejméně po třech měsících nenacvičování.[182][204]
U několika druhů (lachtani jihoameričtí, tuleni obecní, tuleni kuželozubí) byla testována paměť na opakování úkolů. Všechny druhy byly cvičeny vykonat určitý úkol (např. zamávání ploutví na příkaz cvičitele), a poté byli vycvičeni na příkaz „opakovat“, po němž jejích úkolem bylo opakovat poslední příkaz. Všechny druhy se tento příkaz dovedly naučit a opakovat poslední akci i v případě pauzy mezi posledním úkolem a opakovacím příkazem. Nicméně s délkou pauzy se snižovala úspěšnost opakování posledního úkolu a po 12–18 vteřinách se jednalo čistě o náhodu, zda ploutvonožci zopakují poslední příkaz.[205] Vedoucí výzkumu Simeon Smeele z Univerzity jižního Dánska to komentoval tak, že výsledky testu ukazují, že ploutvonožci do určité míry disponují vědomím, nicméně to ještě neprokazuje jejich sebeuvědomění.[206]
Vztah k lidem
Odkaz v kultuře

Ploutvonožci se odráží v lidské kultuře již po tisíciletí. Jedním z nejznámějších příkladů je bájné stvoření selkie, které nabírá v moři podobu tuleně a na souši podobu člověka. Mýty o selkie jsou tradovány hlavně v ostrovních oblastech severního Skotska jako jsou Hebridy či Orkneje.[207] Ploutvonožci mají silné zastoupení i v kultuře Inuitů.[208] V inuitské mytologii vystupuje bohyně Sedna, která vládne mořím a jejím živočichům. Bývá vyobrazována jako mořská panna, občas se spodní částí těla v podobě ploutvonožce. Podle jedné z inuitských legend ploutvonožci spolu s velrybami a ostatními mořskými savci pochází z useknutých prstů Sedny.[209] Ploutvonožci jsou zmiňování už v dílech Homéra a Aristotela a jedna z nejstarších dochovaných mincí starověkého Řecka vyobrazuje hlavu ploutvonožce. Řekové spojovali ploutvonožce s mořem i sluncem a za jejich ochránce považovali Poseidóna a Apollóna.[210] V kultuře jihoamerických Močiků se vyobrazení ploutvonožců často objevuje na jejich keramice a uměleckých předmětech.[211] V moderní kultuře jsou ploutvonožci považováni za roztomilé, hravé a komické postavy.[212] Postavy ploutvonožců se objevují v četných animovaných filmech, jako je Píseň moře, který je o selkie, či Pingu, kde vystupuje postava Rubyho.
Chov v zajetí

Ploutvonožci se pro svou velikost a hravou povahu vyskytují v chovných zařízeních po celém světě.[213] V zajetí byli chováni nejméně od dob starověkého Říma a jejich relativně snadné trénovatelnosti si všiml už Plinius starší. Francouzský zoolog Georges Cuvier v 19. století poznamenal, že divocí ploutvonožci vykazují nevídanou náklonnost k lidem a že ze všech zvířat si hned po opicích lze nejsnáze zkrotit právě tyto mořské šelmy. Francis Galton zase v jednom ze svých děl o domestikaci zvířat poznamenal, že ploutvonožci představují zvířata, u kterých s největší pravděpodobností nikdy nedojde k domestikaci, protože i přes jejich přátelskou povahu ploutvonožci nemají pro člověka praktické využití.[214]
Výběhy ploutvonožců v chovných zařízeních typicky zahrnují bazén, umělé shromaždiště, skály či kameny a případně i menší přístřešky. Propracovanější výběhy mají hluboké bazény s prosklenou stěnou, přes kterou mohou návštěvníci pozorovat chování ploutvonožců pod vodou. Nejčastěji chovaným druhem v zajetí je lachtan kalifornský, který je snadno trénovatelný i dobře přizpůsobivý. K dalším často chovaným druhům patří tuleň kuželozubý a tuleň obecný. Větší druhy jako mrož nebo lachtan ušatý se chovají podstatně méně.[213] Některé organizace jako je Světová společnost pro ochranu zvířat (World Animal Protection, WSPA) či Humánní společnost Spojených států (Humane Society of the United States, HSUS) nesouhlasí s držením ploutvonožců (ostatně i dalších mořských savců) v zajetí. Podle těchto organizací výběhy ploutvonožců nikdy nemohou být dostatečně velké na to, aby pojaly velká zvířata zvyklá migrovat na velké vzdálenosti, a navíc bazén nedokáže nahradit biodiverzitu oceánu. Organizace mají výhrady i k trikům předváděným ploutvonožci, které podle nich představují jen přehnané variace přirozeného chování a odvádějí pozornost lidí od skutečnosti, že se zvířata nachází v nepřirozeném prostředí.[215]
Lachtani kalifornští jsou využíváni námořnictvem Spojených států amerických pro vojenské účely. V rámci „Programu mořských savců Námořnictva Spojených států“ (United States Navy Marine Mammal Program, NMMP) jsou lachtani trénováni k detekci námořních min i potápěčů. V Perském zálivu byli lachtani vytrénováni nenápadně připlavat zezadu k nepřátelskému potápěči a během několika vteřin mu zaháknout za nohu svěrku s lanem. Vojáci pak mohou nepřátelského potápěče jednoduše vytáhnout z vody.[216] Organizace jako je PETA však takovéto praktiky kritizují s tím, že to zvířata vystavuje nebezpečí.[217] Podle amerického námořnictva jsou však lachtani (či trénovaní delfíni) vzati do bezpečí poté, co lokalizují miny či potápěče, zajištění těchto nepřátelských objektů probíhá výhradně cvičenými lidskými jednotkami americké armády.[218]
Lov

Lidé lovili ploutvonožce již od doby kamenné.[219][220] Maso ploutvonožců poskytovalo důležitý zdroj proteinů, železa, zinku i vitamínů A, D3 a B12.[221] Lovci původně zabíjeli dospělce a jejich mláďata na souši, avšak rozšíření harpun umožnilo lov zvířat i na moři z lodí. Někdy probíhal lov i s pomocí sítí. S rozmachem střelných zbraní a mořeplavectví v 18. a 19. století začal drasticky stoupat počet ulovených ploutvonožců. Hlavním cílem lovců bylo získání kožešiny a podkožního tuku. Zatímco kožešiny byly využívány v módním průmyslu, tuk se používal pro svícení do lamp nebo v průmyslové výrobě (např. k lubrikaci strojů a kůží, na výrobu lan či při výrobě textilií).[222][223] Z mrožů byly zpracovávány i jejich kly, do kterých se ryly ornamenty a poté byly výrobky prodávány jako dekorativní předměty.[222] Komerční lovci nejčastěji lovili tuleně grónské, čepcoly hřebenaté, tuleně kaspické, rypouše, mrože a všechny druhy lachtanů z podčeledi Arctocephalinae známé kvalitou srsti.[224].
Komerční lov ploutvonožců zažil rozmach mezi koncem 18. a druhou polovinou 19. století v závislosti na lokaci. Úspěch lovců závisel na výskytu ploutvonožců. Např. na Novém Zélandu zažil lov ploutvonožců rozmach v nultých letech 19. století, posléze upadl, aby se ve 20. letech dočkal nového rozmachu, načež znovu upadl.[225] V Kanadě se lov ploutvonožců těšil rozmachu mezi 10.–60. lety 19. století.[219] Většina populací ploutvonožců v 19. století drasticky poklesla (existují výjimky, např. tuleň Weddellův obývá natolik těžko dostupné oblasti Antarktidy, že se mu komerční lov vyhnul[226]), až se komerční rybolov přestal vyplácet – lovci se museli plavit za ploutvonožci do stále hůře přístupných oblastí, až se postupně tuleňářství stalo jednou z nejtěžších a nejnebezpečnějších námořních profesí. Lov ploutvonožců pokračoval i v průběhu 20. století, i když v mnohem menším měřítku[220] (např. v Jižní Georgii[227]). Většina komerčních aktivit ustala v 60. letech 20. století.[222]
Útlum komerčního lovu ploutvonožců byl způsoben vedle drastického úbytku zvířat i rozmachem komerčního používání potroleje a vynálezem elektřiny, které vedly ke konci poptávky po oleji ploutvonožců na svícení.[219] Další tlaky na lov ploutvonožců i ostatních mořských savců, zejména kytovců, byly rapidně sníženy s rozmachem syntetických materiálů a ropných produktů.[228] Ploutvonožcům pomohly i místní (národní) i mezinárodní ochranné úmluvy jako byla Úmluva o lachtanech ze severního Tichomoří (1911 North Pacific Fur Seal Convention) či Úmluva o zachování antarktických tuleňů z roku 1972.[228]
Po konci komerčního lovu začaly počty některých ploutvonožců opět stoupat. Například populace lachtanů antarktických se obnovila na počty před komerčním lovem. Rypouši severní byli koncem 19. století loveni takřka k vyhynutí a přežila jen malá populace na ostrově Guadalupe. Od té doby se však znovu rozšířili do míst svého původního výskytu, nicméně jejich genetická diverzita byla razantně snížena.[224] Naopak tuleni středomořští se do svých původních areálů, odkud byli vyhubeni, nikdy nevrátili a jejich roztříštěná populace obývá hlavně severovýchodní Středomoří a vody kolem severozápadní Afriky.[229]

Některé druhy ploutvonožců jsou nadále loveny. Úmluva o zachování antarktických tuleňů, která je součástí Antarktického smluvního systému, umožňuje omezený lov tuleňů krabožravých, tuleňů leopardích a tuleňů Weddelových, jiné druhy jako jsou rypouši sloní, tuleni Rossovi nebo lachtani antarktičtí naopak chrání.[224] Kanadská vláda povoluje lov tuleňů grónských, což se u části veřejnosti občas setkává s negativní odezvou. Zastánci lovu tuleňů argumentují tím, že zvířata jsou zabíjena humánně a mláďata s bílou srstí nejsou zabíjena vůbec. Odpůrci zase poukazují na to, že zabíjení tuleňů je nezodpovědné, protože už tak jsou ohroženi úbytkem svého habitatu.[230][231] Zvláštní postavení v otázce lovu ploutvonožců mají některé arktické národy, které na rozdíl od komerčních lovců často využívají ploutvonožce čistě pro své potřeby a jsou na nich do určité míry závislí pro vlastní přežití.[224] Jejich využití zvířat také bývá hospodárnější a méně destruktivní pro životní prostředí. Národní i mezinárodní organizace tak přisuzují některým domorodým národům zvláštní postavení a umožňují jim omezený lov. Výjimečnost postavení domorodých národů je nicméně někdy zpochybňována, jelikož i domorodé národy dnes již využívají moderní zbraně a motorizované metody dopravy a prodávají produkty z ploutvonožců. Každý rok příslušníci domorodých národů zabijí až 100 tisíc tuleňů (většinou tuleňů kroužkovaných) a cca 10 tisíc mrožů.[222]
Dva druhy ploutvonožců byly úplně vyhubeny následkem lovu člověkem, a sice tuleň karibský a lachtan japonský (tento druh je někdy považován za poddruh lachtana kalifornského). Tuleni karibští byli loveni evropskými osadníky již od konce 15. století. V místech výskytu tuleňů (Mexický záliv, Karibské moře, západní Atlantik) se nenacházelo mnoho suchozemských predátorů, takže tuleni byli přirozeně krotcí a představovali tak snadný terč pro organizované lovce, rybáře a bukanýry. Poslední věrohodný záznam o výskytu tuleně karibského pochází z roku 1952. IUCN prohlásil druh za vyhynulý v roce 1996.[232] Lachtan japonský se běžně vyskytoval u Japonského souostroví, avšak nadměrný lov spolu s prudkým nárůstem komerčního rybolovu způsobil drastické snížení populace. Poslední věrohodná pozorování druhu pochází ze 40. či 50. let 20. století. IUCN považuje druh za vyhynulý.[233]
Ohrožení a ochrana

K roku 2021 IUCN rozeznává 36 druhů ploutvonožců. Lachtan japonský a tuleň karibský jsou považováni za vyhynulé. Sedm druhů je považováno za ohrožené (tuleň havajský, středomořský a kaspický; lachtan galapážský, šedý, Forsterův a mořský), a tři za zranitelné (čepcol hřebenatý, lachtan medvědí, mrož lední).[22] Druhy žijící v polárních oblastech jsou silně náchylné na probíhající klimatické změny, zvláště pak na ústup mořského ledu.[234]
Některé druhy ploutvonožců se od doby zavedení jejich ochrany od 2. poloviny 20. století natolik rozšířily, že se dostávají do konfliktu s lidmi. Ve Spojených státech chrání ploutvonožce Zákon o ochraně mořských savců z roku 1972 (Marine Mammal Protection Act, MMPA). Od té doby se lachtani kalifornští opět hojně rozšířili po obou stranách Kalifornského poloostrova, kde často využívají k odpočinku lidské předměty a stavby, jako jsou přístavní doky a mola. Tyto stavby často nebyly navrženy k tomu, aby udržely váhu několika odpočívajících lachtanů. U řady doků proto došlo k přestavbě s cílem udržet váhu skupiny lachtanů.[235][236] Někteří ploutvonožci se dostávají do konfliktu i s rybáři, protože loví stejná hejna ryb.[237] V roce 2007 byl ke zmíněnému Zákonu o ochraně mořských savců schválen dodatek, který umožňuje usmrcení lachtanů v blízkosti migračních tras lososů na oregonské přehradě Bonneville na řece Columbii[238][239] (vedle lososů čavyča jsou ohroženy i některé druhy jeseterů, mihulotvarých, pstruhů a koruška americká).[239][240] Cílem tohoto dodatku je snížit tlaky, kterým ubývající populace lososů čelí.[241] Místní ochránci se snažili zabránit lachtanům v hodování na lososech pomocí podvodních výbušnin (cca 8 cm dlouhá výbušnina připomínající velkou petardu, která exploduje pod vodou a vydá hlasitý zvuk, který se vodou šíří desítky kilometrů daleko[242]) i gumových projektilů, avšak žádná z těchto metod nebyla úspěšná.[243][244] Kritici odstřelu lachtanů jako Humánní společnost Spojených států namítají, že vodní elektrárny představují pro lososy mnohem větší hrozbu než lachtani.[243] Podobný konflikt existuje i v Jihoafrické republice s lachtany jihoafrickými. V 80. a 90. letech 20. století někteří jihoafričtí politici a rybáři požadovali zabíjení lachtanů, jelikož měli mít negativní dopad na místní rybářský průmysl.[245] Podobně také australský politik Adrian Pederick navrhl omezený lov lachtanů v některých oblastech Austrálie.[246]
Ploutvonožci jsou ohrožení lidskými aktivitami i nepřímo. Občas představují vedlejší úlovky komerčních rybářských operací (typicky se zamotají do sítí nebo omylem spolknou rybářský háček). Zvláště nevody a tenatové sítě způsobují vysokou úmrtnost ploutvonožců a i dalších mořských savců. Ke druhům nejčastěji postiženým komerčním rybářstvím patří lachtani kalifornští, medvědí, jihoafričtí a tuleni havajští.[224] Na ploutvonožce má též negativní vliv znečištění moří a oceánů. Jelikož mají ploutvonožci silnou vrstvu podkožního tuku a nachází se na vrcholu potravní pyramidy, v jejich tělech se často koncentrují perzistentní organické látky. Tyto chemikálie mohou způsobit rakovinu zažívacího traktu, snížit schopnost reprodukce i zredukovat odolnost vůči infekčním chorobám. Kojící samice mohou tyto chemikálie přenést na svá mláďata.[247] K dalším hrozbám pro ploutvonožce patří ničení jejich přirozeného životního prostředí ropnými a plynovými plošinami, srážky s loděmi[224] či podvodní hluk.[248]
Odkazy
Reference
V tomto článku byl použit překlad textu z článku Pinniped na anglické Wikipedii.
- List of Marine Mammal Species and Subspecies [online]. The Society for Marine Mammalogy [cit. 2021-04-21]. Dostupné online. (anglicky)
- LINNÉ, Carl von. Caroli Linnæi Systema naturæ. Švédsko: Lipsiæ, Sumptibus Guilielmi Engelmann, 1758. Dostupné online. (latinsky)
- Allen, J. A. History of North American pinnipeds, a monograph of the walruses, sea-lions, sea-bears and seals of North America. Washington: Government Printing Office, 1880. Dostupné online. (anglicky)
- Scheffer 1958.
- ELIAS, Joseph S. Science terms made easy : a lexicon of scientific words and their root language origins. Westport, Connecticut.: Greenwood Press, 2007. Dostupné online. ISBN 978-0-313-33896-0. S. 157. (anglicky)
- KORETSKY, I. A.; BARNES, L. G.; RAHMAT, S. J. Re-Evaluation of Morphological Characters Questions Current Views of Pinniped Origins. S. 327–354. Vestnik Zoologii [online]. 2016-08-01 [cit. 2021-04-17]. Roč. 50, čís. 4, s. 327–354. DOI 10.1515/vzoo-2016-0040. (anglicky)
- Encyclopædia Britannica. Spojené státy americké: William Benton, 1966. Kapitola Carnivora. (anglicky)
- SIMPSON, George Gaylord. The Principles of Classification and Classification of Mammals. New York: American Museum of Natural History (Volume 85), 1945. Dostupné online. (anglicky)
- BERTA, A.; RAY, C. E.; WYSS, A. R. Skeleton of the Oldest Known Pinniped, Enaliarctos mealsi. S. 60–62. Science [online]. 1989-04-07 [cit. 2021-02-27]. Roč. 244, čís. 4900, s. 60–62. DOI 10.1126/science.244.4900.60. (anglicky)
- BERTA, Annalisa. New Specimens of the Pinnipediform Pteronarctos from the Miocene of Oregon. S. 1–30. Smithsonian Contributions to Paleobiology [online]. 1994 [cit. 2021-06-01]. Čís. 78, s. 1–30. Dostupné online. DOI 10.5479/si.00810266.78.1. (anglicky)
- PATERSON, Ryan S.; RYBCZYNSKI, Natalia; KOHNO, Naoki; MADDIN, Hillary C. A Total Evidence Phylogenetic Analysis of Pinniped Phylogeny and the Possibility of Parallel Evolution Within a Monophyletic Framework. S. 457. Frontiers in Ecology and Evolution [online]. 2020-01-17 [cit. 2021-05-26]. Roč. 7, s. 457. DOI 10.3389/fevo.2019.00457. (anglicky)
- YONEZAWA, Takahiro; NIKAIDO, Masato; KOHNO, Naoki; FUKUMOTO, Yukio; OKADA, Norihiro; HASEGAWA, Masami. Molecular phylogenetic study on the origin and evolution of Mustelidae. S. 1–12. Gene [online]. 2007-07 [cit. 2021-06-01]. Roč. 396, čís. 1, s. 1–12. DOI 10.1016/j.gene.2006.12.040. (anglicky)
- DELISLE, Isabelle; STROBECK, Curtis. A phylogeny of the Caniformia (order Carnivora) based on 12 complete protein-coding mitochondrial genes. S. 192–201. Molecular Phylogenetics and Evolution [online]. 2005-10 [cit. 2021-06-01]. Roč. 37, čís. 1, s. 192–201. DOI 10.1016/j.ympev.2005.04.025. (anglicky)
- AGNARSSON, Ingi; KUNTNER, Matjaž; MAY-COLLADO, Laura J. Dogs, cats, and kin: A molecular species-level phylogeny of Carnivora. S. 726–745. Molecular Phylogenetics and Evolution [online]. 2010-03 [cit. 2021-06-01]. Roč. 54, čís. 3, s. 726–745. DOI 10.1016/j.ympev.2009.10.033. (anglicky)
- ARNASON, Ulfur; GULLBERG, Anette; JANKE, Axel; KULLBERG, Morgan. Mitogenomic analyses of caniform relationships. S. 863–874. Molecular Phylogenetics and Evolution [online]. 2007-12 [cit. 2021-02-27]. Roč. 45, čís. 3, s. 863–874. DOI 10.1016/j.ympev.2007.06.019. (anglicky)
- Superfamily Otarioidea Lucas 1899 [online]. Paleobiology Database [cit. 2021-02-27]. Dostupné online. (anglicky)
- Berta, Annalisa. „Pinnipedia, overview“, s. 1148–1152. In Perrin, Würsig a Thewissen 2009.
- Superfamily Phocoidea Smirnov 1908 [online]. Paleobiology Database [cit. 2021-02-27]. Dostupné online. (anglicky)
- BERTA, Annalisa; CHURCHILL, Morgan. Pinniped taxonomy: review of currently recognized species and subspecies, and evidence used for their description: Pinniped taxonomy: evidence for species and subspecies. S. 207–234. Mammal Review [online]. 2012-07 [cit. 2021-02-02]. Roč. 42, čís. 3, s. 207–234. DOI 10.1111/j.1365-2907.2011.00193.x. (anglicky)
- NYAKATURA, Katrin; BININDA-EMONDS, Olaf RP. Updating the evolutionary history of Carnivora (Mammalia): a new species-level supertree complete with divergence time estimates. S. 12. BMC Biology [online]. 2012-12 [cit. 2021-02-28]. Roč. 10, čís. 1, s. 12. DOI 10.1186/1741-7007-10-12. (anglicky)
- Berta, Sumich a Kovacs 2006, s. 27.
- Taxonomy Search: Phocidae, Otarridae, Odobenidae [online]. IUCN Red List of Threatened Species [cit. 2021-03-17]. Reference založena na hledání pomocí tlačítka "Taxonomy" v levém panelu. Dostupné online. (anglicky)
- DEMÉRÉ, Thomas A.; BERTA, Annalisa; ADAM, Peter J. Chapter 3. S. 32–76. Bulletin of the American Museum of Natural History [online]. 2003 [cit. 2021-02-28]. Roč. 2003, čís. 279, s. 32–76. DOI 10.1206/0003-0090(2003)279<0032:C>2.0.CO;2. (anglicky)
- HIGDON, Jeff W.; BININDA-EMONDS, Olaf RP; BECK, Robin MD; FERGUSON, Steven H. Phylogeny and divergence of the pinnipeds (Carnivora: Mammalia) assessed using a multigene dataset. BMC Ecology and Evolution [online]. 2007 [cit. 2021-05-28]. Roč. 7. Dostupné online. DOI 10.1186/1471-2148-7-216. (anglicky)
- Riedman 1990, s. 64.
- Riedman 1990, s. 68–70.
- WYNEN, Louise P.; GOLDSWORTHY, Simon D.; INSLEY, Stephen J.; ADAMS, Mark; BICKHAM, John W.; FRANCIS, John; GALLO, Juan Pablo. Phylogenetic Relationships within the Eared Seals (Otariidae: Carnivora): Implications for the Historical Biogeography of the Family. S. 270–284. Molecular Phylogenetics and Evolution [online]. 2001-11 [cit. 2021-02-28]. Roč. 21, čís. 2, s. 270–284. DOI 10.1006/mpev.2001.1012. (anglicky)
- ARNASON, Ulfur; GULLBERG, Anette; JANKE, Axel; KULLBERG, Morgan; LEHMAN, Niles; PETROV, Evgeny A.; VÄINÖLÄ, Risto. Pinniped phylogeny and a new hypothesis for their origin and dispersal. S. 345–354. Molecular Phylogenetics and Evolution [online]. 2006-11 [cit. 2021-02-28]. Roč. 41, čís. 2, s. 345–354. DOI 10.1016/j.ympev.2006.05.022. (anglicky)
- Riedman 1990, s. 82–83.
- Anděra 2003, s. 76.
- Berta, Annalisa. „Pinniped evolution“, s. 861–866. In Perrin, Würsig a Thewissen 2009.
- BERTA, Annalisa; CHURCHILL, Morgan; BOESSENECKER, Robert W. The Origin and Evolutionary Biology of Pinnipeds: Seals, Sea Lions, and Walruses. S. 203–228. Annual Review of Earth and Planetary Sciences [online]. 2018-05-30 [cit. 2021-03-02]. Roč. 46, čís. 1, s. 203–228. DOI 10.1146/annurev-earth-082517-010009. (anglicky)
- Kirkwood 2013, s. 8.
- LENTO, G. M.; HICKSON, R. E.; CHAMBERS, G. K.; PENNY, D. Use of spectral analysis to test hypotheses on the origin of pinnipeds.. S. 28–52. Molecular Biology and Evolution [online]. 1995-01-01 [cit. 2021-03-02]. Roč. 12, čís. 1, s. 28–52. DOI 10.1093/oxfordjournals.molbev.a040189. (anglicky)
- Hunt, R. M. Jr.; Barnes, L. G. Basicranial evidence for ursid affinity of the oldest pinnipeds. Proceedings of the San Diego Society of Natural History. 1994, roč. 29, s. 57–67. Dostupné online. (anglicky)
- HIGDON, Jeff W; BININDA-EMONDS, Olaf RP; BECK, Robin MD; FERGUSON, Steven H. Phylogeny and divergence of the pinnipeds (Carnivora: Mammalia) assessed using a multigene dataset. S. 216. BMC Evolutionary Biology [online]. 2007 [cit. 2021-03-02]. Roč. 7, čís. 1, s. 216. DOI 10.1186/1471-2148-7-216. (anglicky)
- SATO, Jun J.; WOLSAN, Mieczyslaw; SUZUKI, Hitoshi; HOSODA, Tetsuji; YAMAGUCHI, Yasunori; HIYAMA, Kozue; KOBAYASHI, Mari. Evidence from Nuclear DNA Sequences Sheds Light on the Phylogenetic Relationships of Pinnipedia: Single Origin with Affinity to Musteloidea. S. 125–146. Zoological Science [online]. 2006-02 [cit. 2021-03-02]. Roč. 23, čís. 2, s. 125–146. DOI 10.2108/zsj.23.125. (anglicky)
- FLYNN, John J.; FINARELLI, John A.; ZEHR, Sarah; HSU, Johnny; NEDBAL, Michael A. Molecular Phylogeny of the Carnivora (Mammalia): Assessing the Impact of Increased Sampling on Resolving Enigmatic Relationships. S. 317–337. Systematic Biology [online]. 2005-04-01 [cit. 2021-03-02]. Roč. 54, čís. 2, s. 317–337. DOI 10.1080/10635150590923326. (anglicky)
- HAMMOND, John A.; HAUTON, Chris; BENNETT, Kimberley A.; HALL, Ailsa J. Phocid Seal Leptin: Tertiary Structure and Hydrophobic Receptor Binding Site Preservation during Distinct Leptin Gene Evolution. S. e35395. PLoS ONE [online]. 2012-04-19 [cit. 2021-03-02]. Roč. 7, čís. 4, s. e35395. DOI 10.1371/journal.pone.0035395. (anglicky)
- RYBCZYNSKI, Natalia; DAWSON, Mary R.; TEDFORD, Richard H. A semi-aquatic Arctic mammalian carnivore from the Miocene epoch and origin of Pinnipedia. S. 1021–1024. Nature [online]. 2009-04 [cit. 2021-03-02]. Roč. 458, čís. 7241, s. 1021–1024. DOI 10.1038/nature07985. (anglicky)
- Paleo-Breakthrough (Seal Evolution) from an Arctic Fossil [online]. Canadian Museum of Nature [cit. 2021-03-02]. Dostupné online. (anglicky)
- CULLEN, Thomas M.; FRASER, Danielle; RYBCZYNSKI, Natalia; SCHRÖDER-ADAMS, Claudia. EARLY EVOLUTION OF SEXUAL DIMORPHISM AND POLYGYNY IN PINNIPEDIA: EVOLUTION OF SEXUAL DIMORPHISM IN PINNIPEDS. S. 1469–1484. Evolution [online]. 2014-05. Roč. 68, čís. 5, s. 1469–1484. DOI 10.1111/evo.12360. (anglicky)
- FYLER, C. A.; REEDER, T. W.; BERTA, A.; ANTONELIS, G.; AGUILAR, A.; ANDROUKAKI, E. Historical biogeography and phylogeny of monachine seals (Pinnipedia: Phocidae) based on mitochondrial and nuclear DNA data: Monachine seal historical biogeography. S. 1267–1279. Journal of Biogeography [online]. 2005-06-13 [cit. 2021-03-02]. Roč. 32, čís. 7, s. 1267–1279. DOI 10.1111/j.1365-2699.2005.01281.x. (anglicky)
- BOESSENECKER, Rw; CHURCHILL, M. The origin of elephant seals: implications of a fragmentary late Pliocene seal (Phocidae: Miroungini) from New Zealand. S. 544–550. New Zealand Journal of Geology and Geophysics [online]. 2016-10-01 [cit. 2021-03-02]. Roč. 59, čís. 4, s. 544–550. DOI 10.1080/00288306.2016.1199437. (anglicky)
- †Prototaria Takeyama and Ozawa 1984 (walrus) [online]. Fossilworks [cit. 2021-03-02]. Dostupné v archivu pořízeném dne 2021-03-21. (anglicky)
- †Proneotherium Kohno et al. 1995 (walrus) [online]. Fossilworks [cit. 2021-03-02]. Dostupné v archivu pořízeném dne 2021-03-21. (anglicky)
- Berta, Annalisa. „Pinnipedia, overview“, s. 881–884. In Perrin, Würsig a Thewissen 2009.
- KARLESKIN, G.; TURNER, R. L.; SMALL, J. W. Introduction to Marine Biology. Pacific Grove, Kalifornie: Cengage Learning, 2009. ISBN 978-0-495-56197-2. S. 328. (anglicky)
- Berta, Sumich a Kovacs 2006, s. 165.
- Scheffer 1958, s. 16–17.
- Riedman 1990, s. 162–164.
- GOODMAN, S. Pusa sibirica [online]. The IUCN Red List of Threatened Species 2016, 2015-07-10. DOI 10.2305/IUCN.UK.2016-1.RLTS.T41676A45231738.en. (anglicky)
- PÁEZ-ROSAS, Diego; RIOFRÍO-LAZO, Marjorie; ORTEGA, Jorge; MORALES, Juan de Dios; CARVAJAL, Raúl; ALAVA, Juan José. Southern elephant seal vagrants in Ecuador: a symptom of La Niña events?. S. 13. Marine Biodiversity Records [online]. 2018-12 [cit. 2021-03-03]. Roč. 11, čís. 1, s. 13. DOI 10.1186/s41200-018-0149-y. (anglicky)
- MCCANN, T. S.; FEDAK, M. A.; HARWOOD, J. Parental investment in southern elephant seals, Mirounga leonina. S. 81–87. Behavioral Ecology and Sociobiology [online]. 1989-08 [cit. 2021-01-31]. Roč. 25, čís. 2, s. 81–87. DOI 10.1007/BF00302924. (anglicky)
- Berta 2012, s. 73–74.
- Ralls, K.; Mesnick, S. „Sexual dimorphism“, s. 1005–1011. In Perrin, Würsig a Thewissen 2009.
- GONZÁLEZ-SUÁREZ, Manuela; CASSINI, Marcelo H. Variance in male reproductive success and sexual size dimorphism in pinnipeds: testing an assumption of sexual selection theory: Does sexual selection explain size dimorphism?. S. 88–93. Mammal Review [online]. 2014-04 [cit. 2021-03-09]. Roč. 44, čís. 2, s. 88–93. DOI 10.1111/mam.12012. (anglicky)
- KRÜGER, Oliver; WOLF, Jochen B. W.; JONKER, Rudy M.; HOFFMAN, Joseph I.; TRILLMICH, Fritz. DISENTANGLING THE CONTRIBUTION OF SEXUAL SELECTION AND ECOLOGY TO THE EVOLUTION OF SIZE DIMORPHISM IN PINNIPEDS: SIZE DIMORPHISM AND MATING SYSTEM IN SEALS. S. 1485–1496. Evolution [online]. 2014-05 [cit. 2021-03-03]. Roč. 68, čís. 5, s. 1485–1496. DOI 10.1111/evo.12370. (anglicky)
- Riedman 1990, s. 3–9.
- Riedman 1990, s. 16.
- Berta 2012, s. 62.
- TKADLEC, Emil. Populační ekologie : struktura, růst a dynamika populací. Olomouc: Univerzita Palackého v Olomouci, 2008. ISBN 978-80-244-2149-0. S. 297.
- Riedman 1990, s. 253–255.
- Kirkwood 2013, s. 19–20.
- Berta, Sumich a Kovacs 2006, s. 317.
- Riedman 1990, s. 31–32.
- Berta 2012, s. 62–64.
- Anděra 2003, s. 72.
- FISH, F. E. Maneuverability by the sea lion Zalophus californianus: turning performance of an unstable body design. S. 667–674. Journal of Experimental Biology [online]. 2003-02-15 [cit. 2021-03-04]. Roč. 206, čís. 4, s. 667–674. DOI 10.1242/jeb.00144. (anglicky)
- FISH, Frank E. Transitions from Drag-based to Lift-based Propulsion in Mammalian Swimming. S. 628–641. American Zoologist [online]. 1996-12 [cit. 2021-03-04]. Roč. 36, čís. 6, s. 628–641. DOI 10.1093/icb/36.6.628. (anglicky)
- Riedman 1990, s. 11–12.
- ENGLISH, Arthur Wm. Limb movements and locomotor function in the California sea lion (Zalophus californianus). S. 341–364. Journal of Zoology [online]. 2009-08-20 [cit. 2021-03-04]. Roč. 178, čís. 3, s. 341–364. DOI 10.1111/j.1469-7998.1976.tb02274.x. (anglicky)
- Riedman 1990, s. 43–45.
- Berta 2012, s. 67.
- HANKE, Frederike D.; HANKE, Wolf; SCHOLTYSSEK, Christine; DEHNHARDT, Guido. Basic mechanisms in pinniped vision. S. 299–311. Experimental Brain Research [online]. 2009-12 [cit. 2021-03-04]. Roč. 199, čís. 3–4, s. 299–311. DOI 10.1007/s00221-009-1793-6. (anglicky)
- MASS, Alla M.; SUPIN, Alexander YA. Adaptive features of aquatic mammals' eye. S. 701–715. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology [online]. 2007-06 [cit. 2021-03-04]. Roč. 290, čís. 6, s. 701–715. DOI 10.1002/ar.20529. (anglicky)
- Lavigne, D. M. „Harp seal“, s. 542–546. In Perrin, Würsig a Thewissen 2009.
- GRIEBEL, Ulrike; PEICHL, Leo. Colour vision in aquatic mammals—facts and open questions. S. 18–30. Aquatic Mammals [online]. 2003-01-01 [cit. 2021-03-04]. Roč. 29, čís. 1, s. 18–30. DOI 10.1578/016754203101024040. (anglicky)
- Kastelein, R. A. „Walrus“, s. 1212–1216. In Perrin, Würsig a Thewissen 2009.
- Riedman 1990, s. 49.
- Riedman 1990, s. 39.
- KASTAK, David; SCHUSTERMAN, Ronald J. Low-frequency amphibious hearing in pinnipeds: Methods, measurements, noise, and ecology. S. 2216–2228. The Journal of the Acoustical Society of America [online]. 1998-04 [cit. 2021-03-04]. Roč. 103, čís. 4, s. 2216–2228. DOI 10.1121/1.421367. (anglicky)
- KOWALEWSKY, Sylvia; DAMBACH, Martin; MAUCK, Björn; DEHNHARDT, Guido. High olfactory sensitivity for dimethyl sulphide in harbour seals. S. 106–109. Biology Letters [online]. 2006-03-22 [cit. 2021-03-04]. Roč. 2, čís. 1, s. 106–109. DOI 10.1098/rsbl.2005.0380. (anglicky)
- Riedman 1990, s. 40.
- SCHUSTERMAN, Ronald J.; KASTAK, David; LEVENSON, David H.; REICHMUTH, Colleen J.; SOUTHALL, Brandon L. Why pinnipeds don’t echolocate. S. 2256–2264. The Journal of the Acoustical Society of America [online]. 2000-04 [cit. 2021-03-04]. Roč. 107, čís. 4, s. 2256–2264. DOI 10.1121/1.428506. (anglicky)
- GRANT, Robyn; WIESKOTTEN, Sven; WENGST, Nina; PRESCOTT, Tony; DEHNHARDT, Guido. Vibrissal touch sensing in the harbor seal (Phoca vitulina): how do seals judge size?. S. 521–533. Journal of Comparative Physiology A [online]. 2013-06 [cit. 2021-03-04]. Roč. 199, čís. 6, s. 521–533. DOI 10.1007/s00359-013-0797-7. (anglicky)
- MIERSCH, L.; HANKE, W.; WIESKOTTEN, S.; HANKE, F. D.; OEFFNER, J.; LEDER, A.; BREDE, M. Flow sensing by pinniped whiskers. S. 3077–3084. Philosophical Transactions of the Royal Society B: Biological Sciences [online]. 2011-11-12 [cit. 2021-03-04]. Roč. 366, čís. 1581, s. 3077–3084. DOI 10.1098/rstb.2011.0155. (anglicky)
- DEHNHARDT, G. Hydrodynamic Trail-Following in Harbor Seals (Phoca vitulina). S. 102–104. Science [online]. 2001-07-06 [cit. 2021-03-04]. Roč. 293, čís. 5527, s. 102–104. DOI 10.1126/science.1060514. (anglicky)
- SCHULTE-PELKUM, N.; WIESKOTTEN, S.; HANKE, W.; DEHNHARDT, G.; MAUCK, B. Tracking of biogenic hydrodynamic trails in harbour seals (Phoca vitulina). S. 781–787. Journal of Experimental Biology [online]. 2007-03-01 [cit. 2021-03-04]. Roč. 210, čís. 5, s. 781–787. DOI 10.1242/jeb.02708. (anglicky)
- HYVÄRINEN, Heikki. Diving in darkness: whiskers as sense organs of the ringed seal (Phoca hispida saimensis). S. 663–678. Journal of Zoology [online]. 1989-08 [cit. 2021-03-04]. Roč. 218, čís. 4, s. 663–678. DOI 10.1111/j.1469-7998.1989.tb05008.x. (anglicky)
- MURPHY, Christin T.; EBERHARDT, William C.; CALHOUN, Benton H.; MANN, Kenneth A.; MANN, David A. Effect of Angle on Flow-Induced Vibrations of Pinniped Vibrissae. S. e69872. PLoS ONE [online]. 2013-07-26 [cit. 2021-03-05]. Roč. 8, čís. 7, s. e69872. DOI 10.1371/journal.pone.0069872. (anglicky)
- GINTER, Carly C.; FISH, Frank E.; MARSHALL, Christopher D. Morphological analysis of the bumpy profile of phocid vibrissae. Marine Mammal Science [online]. 2009-12 [cit. 2021-03-05]. Roč. 26, čís. 3. DOI 10.1111/j.1748-7692.2009.00365.x. (anglicky)
- RENOUF, D. The Behaviour of pinnipeds. London: Chapman and Hall, 1991. ISBN 978-0-412-30540-5. S. 373. (anglicky)
- Riedman 1990, s. 42.
- Riedman 1990, s. 25.
- Berta 2012, s. 69.
- Berta, Sumich a Kovacs 2006, s. 245.
- KOOYMAN, G L; CASTELLINI, M A; DAVIS, R W. Physiology of Diving in Marine Mammals. S. 343–356. Annual Review of Physiology [online]. 1981-10 [cit. 2021-03-05]. Roč. 43, čís. 1, s. 343–356. DOI 10.1146/annurev.ph.43.030181.002015. (anglicky)
- MILLER, Natalie J.; POSTLE, Anthony D.; ORGEIG, Sandra; KOSTER, Grielof; DANIELS, Christopher B. The composition of pulmonary surfactant from diving mammals. S. 152–168. Respiratory Physiology & Neurobiology [online]. 2006-06 [cit. 2021-03-05]. Roč. 152, čís. 2, s. 152–168. DOI 10.1016/j.resp.2005.08.001. (anglicky)
- DENISON, D.M.; KOOYMAN, G.L. The structure and function of the small airways in pinniped and sea otter lungs. S. 1–10. Respiration Physiology [online]. 1973-01. Roč. 17, čís. 1, s. 1–10. DOI 10.1016/0034-5687(73)90105-9. (anglicky)
- COSTA, Daniel P. Diving Physiology of Marine Vertebrates. S. a0004230. Encyclopedia of Life Sciences [online]. 2007-01-15 [cit. 2021-03-05]. S. a0004230. Dostupné online. DOI 10.1002/9780470015902.a0004230. (anglicky)
- Berta, Sumich a Kovacs 2006, s. 241.
- Gili, Meijer a Lacave 2018, s. 14–16.
- CHAISE, Laureline L.; MCCAFFERTY, Dominic J.; KRELLENSTEIN, Adélie; GALLON, Susan L.; PATERSON, William D.; THÉRY, Marc; ANCEL, André. Environmental and physiological determinants of huddling behavior of molting female southern elephant seals (Mirounga leonina). S. 182–190. Physiology & Behavior [online]. 2019-02 [cit. 2021-01-30]. Roč. 199, s. 182–190. DOI 10.1016/j.physbeh.2018.10.016. (anglicky)
- Riedman 1990, s. 14.
- Berta 2012, s. 65.
- Berta, Sumich a Kovacs 2006, s. 220.
- CHAISE, Laureline L.; PRINET, Iris; TOSCANI, Camille; GALLON, Susan L.; PATERSON, William; MCCAFFERTY, Dominic J.; THÉRY, Marc. Local weather and body condition influence habitat use and movements on land of molting female southern elephant seals ( Mirounga leonina ). S. 6081–6090. Ecology and Evolution [online]. 2018-06 [cit. 2021-03-05]. Roč. 8, čís. 12, s. 6081–6090. DOI 10.1002/ece3.4049. (anglicky)
- MAUCK, B. Thermal windows on the trunk of hauled-out seals: hot spots for thermoregulatory evaporation?. S. 1727–1738. Journal of Experimental Biology [online]. 2003-05-15 [cit. 2021-03-05]. Roč. 206, čís. 10, s. 1727–1738. DOI 10.1242/jeb.00348. (anglicky)
- MASCETTI, Gian Gastone. Unihemispheric sleep and asymmetrical sleep: behavioral, neurophysiological, and functional perspectives. S. 221–238. Nature and Science of Sleep [online]. 2016-07-12 [cit. 2021-01-20]. Čís. 8, s. 221–238. DOI 10.2147/NSS.S71970. (anglicky)
- TOMAN, Jan. Jaká je role REM spánku? Záhadu pomáhají vyřešit lachtani [online]. Nedd.cz, 2018-06-25 [cit. 2021-01-20]. Dostupné online.
- LAPIERRE, J. L.; KOSENKO, P. O.; KODAMA, T.; PEEVER, J. H.; MUKHAMETOV, L. M.; LYAMIN, O. I.; SIEGEL, J. M. Symmetrical Serotonin Release during Asymmetrical Slow-Wave Sleep: Implications for the Neurochemistry of Sleep-Waking States. S. 2555–2561. Journal of Neuroscience [online]. 2013-02-06 [cit. 2021-03-05]. Roč. 33, čís. 6, s. 2555–2561. DOI 10.1523/JNEUROSCI.2603-12.2013. (anglicky)
- MITANI, Yoko; ANDREWS, Russel D.; SATO, Katsufumi; KATO, Akiko; NAITO, Yasuhiko; COSTA, Daniel P. Three-dimensional resting behaviour of northern elephant seals: drifting like a falling leaf. S. 163–166. Biology Letters [online]. 2010-04-23 [cit. 2021-03-05]. Roč. 6, čís. 2, s. 163–166. DOI 10.1098/rsbl.2009.0719. (anglicky)
- Lavinge, D. M.; Kovacs, K. M.; Bonner, W. N. „Seals and Sea lions“, s. 147–155. In MacDonald 2001.
- Riedman 1990, s. 61.
- Riedman 1990, s. 94–95.
- Riedman 1990, s. 96.
- Riedman 1990, s. 99.
- Forcada, J. „Distribution“, s. 316–321. In Perrin, Würsig a Thewissen 2009.
- LOVE, Cassandra. Photo Ark: Northern Elephant Seal [online]. National Geographic, 2018-12-15 [cit. 2021-03-06]. Dostupné online. (anglicky)
- Riedman 1990, s. 256–257.
- Riedman 1990, s. 172–175.
- BLIX, Arnoldus Schytte. Adaptations to deep and prolonged diving in phocid seals. The Journal of Experimental Biology [online]. 2018-06-15 [cit. 2021-03-06]. Roč. 221, čís. 12. DOI 10.1242/jeb.182972. (anglicky)
- COSTA, D. P.; HUCKSTADT, L. A.; CROCKER, D. E.; MCDONALD, B. I.; GOEBEL, M. E.; FEDAK, M. A. Approaches to Studying Climatic Change and its Role on the Habitat Selection of Antarctic Pinnipeds. S. 1018–1030. Integrative and Comparative Biology [online]. 2010-12-01 [cit. 2021-03-06]. Roč. 50, čís. 6, s. 1018–1030. DOI 10.1093/icb/icq054. (anglicky)
- HINDELL, Mark A.; SLIP, David J.; BURTON, Harry R.; BRYDEN, Michael M. Physiological implications of continuous, prolonged, and deep dives of the southern elephant seal ( Mirounga leonina ). S. 370–379. Canadian Journal of Zoology [online]. 1992-02-01 [cit. 2021-03-06]. Roč. 70, čís. 2, s. 370–379. DOI 10.1139/z92-055. (anglicky)
- Berta 2012, s. 70.
- Berta 2012, s. 78.
- Riedman 1990, s. 242.
- Riedman 1990, s. 234–237.
- Riedman 1990, s. 144–145.
- Riedman 1990, s. 166–168.
- ROFFE, Thomas J.; MATE, Bruce R. Abundances and Feeding Habits of Pinnipeds in the Rogue River, Oregon. S. 1262. The Journal of Wildlife Management [online]. 1984-10 [cit. 2021-03-06]. Roč. 48, čís. 4, s. 1262. DOI 10.2307/3801787. (anglicky)
- Riedman 1990, s. 153.
- Anděra 2003, s. 81.
- Riedman 1990, s. 155.
- MOORE, Peter J.; CHARTERIS, Matt; LARSEN, Eric J. Notes on New Zealand mammals 8. Predation on nesting southern royal albatrosses Diomedea epomophora by a New Zealand sea lion Phocarctos hookeri. S. 201–204. New Zealand Journal of Zoology [online]. 2008-01-01 [cit. 2021-03-07]. Roč. 35, čís. 3, s. 201–204. DOI 10.1080/03014220809510115. (anglicky)
- NORDØY, Erling Sverre. Gastroliths in the harp seal Phoca Groenlandica. S. 335–338. Polar Research [online]. 1995-01-12 [cit. 2021-03-06]. Roč. 14, čís. 3, s. 335–338. DOI 10.3402/polar.v14i3.6673. (anglicky)
- Weller, D. W. „Predation on marine mammals“, s. 923–931. In Perrin, Würsig a Thewissen 2009.
- VAN DEN HOFF, John; MORRICE, Margaret G. Sleeper shark (Somniosus antarcticus) and other bite wounds observed on southern elephant seals (Mirounga leonina) at Macquarie Island. S. 239–247. Marine Mammal Science [online]. 2008-01 [cit. 2021-02-13]. Roč. 24, čís. 1, s. 239–247. DOI 10.1111/j.1748-7692.2007.00181.x. (anglicky)
- DEMASTER, Douglas P.; STIRLING, Ian. Ursus maritimus. S. 1. Mammalian Species [online]. 1981-05-08 [cit. 2021-03-07]. Čís. 145, s. 1. DOI 10.2307/3503828. (anglicky)
- Do Polar Bears Eat Walruses? - Polar Bear-Walrus Relationship [online]. PolarBearFacts.net, 2018-01-04. Dostupné online. (anglicky)
- STANDER, P. E. Lions (Panthera leo) specialising on a marine diet in the Skeleton Coast Park, Namibia. S. A–10. Namibian Journal of Environment [online]. 2019-01-10 [cit. 2021-05-28]. Roč. 3, s. A–10. Dostupné online. ISSN 2026-8327. (anglicky)
- Riedman 1990, s. 218.
- Riedman 1990, s. 138.
- Nesting of the extremely endangered Saimaa ringed seal was saved with man-made snow banks [online]. WWF Suomi, 2017-06-13. Dostupné online. (anglicky)
- OVSYANIKOV, Nikita. Polar bears. Stillwater: Voyageur Press, 1998. Dostupné online. ISBN 978-0-89658-358-0. S. 23. (anglicky)
- SINIFF, D. B.; BENGTSON, J. L. Observations and Hypotheses Concerning the Interactions among Crabeater Seals, Leopard Seals, and Killer Whales. S. 414–416. Journal of Mammalogy [online]. 1977-08-20 [cit. 2021-03-07]. Roč. 58, čís. 3, s. 414–416. DOI 10.2307/1379341. (anglicky)
- Riedman 1990, s. 176.
- CAPPOZZO, H. L. Marine mammals : biology and conservation. New York: Kluwer Academic/Plenum Publishers ISBN 978-0-306-46573-4. Kapitola New perspectives on the behavioural ecology of pinnipeds, s. 243. (anglicky)
- Riedman 1990, s. 178–179.
- LE BOEUF, B. J.; CROCKER, D. E.; COSTA, D. P.; BLACKWELL, S. B.; WEBB, P. M.; HOUSER, D. S. FORAGING ECOLOGY OF NORTHERN ELEPHANT SEALS. S. 353–382. Ecological Monographs [online]. 2000-06 [cit. 2021-03-09]. Roč. 70, čís. 3, s. 353–382. DOI 10.1890/0012-9615(2000)070[0353:feones2.0.co;2]. (anglicky)
- Riedman 1990, s. 184–188.
- Riedman 1990, s. 212.
- Riedman 1990, s. 196.
- Mesnick, S. L.; Ralls, K. „Mating systems“, s. 712–718. In Perrin, Würsig a Thewissen 2009.
- SANDEGREN, Finn Eskil. Breeding And Maternal Behavior Of The Steller Sea Lion (Eumetopias Jubata) In Alaska. , 1970. . University of Alaska Fairbanks. . Dostupné online.
- Odell, D. K. „The Fight to Mate: Breeding strategy of California sea lions“, s. 172–173. In MacDonald 2001.
- Campagna, C. „Aggressive behavior, intraspecific“, s. 18–22. In Perrin, Würsig and Thewissen 2009.
- Dubzinski, K. M.; Thomas, J. A.; Gregg, J. D. „Communication in marine mammals“, s. 260–268. In Perrin, Würsig a Thewissen 2009.
- Riedman 1990, s. 212–215.
- SOTO, Karim H.; TRITES, Andrew W. South American sea lions in Peru have a lek-like mating system. S. 306–333. Marine Mammal Science [online]. 2011-04 [cit. 2021-03-10]. Roč. 27, čís. 2, s. 306–333. DOI 10.1111/j.1748-7692.2010.00405.x. (anglicky)
- BONESS, Daryl J.; BOWEN, W. Don; BUHLEIER, Birgit M.; MARSHALL, Gregory J. Mating tactics and mating system of an aquatic-mating pinniped: the harbor seal, Phoca vitulina. S. 119–130. Behavioral Ecology and Sociobiology [online]. 2006-10-04. Roč. 61, čís. 1, s. 119–130. DOI 10.1007/s00265-006-0242-9. (anglicky)
- FABIANI, Anna; GALIMBERTI, Filippo; SANVITO, Simona; HOELZEL, A. Rus. Extreme polygyny among southern elephant seals on Sea Lion Island, Falkland Islands. S. 961–969. Behavioral Ecology [online]. 2004-11 [cit. 2021-03-10]. Roč. 15, čís. 6, s. 961–969. DOI 10.1093/beheco/arh112. (anglicky)
- LIDGARD, Damian C.; BONESS, Daryl J.; BOWEN, W. Don; MCMILLAN, Jim I. State-dependent male mating tactics in the grey seal: the importance of body size. S. 541–549. Behavioral Ecology [online]. 2005-05-01 [cit. 2021-03-10]. Roč. 16, čís. 3, s. 541–549. DOI 10.1093/beheco/ari023. (anglicky)
- BONESS, Daryl J.; BOWEN, W. Don; OFTEDAL, Olav T. Evidence of polygyny from spatial patterns of hooded seals ( Cystophora cristata ). S. 703–706. Canadian Journal of Zoology [online]. 1988-03-01 [cit. 2021-03-10]. Roč. 66, čís. 3, s. 703–706. DOI 10.1139/z88-104. (anglicky)
- Riedman 1990, s. 209–210.
- COX, Cathleen R.; LE BOEUF, Burney J. Female Incitation of Male Competition: A Mechanism in Sexual Selection. S. 317–335. The American Naturalist [online]. 1977-03 [cit. 2021-03-10]. Roč. 111, čís. 978, s. 317–335. DOI 10.1086/283163. (anglicky)
- BONESS, Daryl J.; ANDERSON, Sheila S.; COX, Cathleen R. Functions of female aggression during the pupping and mating season of grey seals, Halichoerus grypus (Fabricius). S. 2270–2278. Canadian Journal of Zoology [online]. 1982-10-01 [cit. 2021-03-10]. Roč. 60, čís. 10, s. 2270–2278. DOI 10.1139/z82-293. (anglicky)
- REITER, Joanne; PANKEN, Kathy J.; LE BOEUF, Burney J. Female competition and reproductive success in northern elephant seals. S. 670–687. Animal Behaviour [online]. 1981-08 [cit. 2021-03-10]. Roč. 29, čís. 3, s. 670–687. DOI 10.1016/S0003-3472(81)80002-4. (anglicky)
- Berta 2012, s. 76–78.
- Walrus [online]. Seal Conservation Society [cit. 2021-03-11]. Dostupné online. (anglicky)
- Riedman 1990, s. 222–224.
- VILLEGAS-AMTMANN, S.; ATKINSON, S.; COSTA, D. P. Low Synchrony in the Breeding Cycle of Galapagos Sea Lions Revealed by Seasonal Progesterone Concentrations. S. 1232–1237. Journal of Mammalogy [online]. 2009-10-15 [cit. 2021-12-08]. Roč. 90, čís. 5, s. 1232–1237. DOI 10.1644/08-MAMM-A-319.1. (anglicky)
- Riedman 1990, s. 245.
- Riedman 1990, s. 256.
- RIEDMAN, Marianne L.; LE BOEUF, Burney J. Mother-pup separation and adoption in northern elephant seals. S. 203–215. Behavioral Ecology and Sociobiology [online]. 1982-11 [cit. 2021-03-11]. Roč. 11, čís. 3, s. 203–215. DOI 10.1007/BF00300063. (anglicky)
- Mann, J. „Parental behavior“, s. 830–835. In Perrin, Würsig and Thewissen 2009.
- KRYUKOVA, Natalia. Suckling behaviour in wild Pacific walruses (Odobenus rosmarus divergens) [online]. COLLECTION OF SCIENTIFIC PAPERS VOLUME 1 After the 9th International Conference Astrakhan, Russia 31 October – 05 November, 2016. Marine Mammal Council (Russia), 2016 [cit. 2021-03-11]. Dostupné online. (anglicky)
- RENOUF, Deane. The Behaviour of Pinnipeds. Dordrecht: Springer Netherlands, 2012. ISBN 9789401131001. S. 263. (anglicky)
- CASSINI, M. H. The evolution of reproductive systems in pinnipeds. S. 612–616. Behavioral Ecology [online]. 1999-09-01 [cit. 2021-03-12]. Roč. 10, čís. 5, s. 612–616. DOI 10.1093/beheco/10.5.612. (anglicky)
- Riedman 1990, s. 290.
- NOWAK, Ronald M. Walker's marine mammals of the world. Baltimore, Md.: Johns Hopkins University Press, 2003. Dostupné online. ISBN 978-0-8018-7343-0. S. 80-83. (anglicky)
- CAPPOZZO, Humberto L.; LE BOEUF, Burney J.; CAMPAGNA, Claudio. Pup Abduction and Infanticide in Southern Sea Lions. S. 44–60. Behaviour [online]. 1988 [cit. 2021-03-12]. Roč. 107, čís. 1–2, s. 44–60. DOI 10.1163/156853988X00188. (anglicky)
- CZIKO, Paul A.; MUNGER, Lisa M.; SANTOS, Nicholas R.; TERHUNE, John M. Weddell seals produce ultrasonic vocalizations. S. 3784–3796. The Journal of the Acoustical Society of America [online]. 2020-12 [cit. 2021-03-15]. Roč. 148, čís. 6, s. 3784–3796. DOI 10.1121/10.0002867. (anglicky)
- Riedman 1990, s. 335.
- Riedman 1990, s. 327–330.
- SANVITO, Simona; GALIMBERTI, Filippo; MILLER, Edward H. Having a big nose: structure, ontogeny, and function of the elephant seal proboscis. S. 207–220. Canadian Journal of Zoology [online]. 2007-02 [cit. 2021-03-11]. Roč. 85, čís. 2, s. 207–220. DOI 10.1139/z06-193. (anglicky)
- E.T. the Walrus practices his vocalizations at Point Defiance Zoo & Aquarium [online]. PtDefianceZoo1 [cit. 2021-03-11]. Dostupné online. (anglicky)
- TERHUNE, J. M.; HEALEY, S. R.; BURTON, H. R. Easily Measured Call Attributes Can Detect Vocal Differences Between Weddell Seals from Two Areas. S. 211–222. Bioacoustics [online]. 2001-01-01 [cit. 2021-03-11]. Roč. 11, čís. 3, s. 211–222. DOI 10.1080/09524622.2001.9753463. (anglicky)
- Thomas, J. A.; Terhune, J. „Weddell seal Leptonychotes weddellii“, s. 1217–1219. In Perrin, Würsig a Thewissen 2009.
- Rarely-heard Weddell Seal Sounds in Antarctica [online]. Henkjoost [cit. 2021-03-11]. Dostupné online. (anglicky)
- VAN OPZEELAND, I; VAN PARIJS, S; BORNEMANN, H; FRICKENHAUS, S; KINDERMANN, L; KLINCK, H; PLÖTZ, J. Acoustic ecology of Antarctic pinnipeds. S. 267–291. Marine Ecology Progress Series [online]. 2010-09-13 [cit. 2021-03-11]. Roč. 414, s. 267–291. DOI 10.3354/meps08683. (anglicky)
- Riedman 1990, s. 325–326.
- Riedman 1990, s. 332.
- Riedman 1990, s. 328–330.
- PETERSON, Richard S.; BARTHOLOMEW, George A. Airborne vocal communication in the california sea lion, Zalophus californianus. S. 17–24. Animal Behaviour [online]. 1969-02 [cit. 2021-03-11]. Roč. 17, s. 17–24. DOI 10.1016/0003-3472(69)90108-0. (anglicky)
- STEWART, Brent S.; HUBER, Harriet R. Mirounga angustirostris. S. 1–10. Mammalian Species [online]. 1993-11-15 [cit. 2021-03-11]. Čís. 449, s. 1–10. DOI 10.2307/3504174. (anglicky)
- SCHUSTERMAN, Ronald J.; KASTAK, David. A California Sea Lion (Zalophus Californianus) is Capable of Forming Equivalence Relations. S. 823–839. The Psychological Record [online]. 1993-10 [cit. 2021-03-11]. Roč. 43, čís. 4, s. 823–839. Dostupné online. DOI 10.1007/BF03395915. (anglicky)
- GISINER, Robert; SCHUSTERMAN, Ronald J. Sequence, syntax, and semantics: Responses of a language-trained sea lion (Zalophus californianus) to novel sign combinations.. S. 78–91. Journal of Comparative Psychology [online]. 1992-03 [cit. 2021-03-11]. Roč. 106, čís. 1, s. 78–91. DOI 10.1037/0735-7036.106.1.78. (anglicky)
- COOK, Peter; ROUSE, Andrew; WILSON, Margaret; REICHMUTH, Colleen. A California sea lion (Zalophus californianus) can keep the beat: Motor entrainment to rhythmic auditory stimuli in a non vocal mimic.. S. 412–427. Journal of Comparative Psychology [online]. 2013 [cit. 2021-03-11]. Roč. 127, čís. 4, s. 412–427. DOI 10.1037/a0032345. (anglicky)
- RALLS, Katherine; FIORELLI, Patricia; GISH, Sheri. Vocalizations and vocal mimicry in captive harbor seals, Phoca vitulina. S. 1050–1056. Canadian Journal of Zoology [online]. 1985-05-01 [cit. 2021-03-11]. Roč. 63, čís. 5, s. 1050–1056. DOI 10.1139/z85-157. (anglicky)
- RAVIGNANI, Andrea; FITCH, W. Tecumseh; HANKE, Frederike D.; HEINRICH, Tamara; HURGITSCH, Bettina; KOTZ, Sonja A.; SCHARFF, Constance. What Pinnipeds Have to Say about Human Speech, Music, and the Evolution of Rhythm. Frontiers in Neuroscience [online]. 2016-06-20 [cit. 2021-03-11]. Roč. 10. DOI 10.3389/fnins.2016.00274. (anglicky)
- Riedman 1990, s. 331.
- Bimbo the sea lion balancing a ball, Blackpool Zoo [online]. Blackpool Zoo, 2009-07-13 [cit. 2021-03-11]. Dostupné online. (anglicky)
- SMEELE, Simeon Quirinus; ANDERSON HANSEN, Kirstin; ORTIZ, Sara Torres; JOHANSSON, Fredrik; KRISTENSEN, Jakob Højer; LARSSON, Josefin; SIEBERT, Ursula. Memory for own behaviour in pinnipeds. S. 947–958. Animal Cognition [online]. 2019-11 [cit. 2021-03-11]. Roč. 22, čís. 6, s. 947–958. DOI 10.1007/s10071-019-01286-x. (anglicky)
- MARSHALL, Michael. Seals remember what they just did – but only for about 18 seconds [online]. NewScientis, 2019-07-03 [cit. 2021-03-11]. Dostupné online. (anglicky)
- JØN, A. Asbjørn. Dugongs and Mermaids, Selkies and Seals. S. 94–98. Australian Folklore [online]. 1998 [cit. 2021-03-12]. Roč. 13, s. 94–98. Dostupné online. (anglicky)
- Riedman 1990, s. xxi.
- ROSEN, Brenda. The mythical creatures bible : the definitive guide to legendary beings. New York: Sterling, 2009. ISBN 978-1-4027-6536-0. S. 131. (anglicky)
- PBS - The Voyage of the Odyssey - Track the Voyage - MEDITERRANEAN SEA [online]. Pbs.org, 2004-07-07 [cit. 2021-03-12]. Dostupné online. (anglicky)
- BEERINJK, K. The spirit of ancient Peru : treasures from the Museo Arqueológico Rafael Larco Herrera. New York, N.Y.: Thames and Hudson, 1997. ISBN 978-0-500-01802-6. (anglicky)
- SIGVALDADÓTTIR, Sigurrós Björg. Seals as humans - ideas of anthropomorphism and disneyfication [online]. [cit. 2021-03-12]. SELASETUR WORKING PAPER NO. 0107. Dostupné online. (anglicky)
- LARSON, S. Encyclopedia of the world's zoos. Svazek 2. svazek. Chicago: Fitzroy Dearborn, 1999. ISBN 978-1-57958-174-9. S. 1148–1150. (anglicky)
- DICKENSON, Victoria. Seal. London, UK: Reaktion Books, rok. ISBN 978-1780234892. S. 59–61. (anglicky)
- ROSE, Naomi A.; PARSONS, E.C.M.; FARINAT, Richard. THE CASE AGAINST Marine Mammals in Captivity [online]. Humane Society of the United States, 2009 [cit. 2021-03-16]. Dostupné online. (anglicky)
- LEINWAND, Donna. Sea lions called to duty in Persian Gulf [online]. USATODAY.com, 2003-02-16 [cit. 2021-03-16]. Dostupné online. (anglicky)
- The Real Navy Seals – and Sea Lions and Dolphins and Whales [online]. A.B.C., 2011-05-27 [cit. 2021-03-16]. Dostupné online. (anglicky)
- Frequently Asked Questions [online]. U.S. Navy Marine Mammal Program [cit. 2013-06-19]. Dostupné online. (anglicky)
- RONALD, K.; LAVIGNE, David M. Sealing [online]. The Canadian Encyclopedia, 2006, rev. 2015 [cit. 2021-05-28]. Dostupné online. (anglicky)
- ROBERTS, Glyn. Archivovaná kopie [online]. Tasmania: Parks and Wildlife Service, 1999 [cit. 2021-05-28]. Dostupné v archivu pořízeném z originálu dne 2018-03-17. (anglicky)
- BRUNBORG, Linn Anne; JULSHAMN, Kåre; NORTVEDT, Ragnar; FRØYLAND, Livar. Nutritional composition of blubber and meat of hooded seal (Cystophora cristata) and harp seal (Phagophilus groenlandicus) from Greenland. S. 524–531. Food Chemistry [online]. 2006-06 [cit. 2021-05-28]. Roč. 96, čís. 4, s. 524–531. DOI 10.1016/j.foodchem.2005.03.005. (anglicky)
- Reeves, R. „Hunting of marine mammals“, s. 585–588 . In Perrin, Würsig a Thewissen 2009.
- MEGGS, Jm; TAYLOR, Rj. Distribution and conservation status of the Mt Mangana stag beetle, Lissotes menalcas (Coleoptera: Lucanidae). S. 23–28. Papers and Proceedings of the Royal Society of Tasmania [online]. 1999 [cit. 2021-02-14]. S. 23–28. Dostupné online. DOI 10.26749/rstpp.133.1.51. (anglicky)
- Riedman 1990, s. 111–115.
- The rise and fall of sealing [online]. New Zealand Ministry for Culture and Heritage Te Manatu [cit. 2021-05-28]. Dostupné online. (anglicky)
- Weddell Seal [online]. Seal Conservation Society [cit. 2021-05-28]. Dostupné online. (anglicky)
- Sealing [online]. South Georgia Heritage Trust [cit. 2021-05-29]. Dostupné online. (anglicky)
- Merrick, Richard; Silber, Gregorz K.; Demaster, Douglas P.; Reynolds, John E. „Endangered Species and Populations“, s. 368–374. In Perrin, Würsig a Thewissen 2009.
- JOHNSON, William M.; KARAMANLIDIS, Alexandros A.; DENDRINOS, Panagiotis; DE LARRINOA, Pablo Fernández; GAZO, Manel; LUIS MARIANO, González; GÜÇLÜSOY, Harun. Mediterranean Monk Seal Fact Files: Overview [online]. [cit. 2021-03-17]. Dostupné online. (anglicky)
- NORONHA, C. Canada's harp seal hunt kicks off [online]. NBCnews.com, 2010-04-04 [cit. 2021-03-17]. Dostupné online. (anglicky)
- GILLIES, Rob. Canada Seal Hunt Begins Amid Controversy [online]. Huffington Post, 2009-03-23 [cit. 2021-03-17]. Dostupné online. (anglicky)
- Duffield, D. A. „Extinction, specific“, s. 402–404. In Perrin, Würsig a Thewissen 2009.
- LOWRY, L. Zalophus japonicus [online]. The IUCN Red List of Threatened Species 2017: e.T41667A113089431, 2014-10-26 [cit. 2021-03-17]. DOI 10.2305/IUCN.UK.2017-1.RLTS.T41667A113089431.en.. (anglicky)
- LAIDRE, Kristin L.; STIRLING, Ian; LOWRY, Lloyd F.; WIIG, Øystein; HEIDE-JØRGENSEN, Mads Peter; FERGUSON, Steven H. QUANTIFYING THE SENSITIVITY OF ARCTIC MARINE MAMMALS TO CLIMATE-INDUCED HABITAT CHANGE. S. S97–S125. Ecological Applications [online]. 2008-03 [cit. 2021-03-17]. Roč. 18, čís. sp2, s. S97–S125. DOI 10.1890/06-0546.1. (anglicky)
- FRENCH, Catherine. Sea Lions Take Over Ventura Docks [online]. the Log.com, 2013-04-10 [cit. 2021-03-18]. Dostupné online. (anglicky)
- BRUSCAS, Angelo. Shocking new idea for sea lion control [online]. The Daily World, 2012-07-27 [cit. 2021-03-18]. Dostupné online. (anglicky)
- SHEETS, Bill; WRITER, Herald. As sea lion populations grow, conflicts increase [online]. HeraldNet [cit. 2021-03-18]. Dostupné online. (anglicky)
- Endangered Salmon Predation Prevention Act [online]. Northwest Regional Office, 2007 [cit. 2021-03-18]. Dostupné online. (anglicky)
- Endangered Salmon Predation Prevention Act [online]. House of Representatives, 2016-11-17 [cit. 2021-03-18]. Dostupné online. (anglicky)
- HARRISON, Jon. Freshwater Slaughter [online]. Northwest Power and Conservation Council [cit. 2021-05-27]. Dostupné online. (anglicky)
- Lethal Removal of Sea Lions Necessary to Ensure Survival of Endangered Salmon Populations in Pacific Northwest [online]. Doc Hastings, 2011-06-14 [cit. 2021-03-18]. Dostupné online. (anglicky)
- BROWN, Katie. West Coast fishermen have few options against sea lions [online]. High Country News, 2019-08-22 [cit. 2021-05-27]. Dostupné online. (anglicky)
- HAIGHT, Abby. Sea lion death warrants [online]. KATU.com - Portland News, Sports, Traffic Weather and Breaking News - Portland, Oregon, 2013-05-10 [cit. 2021-03-18]. Dostupné online. (anglicky)
- Columbia River Sea Lion Management [online]. Columbia River Sea Lions: Restoring balance between predators and salmon | Washington Department of Fish & Wildlife, 2017-10-21 [cit. 2021-03-18]. Dostupné online. (anglicky)
- LAVIGNE, D. Marine mammals : fisheries, tourism and management issues. In: GALES, N.; HINDELL, M.; KIRKWOOD, R. Collingwood, Vic.: Csiro Publishing ISBN 978-0-643-06953-4. (anglicky)
- Liberal MP campaigns to shoot 'booming' fur seal population in SA [online]. ABC Radio Adelaide, 2015-06-30 [cit. 2021-03-18]. Dostupné online. (anglicky)
- METCHALFE, C. Persistent organic pollutants in the marine food chain - United Nations University [online]. United Nations University, 2012-08-16 [cit. 2021-03-18]. Dostupné online. (anglicky)
- RICHARDSON, W. John; MALME, Charles I.; GREENE, JR., Charles R.; THOMSON, Denis H. Marine mammals and noise. San Diego: Academic Press, 1995. ISBN 978-0-08-057303-8. (anglicky)
Literatura
- česky
- ANDĚRA, Miloš, 2003. Savci. (2), Šelmy, luskouni, hrabáči, hlodavci. Praha: Albatros. ISBN 80-00-00677-4.
- anglicky
- BERTA, A., 2012. Return to the Sea: The life and evolutionary times of marine mammals. Berkeley, California, USA: University of California Press. ISBN 978-0-520-27057-2. (anglicky)
- BERTA, A.; SUMICH, J. L.; KOVACS, K. K., 2006. Marine Mammals. Amsterdam: Academic Press. ISBN 978-0-12-369499-7. (anglicky)
- GILI, Claudia; MEIJER, Gerard; LACAVE, Geraldine. EAZA and EAAM Best Practice Guidelines for Otariidae and Phocidae (Pinnipeds) [online]. 2018 [cit. 2021-01-20]. Dostupné online. (anglicky)
- KIRKWOOD, Roger, 2013. Fur seals and sea lions. Collingwood, VIC: CSIRO Pub. ISBN 9780643109841. (anglicky)
- MACDONALD, David W. (eds.), 2001. The Encyclopedia of Mammals. New York: Oxford University Press. ISBN 978-0-7607-1969-5. (anglicky)
- PERRIN, W.F.; WÜRSIG, Bernd G.; THEWISSEN, J. G. M., 2009. Encyclopedia of marine mammals. Amsterdam: Elsevier/Academic Press. ISBN 0-12-373553-X. (anglicky)
- RIEDMAN, M., 1990. The Pinnipeds: Seals, sea lions, and walruses. California: University of California Press. Dostupné online. ISBN 978-0-520-06497-3. (anglicky)
- SCHEFFER, V. B., 1958. Seals, Sea Lions, and Walruses: A Review of the Pinnipedia. Standford: Stanford University Press. ISBN 978-0-8047-0544-8. (anglicky)
Externí odkazy
 Obrázky, zvuky či videa k tématu ploutvonožci na Wikimedia Commons
Obrázky, zvuky či videa k tématu ploutvonožci na Wikimedia Commons  Taxon Pinnipedia ve Wikidruzích
Taxon Pinnipedia ve Wikidruzích Slovníkové heslo ploutvonožec ve Wikislovníku
Slovníkové heslo ploutvonožec ve Wikislovníku Encyklopedické heslo Ploutvonožci v Ottově slovníku naučném ve Wikizdrojích
Encyklopedické heslo Ploutvonožci v Ottově slovníku naučném ve Wikizdrojích- (anglicky) Seal Conservation Society
- (anglicky) Pinniped Lab | University of California Santa Cruz