Migrace ptáků
Migrace ptáků je sezónní tah ptactva mezi jejich hnízdišti a zimovišti. Na migrační cestu se pravidelně vydává méně než 20 % z více než 10 tisíc ptačích druhů. Opeřenci vykonávající migraci se označují jako tažní či stěhovaví ptáci. Důvodem migrace je hlavně dostupnost potravy, obecněji vytěžení dvou či více biotopů, jejichž vhodnost pro život ptáků se sezónně proměňuje. Migrace je v ptácích geneticky zafixována, sociální učení hraje druhotnou, i když někdy poměrně významnou roli (např. velké sociální druhy jako čápi a dravci získávají detaily o migrační trase učením). Většina ptáků migruje v noci, což s sebou nese řadu výhod jako je vyhnutí se predátorům nebo nižší teploty a tedy i menší riziko dehydratace.

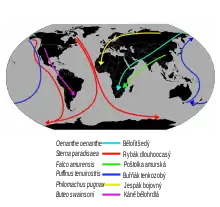
K migraci dochází podél tahových cest, které nejčastěji směřují ze severu na jih s různě velkými odchylkami k západu či východu. Tahové cesty většinou vedou podél horských pásem, vodních toků a dalších výrazných topografických linií. Vodní ptáci táhnou podél řek a pobřeží, plachtící druhy (např. dravci) vyhledávají pevninské oblasti bohaté na termické proudy, které jim usnadňují let a pomáhají tak snížit vysoké energetické nároky tahu. Vedle topografie se na vzniku tahových cest podílely i glaciální cykly nebo historické rozšíření ptáků. Stěhovavé ptáky lze rozlišit podle délky tahu na migranty na dlouhou a krátkou vzdálenost. Migranti na dlouhou vzdálenost typicky překračují velkou geografickou překážku, podnebné pásmo nebo kontinent (např. břehouši, rybáci) a často migrují i tisíce, v extrémních případech i desetitisíce kilometrů daleko. Migranti na krátkou vzdálenost migrují spíše lokálně (např. kavka obecná, která migruje ze střední do jihozápadní Evropy). Řada ptáků vykonává výškovou migraci, tedy přesun mezi různými nadmořskými výškami (typicky z výše položených hnízdišť do níže položených zimovišť). Někdy nemigrují všichni jedinci, ale pouze část populace druhu – např. jen určitý poddruh či jen dospělí jedinci. Takovéto případy se označují jako částečná migrace.
Načasování migrace se řídí geneticky fixovaným endogenním (vnitřním) cirkanuálním rytmem seřízeným podle délky světla během dne. Vedle toho mají na načasování vliv i genetická výbava, počasí, sociální stimuly, biologická zdatnost, teplota nebo dostupnost potravy. Ptáci pro navigaci používají magnetické pole Země, olfaktorické (čichové) vnímání, vizuální prvky v terénu i polohu Slunce a hvězd. Občas se stane, že se ptáci zatoulají, což je tradičně připisováno na vrub navigačním chybám jedinců, nicméně novější bádání ukazuje, že zbloudění zároveň umožňuje ptákům objevovat nová hnízdiště a zimoviště.
U stěhovavých ptáků se vyvinula řada fyziologických, hormonálních i neurologických adaptací, které jim pomáhají vyrovnat se s extrémní povahou migrace. K těmto adaptacím patří schopnost těla navýšit zásoby tuku před migrací, kontrola spánku, smršťování orgánů před tahem nebo načasování přepeřování. Se současným globálním oteplováním probíhá u stěhovavých ptáků řada velkých změn; mj. dochází k časnějším odletům do zimovišť. Ochrana tažných druhů je organizačně náročná a vyžaduje koordinaci mnoha států, pročež byla vytvořena řada úmluv jako je tzv. Bonnská úmluva, v rámci které byla sjednána i Dohoda o ochraně africko-euroasijských stěhovavých vodních ptáků.
Migrace ptáků fascinovala lidstvo od nepaměti. O tahu ptáků se objevují zmínky už ve Starém zákoně či Homérově Illiadě. Tažné ptactvo studoval i starořecký klasik Aristotelés, který vyslovil myšlenku, že vlaštovky zimují v bahně močálů. Tato hypotéza se stala převládajícím vysvětlením každoročního mizení vlaštovek po 2 tisíce let a průlom nastal až ve druhé polovině 19. století s nástupem moderních vědeckých pokusů a metod, především kroužkování. To je dodnes nejčastější metodou studia ptačí migrace. Dalším milníkem ve studiu tažných ptáků nastal s nástupem telemetrických zařízení, zejména ultralehkých geolokátorů (GLS). V polovině 10. let 21. století došlo i k prvním testům telemetrických zařízení o váze kolem 1 g, která lze připevnit i na malé druhy pěvců. I přes poměrně intenzivní zájem vědců a použití moderních technologií zůstává řada otázek okolo migrace ptáků neobjasněna.
Charakteristika migrace
Z hlediska pohybu lze ptáky rozdělit na:
- stálé (ptáci, kteří žijí ve stejné oblasti po celý rok);
- potulné (některé populace ptáků jako jsou některé sýkory či pěnkavy, které se mimo hnízdní období „potulují“ po okolní krajině bez specifického řádu);
- tažné / stěhovavé (ptáci u nichž dochází k migraci).[1]
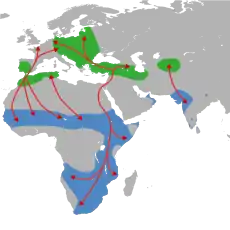
Z celkového počtu více než 10 tisíc druhů ptáků se za tažné považuje kolem 1850 druhů, čili necelých 20 %.[2][3] Migraci ptáků lze definovat jako každoroční hromadný přesun ptáků mezi jejich hnízdišti a zimovišti.[4] Termín „zimoviště“ nicméně může být problematický, a to zvláště u druhů překračujících rovník, takže se někdy místo výrazů „hnízdiště“ a „zimoviště“ používají termíny „hnízdící“ a „nehnízdící“ oblast.[5] Primárním důvodem migrace je dostupnost potravy, resp. vytěžení dvou či více biotopů, jejichž vhodnost se pro život ptáků sezónně proměňuje.[6] K těmto proměnám patří vedle změn v dostupnosti potravy i změny habitatu, případně počasí.[7] Počasí, resp. pokles teplot, může být pro některé druhy druhotný motivující faktor migrace, nicméně řada druhů jako např. někteří kolibříci dokáží přečkat tuhé zimy v hnízdištích, pokud mají dostatek potravy.[4] Navzdory dlouhodobého zájmu vědců je migrační chování ptáků z velké části stále nedostatečně vysvětleno.[8]
Migrace ptáků se odehrává převážně na severní polokouli.[3] Důvodem je to, že většina pevniny severní polokoule se nachází v mírných a subpolárních zeměpisných šířkách, ve kterých dochází v zimě k nedostatku potravy, což způsobuje migraci ptáků na jih, někdy až na jižní polokouli. Poněkud odlišná je situace u mořských ptáků, u kterých je větší pravděpodobnost migrace na jižní polokouli. Důvody jsou spojovány s většími plochami oceánů či větším množstvím vhodných ostrovů k hnízdění.[9]
Migrace ptáků neprobíhá pouze ve vzduchu. Většina tučňáků migruje ve vodě plaváním a v některých případech i chozením – např. tučňák císařský ujde z vody do kolonie na ledu i přes 100 km jedním směrem.[10][11] Podobně pochoduje i tetřívek douglaskový, který vykonává každoroční výškovou migraci o několika sto výškových metrech převážně pochodem po zemi.[12] Pravidelný migrační pochod vykonává i západní populace australského běžce emua hnědého.[13]
Migrační let

K migraci typicky dochází ve vlnách, tzn. v rámci určitého druhu nemigrují všichni ptáci najednou, ale ve skupinách. Jejich doba odletu závisí mj. na času, místních podmínkách a situaci v místě příletu.[14] Je poměrně běžné, že různé věkové skupiny nebo rozdílná pohlaví mají různé migrační vzorce. Např. samice slavíků modráčků ve východní Fennoskandinávii[15] či samice jespáků obecných se vydávají na migrační cestu dříve než samci.[16] U řady dalších druhů jako jsou vodní ptáci vodouš bahenní, jespák křivozobý, jespák rezavý nebo jespák šedý migrují dospělci dříve než juvenilní jedinci.[17] Sýkorky rodů Cyanistes a Parus migrují pouze v prvním roce života.[18] Obecně platí, že jarní migrace bývá rychlejší než podzimní.[19]
Zatímco některé druhy migrují samostatně (např. kolibříci[20] a noční migranti), jiní ptáci migrují v hejnech. Shromažďování ptáků do hejn se nazývá houfování. Tah v hejnech má řadu výhod jako kolektivní rozhodování („víc hlav víc ví“), díky kterému ptáci mohou snadněji nalézt vhodná místa k odpočinku, efektivněji v nich sbírat potravu, vybrat nejlepší tahovou cestu nebo lépe a dříve odhalit predátory. Zatímco v některých hejnech se ptáci drží daleko od sebe (např. někteří pěvci udržují vzájemnou vzdálenost i 50 m a více), jiné druhy se drží tak těsně u sebe, že se takřka dotýkají konečky křídel (např. husy nebo labutě).[21] Cestování v hejnech pomáhá snižovat energetické nároky letu.[22] Velikost hejn se může razantně lišit podle druhu od několika jedinců po tisíce ptáků. Hejna špačků mohou zahrnovat i miliony kusů.[23][24]
Některá hejna větších druhů ptáků jako jsou pelikáni, jeřábi, kormoráni, husy nebo labutě tvoří v letu formaci ve tvaru písmene V. Tato formace umožňuje jedincům po stranách snížit odpor vzduchu a tedy i množství energie nutné k letu. Odhaduje se, že ptáci takto ušetří 20–30 % energie oproti samostatnému letu.[25] Jedincům v čele formace taková úspora neplyne, a proto se ptáci v čele často střídají. Ptáci dokonce dovedou synchronizovat mávání svých křídel tak, aby co nejlépe využívali vztlakových proudů a naopak se efektivně vyhnuli sestupným proudům způsobovaných máváním ptáka před sebou.[26] U jespáků rezavých bylo naměřeno, že při pohybu v hejnech létají o 5 km/h rychleji než při samostatném letu.[27] U pelikánů bílých bylo zaznamenáno, že při letu ve formaci ve tvaru V mají pomalejší srdeční tep a nemusí tolik mávat křídly jako samostatně letící jedinci.[28] Další výhodou této formace je snadné udržování vizuálního kontaktu ptáků mezi sebou.[29]

Migrační let probíhá v různých nadmořských výškách. V oblasti středoevropských nížin ptáci táhnou mezi 0,2–1,5 km nad zemí.[30] Obecně platí, že dlouhé migrační lety probíhají ve vyšších nadmořských výškách než krátké migrační lety. Vyšší nadmořské výšky sice poskytují vhodnější podmínky k letu (nižší teploty, silnější větry), avšak je energeticky náročné se do takových výšek dostat, takže když už tam ptáci vyšplhají, typicky tam zůstávají co nejdelší dobu.[31] K největším „vzdušným horolezcům“ patří husy indické, které pravidelně létají přes himálajské pohoří. Husy se běžně vyskytují ve výškách kolem 5000–6000 m n. m., avšak mohou létat i ve výškách přesahujících 7000 m n. m.[32] Výškový rozdíl 6000 m mohou překonat během cca 8 hodin.[33] Zatímco husy indické jsou do takových výšek přinuceny letět geografickou překážkou, do několikakilometrových výšek se pouští i někteří jiní ptáci, např. břehouš černoocasý. Tito ptáci se pouští do takových výšek za účelem hledání vhodných větrů a vyhnutí se horkému vzduchu v nižších polohách.[34] Druhy pohybující se pomocí plachtění (např. dravci) se většinou pohybují v nadmořských výškách do 1500 m, kde je dostatek termických proudů.[35] Moře, kde tyto vzdušné proudy chybí, překonávají plachtící druhy nad úžinami (v Evropě patří k velmi často využívaným přechodům Gibraltarský průliv[36] nebo Messinská úžina).[37]
Tahové cesty
Ptáci se na migrační cestu vydávají z různých končin, avšak během tahu se jejich pohyb postupně koncentruje do relativně ustálených tras, jimž se říká tahové cesty (někdy též obecněji migrační cesty či migrační trasy).[38][39] Tahové cesty typicky vedou podél horských pásem, oceánského pobřeží a dalších geografických linií. Vodní ptáci často migrují podél řek a vodních toků, kde mohou nalézt potravu. Dravci a další plachtící druhy vyhledávají trasy bohaté na stoupavé vzdušné proudy a termální větry, které usnadňují jejich let, a vyhýbají se otevřeným vodním plochám, kde se tyto větry nevyskytují. Je poměrně běžné, že ptáci volí při cestě do zimovišť jinou trasu než při cestě zpět.[40]
Tahové cesty jen vzácně představují tu nejkratší možnou variantu, jak se dostat z hnízdiště do zimoviště. Klíčové faktory, které ovlivňují délku cesty, jsou topografie, počasí a stanoviště bohatá na potravu.[41] Někdy ptáci následující delší trasu díky zdědění migračních genů svých předků, kteří vedli migraci po určité trase, která však již nepředstavuje tu nejvýhodnější nebo nejkratší možnost. Příkladem může být drozd západní, malý severoamerický pěvec, jehož populace byla během poslední doby ledové rozdělena do dvou geneticky výrazně odlišných skupin, z nichž jedna zůstala ve vnitrozemí a druhá byla vytlačena k aljašskému pobřeží. Během migrace však drozdi z Aljašky nejdříve letí tisíce kilometrů na východ, než zamíří na jih směrem ke svým středoamerickým hnízdištím, místo aby následovali kratší trasu podél západního pobřeží.[42] Často se stává, že sousedící populace téhož druhu migrují do zimovišť jinými trasami nebo migrují do zimovišť v jiných lokalitách. Oblasti mezi těmito populacemi se říká migrační rozhraní. Tento jev způsobují geografické bariéry i historické glaciální cykly, které formovaly výskyt a pohyb ptactva. Migrační rozhraní se vyskytují i na území Česka – zatímco některé moravské populace ptáků táhnou na jih jihovýchodní tahovou cestou, ptáci ze západní části země mohou zvolit jihozápadní migrační cestu.[43] Příkladem mohou být rákosník obecný, rehek domácí[44] nebo čáp bílý.[45]

Nejčastěji dochází k migraci ze severu na jih s menší či větší odchylkou k východu či západu. Na severní polokouli ptáci typicky odlétají na jaře do hnízdišť v severních oblastech, kde začínají kvést rostliny a množí se hmyz, a na podzim se vracejí do teplejších zimovišť na jihu. Na jižní polokouli je tento proces obráceně.[46] Vedle lepší výtěžnosti zdrojů je výhodou tahů po ose sever-jih oproti migracím na trase východ-západ lepší využití denního světla; zejména v polárních a subpolárních oblastech dlouhé letní dny poskytují ptákům více času na shánění potravy a krmení svých mláďat. Díky tomu mívají tažní ptáci větší snůšky než stálí ptáci.[47] Tahové cesty některých druhů vedou spíše z východu na západ. Příkladem mohou být některé africké druhy migrující mezi vyprahlými léty západní Afriky a vlhkými zimami na východě nebo některé druhy mořských ptáků, vrubozubých a bahňáků, kteří na jaře opouští svá zimoviště v arktických oblastech a nejdříve se vydávají východním či západním směrem podél severních hranic kontinentů, než zamíří zahnízdit do vnitrozemí.[48] Obecně platí, že tažní ptáci hnízdící na severní polokouli mají delší migrační trasy než ptáci hnízdící na jižní polokouli.[49]
Koncept tahových cest je široce využíván v managementu a ochraně ptačích populací, které vyžadují mezinárodní spolupráci. V těchto případech jsou tahové cesty značně zjednodušené, nutně ztrácí celou řadu informací. Nereflektují tahy konkrétních jedinců, ale spíše globální trendy.[5] Mezinárodní svaz ochrany přírody rozlišuje 4 hlavní tahové cesty: mezi Jižní a Severní Amerikou, Afrikou a Evropou, střední a jižní Asií, východní Asií a Australasií.[50]
Tahové zastávky

Většina stěhovavých ptáků během migrace zastavuje na vhodných stanovištích. Tato stanoviště poskytují ptákům potravu nutnou k doplnění energetických zásob, bezpečné útočiště před predátory a někdy i ochranu před extrémním počasím jako jsou tropické bouře. Tahové zastávky jsou pro ptáky často naprosto klíčové. Někteří tažní ptáci ztrácí v první části migračního letu i polovinu své váhy a bez potravních zastávek, kde dojde k opětovnému nabrání energie, by nebyli schopni tah dokončit.[51] Někteří ptáci stráví až 80 % doby migrace na odpočinkových stanovištích. Doba strávená na odpočinkovém stanovišti je ovlivněna fyzickým stavem ptáků, jejich biorytmy (resp. endogenními hodinami) a environmentálními podmínkami jako je počasí, míra predace a dostupnost potravy.[52] Tažní ptáci vyskytující se na tahových zastávkách mezi hnízdišti a zimovišti se v rámci dané lokality označují jako protahující druhy (případně průtažní druhy).[53]
Oblasti tahových zastávek jsou oblíbeným místem pro pozorovatele ptáků. V České republice patří k oblíbeným lokalitám např. Lednické rybníky, v Maďarsku národní park Hortobágy, kde lze na jaře pozorovat desetitisíce táhnoucích jeřábů, v Rakousku Neziderské jezero známé velkými hejny kachen a bahňáků.[54]
Ekologické důsledky
Migrační druhy ptáků hrají důležitou roli v místních ekosystémech. Stěhovaví ptáci pomáhají s opylováním rostlin i roznášením semen, kontrolují populaci hmyzu a slouží jako kořist místních predátorů či naopak loví místní živočichy.[55] Jikry některých druhů ryb se dokonce mohou rozptylovat do vzdálených a izolovaných vodních ploch skrze ptačí trávicí trakt.[56] Masivní přesuny ptáků z jednoho místa na druhé mohou zlepšovat odolnost místních ekosystému následkem propojení biotopů, což zvyšuje jejich druhovou rozmanitost.[57] Stěhovavé druhy však nemusí přinášet jen pozitiva; s ptačími migranty přichází i hrozba masivního šíření nemocí a virů.[58] Za příklady může sloužit rozšíření neotropických klíšťat ve Spojených státech následkem jejich introdukce na tělech ptáků[59] nebo šíření ptačí chřipky kolem světa.[60]
Migrační strategie
Existuje celá řada migračních strategií, které se nejčastěji rozdělují podle délky (migrace na krátkou vs. migrace na dlouhou vzdálenost) nebo převažující denní doby migrace (denní vs. noční). Mezi migračními strategiemi není jasně stanovená hranice. Nezřídka se stává, že různé populace či jedinci v rámci téhož druhu praktikují odlišné migrační strategie (např. část populace může migrovat na dlouhou vzdálenost, část na krátkou vzdálenost a část populace je stálá).[61][62]
Migrace na krátkou vzdálenost

Jako migrace na krátkou vzdálenost se označují tahy ptáků bez významnějších geografických překážek, typicky v rámci kontinentů nebo podnebných pásů.[63][61] Maximální vzdálenost, která by odlišovala migraci na krátkou vzdálenosti od dlouhé se většinou neuvádí nebo se liší podle zdroje. Např. americký ornitolog John H. Rappole považuje horní hranici vzdálenosti tahu migrantů na krátkou vzdálenost kolem 2 tisíc kilometrů; delší tahy považuje za migrace na dlouhou vzdálenost.[63]
Migranti na krátkou vzdálenost jsou typicky druhy z tropického, subtropického a jižních oblastí mírného podnebného pásu. Zejména v tropech mohou ptáci migrovat jen velmi krátkou vzdálenost, jelikož změny v habitatu a ročním úhrnu srážek se mohou zásadně lišit v blízkých blízkých areálech a ptákům tak stačí migrovat jen do sousední oblasti.[62] Příkladem migranta na krátkou vzdálenost je strnadec zpěvný, který se zdržuje mezi hnízdišti na hranicích Spojených států a Kanady, a zimovišti ve středních a jižních částech Spojených států.[64] Příkladem českého migranta na krátkou vzdálenost může být kavka obecná, která odlétá na zimu do jižní Francie a karpatské kotliny.[65] Migranti na krátkou vzdálenost hnízdící v Česku táhnou hlavně do západní Evropy, zatímco do Česka táhnou ptáci ze severní Evropy včetně Polska.[66] Obecně platí, že migranti na krátkou vzdálenost táhnou v menších hejnech než migranti na dlouhou vzdálenost.[22]
Migrace na dlouhou vzdálenost
Migrace na dlouhou vzdálenost představuje tah, během kterého ptáci překonávají velkou geografickou překážku, jako je moře či oceán, migrují na jiný kontinent nebo překračující více podnebných pásem.[63][61]
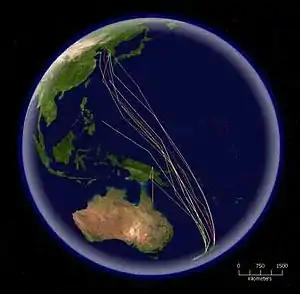
Jedním z nejtypičtějších dálkových migrantů je vlaštovka obecná, jejíž evropská populace (subsp. rustica) podniká každoroční migraci z Evropy do afrických, arabských a asijských tropů.[67] Délka trasy, kterou dálkoví migranti každoročně urazí, může jít do tisíců i desetitisíců kilometrů. Jespáci rezaví každý rok létají z hnízdišť v arktické Kanadě do zimovišť na jižním cípu Jižní Ameriky, přičemž urazí až 6500 km během jediného nepřetržitého letu.[68] Rekordmanem v nejdelším potvrzeném nepřetržitém migračním letu je však břehouš rudý (subsp. baueri), u kterého byla naměřena migrační cesta z Aljašky na Nový Zéland dlouhá 12 200 km. Let trval 11 dní a jedinec letěl rychlostí až 89 km/h. Tento výkon břehoušovi vysloužil zápis v Guinnessově knize rekordů coby „nejdelší nepřetržitá ptačí migrace“.[69]
Vůbec nejdelší vzdálenost mezi hnízdištěm a zimovištěm ze všech zvířat každoročně překonává rybák dlouhoocasý. Rybáci hnízdí v Arktidě a zimují v antarktických oblastech. Nejedná se však o nepřetržitou migraci jako u břehouše rudého; každý migrační let trvá kolem 2–3 měsíců a rybáci během něj často zastavují na odpočinkových stanovištích, aby nabrali síly. Ptáci během přesunu mezi hnízdištěm a zimovištěm nalétají kolem 25 tisíc kilometrů. Za celý rok nalétají i přes 80 000 km.[70] Podobně se na svých cestách zastavují i další migranti, přičemž občas kombinují přímé lety bez zastávky a pomalejší lety s množstvím odpočinku. Příkladem může být lejsek bělokrký, který pozvolna migruje ze střední a východní Evropy až na okraj Středozemního moře, odkud lejsci táhnou i 2–3 dny v kuse do Afriky.[71]
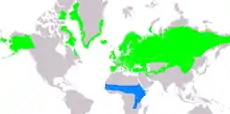
Ojedinělá je migrace bělořitů šedých ze Severní Ameriky. Bělořiti se do severoamerických končin rozšířili z Evropy někdy v pleistocénu. Stejně jako evropská populace, i ta severoamerická migruje do zimovišť v subsaharské Africe. Jedná se o jediného zpěvného ptáka, který migruje ze severní části západní polokoule do Afriky. Migrační trasa jedním směrem má kolem 14 500 km a bělořiti během ní urazí kolem 290 km denně. Zůstává otázkou, jak dokáže tento 25 g vážící, samostatně migrující pěvec úspěšně zdolat takto obrovskou vzdálenost, a to zvláště už v prvních měsících života.[72][73]
Jedněmi z největších migrantů jsou trubkonosí. Řada trubkonosých hnízdí na jižní polokouli a migruje na tu severní.[74] Trubkonosí s oblibou pro své lety využívají dostupné větry, takže jejich migrační trasy často nejsou přímočaré. Např. u buřňáků temných byla naměřena migrace dlouhá 64 tisíc kilometrů, protože buřňáci letěli krouživými a elipsovitými pohyby z Nového Zélandu přes Japonsko a Aljašku do Kalifornie a poté zpět na Nový Zéland.[75]
Výšková migrace
Zvláštním typem migrace je výšková migrace. Jedná se o přesun mezi hnízdišti ve vyšších polohách do níže položených zimovišť, kde je přes zimu dostatek potravy.[76] Odhaduje se, že výškovou migraci vykonává kolem 10 % ze všech druhů ptáků.[77] Na Havaji až 30 % druhů ptáků migruje výškově a v kontinentální Severní Americe více než 20 % druhů.[78]

Výšková migrace u ptáků nastává hlavně v tropech, kde bývá dostatek potravy po celý rok, a ptáci mohou nalézt vhodné podmínky pro zimu přesunem pouhých několika set výškových metrů. Tropické oblasti mají celoroční dostatek světla, takže ptáci nemají potřebu migrovat za delšími dny.[79][80] Typickým příkladem výškových migrantů jsou kolibříci – kolem jedné čtvrtiny druhů kolibříků vykonává výškovou migraci.[76][81]
Zatímco některé druhy vykonávají pouze výškovou migraci (krátký výškový přesun v rámci jednoho pohoří), jiné druhy vykonávají zároveň i dálkovou migraci. Extrémním příkladem jsou pěnice horské, které hnízdí v Himálajích a zimují v jižní Indii.[82] Krajním příkladem z opačného konce jsou tetřívci douglaskoví, kteří migrují v některých případech pouhých 300 m. V Guinessově knize rekordů jsou zapsáni coby vykonavatelé „nejkratší ptačí migrace“. Tito tetřívci, podobně jako některé další ptačí druhy, vykonávají tzv. převrácenou výškovou migraci, jelikož se na zimu přesouvají do vyšších poloh a na léto sestupují do níže položených oblastí.[83]
Částečná migrace
V případě tažných ptáků nemusí nutně migrovat všichni jedinci daného druhu, ale jen část populace. V takovém případě se jedná o tzv. částečnou migraci. Částečná migrace je velmi častá; např. v Austrálii lze za částečně tažné ptáky označit až 44 % pěvců a 32 % ostatních ptáků.[84] V Tasmánii až 80 % ze všech stěhovavých ptáků tvoří částeční migranti. U některých tasmánských druhů bylo zjištěno, že migrují pouze v některých letech. V části Jižní Ameriky, která zasahuje do jižní polokoule, je kolem 70 % stěhovavých ptáků pouze částečnými migranty.[85] Hlavním důvodem částečného tahu je vnitrodruhová konkurence o potravní zdroje v zimovištích, kde jsou potravní zdroje dostatečné pouze pro část populace, takže ta druhá část se vydává za potravou jinde. Zdá se, že částečná migrace nastává hlavně v oblastech se středně mírnými zimami, tzn. kde v zimě nedochází k třeskutým mrazům, avšak ani k nijak vysokým teplotám.[86]
Zvláštní typy tahu
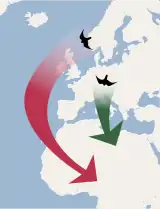
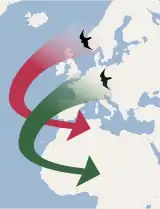
Zvláštním typem migrace je tzv. „leapfrog“ (pojem z angličtiny, kde označuje dětskou hru, během které dítě skáče přes ohnutá záda jiného dítěte). Při tomto typu migrace se určité populace druhu, které obývají vyšší zeměpisné šířky než ostatní populace, přesunou na zimu do oblastí nižších zeměpisných šířek, přičemž s každou migrací přeletí („přeškočí“) přes ostatní populace, které jsou buď stálé, nebo migrují pouze na krátkou vzdálenost.[87] Klasickým příkladem migrace typu „leapfrog“ je strnadec kaštanový ze Severní Ameriky.[88]
Vedle „leapfrog“ se rozlišuje ještě tzv. řetězová migrace (angl. chain migration), při které jižní i severní populace druhu táhnou stejným směrem a urazí zhruba stejnou vzdálenost, takže nakonec skončí ve stejném pořadí, což je zásadní rozdíl oproti „leapfrog“ migraci. Tento typ tahu nastává např. u rorýsů obecných.[89]
Noční migrace
Většina druhů ptáků migruje v noci.[90] Ostatní ptáci migrují buď ve dne, nebo ve dne i v noci.[91] K nočním migrantům patří i denní druhy, tzn. ptáci, kteří jsou jinak aktivní ve dne. Dravci a plachtící druhy se nočním letům vyhýbají, protože během chladných nocí se netvoří dostatek termických větrů. U ostatních druhů není zcela zřejmé, podle jakého klíče se rozhodli pro noční migraci. Svou roli patrně bude hrát celá řada faktorů od fyziologických po ty environmentální.[92] K nesporným výhodám noční migrace patří relativně stabilní větry, možnost navigace podle hvězd, větší množství denního světla na krmení, efektivní vyhnutí se predátorům nebo nižší teploty a vyšší vlhkost, což snižuje riziko dehydratace.[93][94]

Některé druhy predátorů se dokázaly adaptovat na noční migraci ptáků, ze které těží. K těmto predátorům patří ostříž jižní, který hnízdí ve Středozemí. U tohoto dravého ptáka dochází k pozdnímu zahnízdění (červenec–říjen), což je adaptace na hojnost potravy v tomto období. Tu pro ostříže zajišťují noční ptačí migranti (hlavně menší pěvci) mezi Evropou a Afrikou, kteří přes hnízdiště ostřížů na podzim hojně prolétají.[95][96] K dalším příkladům živočicha, který se adaptoval na noční migrace, patří netopýr obrovský. Obecně platí, že jakékoliv druhy netopýrů útočí na ptáky jen zcela výjimečně, avšak španělské populace netopýrů obecných pravidelně konzumují menší migrující ptáky, kteří tudy na jaře a na podzim prolétají. Netopýři obrovští loví zejména menší druhy migrujících pěvců (např. budníčky lesní, červenky obecné), které patrně napadají přímo za letu.[97]
Zatímco denní migranti začínají tah zrána, noční migranti vylétávají navečer. Pokud je to možné, dosedají koncem noci, nicméně např. při přeletu velkých vodních ploch jsou druhy neuzpůsobené pobytu na vodě přinuceny pokračovat v letu i za dne.[98] Hlavní nevýhodu nočního tahu představuje masivní redukce spánku, jelikož ptáci v noci táhnou a ve dne shánějí potravu. Ke krátkému hřadování může docházet v pozdním odpoledni těsně před počátkem tahu,[99] avšak někteří ptáci nespí vůbec (podrobněji viz kapitola Spánek). Krátké odpolední „schrupnutí“ nastává v době migrace i u nočních migrantů chovaných v zajetí.[90]
Noční migranti během letu vydávají častá krátká kontaktní volání. Tato volání slouží k orientaci ptáků v rámci hejna, udržování vhodných rozestupů i prevenci srážek s ostatními ptáky. Zdá se, že kontaktní volání jsou důležitá hlavně pro juvenilní jedince, kteří se během nočních tahů vokálně projevují častěji než dospělci.[100] Při nejmenším u některých druhů mají noční zvuky zakódovanou informaci o pohlaví jedince a jeho identitě.[101]
Evoluce

Migrace se u jednotlivých ptačích taxonů vyvinula dynamicky a nezávisle na sobě.[102] Migrační chování je v ptácích geneticky zafixováno a sociální učení hraje někdy sice důležitou, avšak druhotnou roli. Jedním z hlavních (negenetických) argumentů pro genetickou podporu migrace je to, že čerstvě narození ptáci odlétají z hnízdišť do svých zimovišť ještě před zhoršením životních podmínek v hnízdištích a bez podpory genů by postrádali motivaci pro tak energeticky náročnou a riskantní aktivitu jako je migrační let. Negenetické faktory přesto mohou hrát určitou roli, jejíž důležitost závisí na druhu. Například některé dlouho žijící sociální druhy jako husy, čápi, jeřábi nebo dravci migrují v menších skupinách vedených nejzkušenějšími jedinci, kteří trasu již několikrát absolvovali. Naproti tomu menší tažní ptáci jako jsou pěvci mají tahovou cestu převážně vrozenou.[102][103][104]
Pod silnými selekčními tlaky může dojít ke změně migračních genetických znaků již během několika generací. Při pokusech s uměle odchovanými pěnicemi černohlavými bylo zjištěno, že tito pěvci při selektivním křížení v zajetí opozdili odlet své podzimní migrace o více než týden v průběhu pouhých dvou generací.[105] Stejnému druhu se geneticky zafixovala i změna tahové cesty již během 30 let.[106]

K vývoji migrace došlo postupně – ptáci nejdříve hledali vhodnější podmínky pro život, načež se postupně začali vydávat dále a dále.[107][108] Tato skutečnost vzbuzuje otázku, zda se ptáci vydávali hledat vhodnější zimoviště, čímž by si zlepšili vyhlídky na přežití během zimního období, nebo se vydávali hledat vhodnější podmínky pro zahnízdění, což by zvyšovalo šance jejich reprodukční úspěšnosti. Nejnovější ornitologické výzkumy z počátků 21. století se přiklání k tomu, že ptačí migrace se vyvinula postupným objevováním nových zimovišť.[109] Dlouhodobě je totiž pro ptáky výhodnější věrnost hnízdišti než zimovišti.[109][110][111] Pokud ptáci hnízdí v oblasti s dostatkem zdrojů po celý rok, nedojde k tvorbě selekčních tlaků, následkem kterých se ptáci začnou poohlížet po nových oblastech s příhodnějšími životními, resp. potravními podmínkami.[110]
Jedna z možných příčin vývoje migračních návyků ptáků představují glaciální cykly, kterých se na Zemi vystřídalo jen za posledních 2,5 milionů let kolem 20. S rozšiřováním ledovců během každé doby ledové směrem od polárních oblastí do nižších zeměpisných šířek docházelo k postupnému vytlačování ptáků a jejich hnízdišť na jih; naopak s ústupem ledovců docházelo k odhalování nových úrodných oblastí, které se ptákům zpřístupnily.[111] S roztahováním ledovců nedošlo k pouhému zkrácení délky migračních tras – u některých druhů došlo k úplnému vynechání migrace.[112]
Fyziologie a kontrola
Migrace je spojena s rychlým životním tempem – migrační druhy oproti stálým druhům vykazují vyšší plodnost, dospívají dříve a nežijí tak dlouho. U částečně tažných ptáků je tento rozdíl o něco menší.[8]
Načasování
Život ptáků se řídí vrozeným endogenním celoročním (cirkanuálním) rytmem. Tento geneticky fixovaný rytmus seřízený podle délky světla během dne určuje přibližný čas jarní i podzimní migrace.[93][113][114] Dále mají na načasování migrace vliv i genetická výbava, počasí, sociální stimuly, biologická zdatnost, teplota nebo dostupnost potravy.[93]
Těsně před migrací ptáci začínají vykazovat fyziologický proces zvaný tahový neklid, v odborné literatuře označován německým výrazem Zugunruhe. Tahový neklid se vyznačuje zvýšenou aktivitou a nervozitou někdy až hraničící s úzkostí. U ptáků v zajetí se tento fenomén projevuje vysokou noční aktivitou, nespavostí, neustálým poskakováním a třepotáním křídel. Délka, po kterou ptáci v zajetí vykazují tahový neklid, je přibližně stejná jako délka migrace druhu ve volné přírodě.[99] V době tahového neklidu se ptáci v zajetí orientují směrem k destinaci migrační trasy populace ve volné přírodě.[113]
Orientace a navigace
Ptáci pro navigaci využívají magnetického pole Země, olfaktorické (čichové) vnímání, vizuální prvky v terénu i polohu Slunce a hvězd.[115] Vypadá to, že ptáci během navigace používají kombinaci všech těchto prvků, které mezi sebou porovnávají a vyhodnocují. Zatímco pro některé druhy může být hlavním navigačním prvkem magnetické pole Země, u jiných druhů to je čichová kognitivní mapa (např. trubkonosí).[116][117]
I přes intenzivní vědecký výzkum a řadu praktických pokusů zůstává přesný způsob navigace a orientace ptáků z velké části nevysvětlen.[117] Zejména noční migrace denních druhů vzbuzuje otázku, jakým způsobem se ptáci v noci navigují, protože jejich noční vize často nemusí být o nic lepší než noční vize člověka.[118] Podle laboratorních experimentů se noční migranti mohou navigovat pomocí magnetického pole Země, které patrně mají schopnost vidět skrze zvláštní fotoreceptory zvané kryptochromy. Tyto látky se vyskytují v sítnici některých ptáků, a patrně reagují na elektromagnetické vlnění zemského magnetického pole.[119][120]
Zatoulanci
Občas se stane, že se stěhovaví ptáci zatoulají do končin, kde se běžně nevyskytují. V takovém případě jsou označováni za zatoulance.[53] V krajních případech se mohou ocitnout i tisíce kilometrů od běžného místa výskytu.[121] Nejčastějšími důvody těchto záletů bývá nepříznivé počasí (např. silný vítr nebo snaha vyhnout se bouři) nebo dezorientace ptáků.[122] Při nejmenším někteří ptáci se dokážou s jevy počasí vyrovnat. U orlovců říčních a včelojedů lesních bylo zjištěno, že starší jedinci během letu umí vykompenzovat silné větry změněním kurzu, takže vítr neovlivní směr jejich letu tak jako mladší jedince.[123] Někteří ptáci dokážou silných větrů naopak využít ve svůj prospěch. Menší druhy ptáků ze severní Ameriky využívají severozápadní větry k podpoře letů přes západní Atlantik směrem do Jižní Ameriky.[124]
Zvláštní případ zatoulání ptáků představuje tzv. reverzní migrace (reverse migration). Tento fenomén označuje situaci, kdy ptáci migrují opačným směrem, než kterým by normálně měli letět.[121] Reverzní migrace bývají spojovány mj. s extrémními výkyvy počasí nebo geografickými bariérami, na které narazí juvenilní, prvně migrující jedinci.[125] Britští ornitologové James Gilroy a Alexander Charles Lees nicméně upozornili na to, že u ptáku sice dochází k dezorientaci nejčastěji opačným směrem, nicméně ptáci mohou zamířit i jinými směry. Tyto náhodné rozptyly ptáků jsou podle nich způsobeny genetickými abnormalitami.[126]
Dalším fenoménem migračních zatoulanců je tzv. přestřelení (overshooting), ke kterému dochází, pokud ptáci při tahu z jakéhokoliv důvodu nepřistanou v místě zimoviště či hnízdiště a pokračují v letu dále. Důvody mohou být různé, mj. se může jednat o přehlédnutí ostatních jedinců téhož druhu nebo nerozpoznání vhodného biotopu, takže migrující ptáci letí dále a dále.[126]
Zatoulaní jedinci nemusí nutně znamenat pro ptáky problém – jsou zdokumentovány případy, kdy „zatoulanost“ ptáků vedla k objevení nových tahových cest, zimovišť nebo hnízdišť.[127][128] Zatoulaní ptáci tak mohou hrát významnou roli v areálu rozšíření ptáků.[122]
Adaptační mechanismy

U stěhovavých ptáků se vyvinula řada fyziologických, hormonálních i neurologických adaptací, které jim pomáhají vyrovnat se s extrémní povahou migrace. K těmto adaptacím patří hlavně schopnost těla navýšit zásoby tuku před migrací, kontrola spánku, transformace vnitřních orgánů nebo přepeřování.
Zásobování tukem
Migrační druhy jsou uzpůsobeny jak tvorbě tukových zásob před odletem, tak obdobím půstu (např. během dlouhých migračních letů). Těsně před počátkem tahu mohou tukové zásoby narůst až na 50–60 % celkového tělesného objemu,[129] avšak výše tukových zásob závisí na délce migrační trasy i na tom, jak často nebo jestli vůbec se ptáci chystají podniknout tahové zastávky a doplnit zásoby. V době krmení před migračním letem dochází u metabolismu ptáků k hormonálním změnám, které jim umožní efektivněji nakládat s pozřeným jídlem a ukládat ho ve formě tuků.[130] Hormony (resp. hormon ghrelin, který je spojován s apetitem) patrně rozhodují za ptáky i to, zda jsou jejich zásoby tuku již dostatečné pro nadcházející let.[131]
Zásoby tuku jsou uloženy v tukových tkáních ve formě energeticky bohatých triglyceridů. Oxidací těchto mastných kyselin během letu dochází ke tvorbě a distribuci potřebné energie do svalů a cévní soustavy.[132]
Transformace vnitřních orgánů
U tažných ptáků dochází v průběhu roku ke změnám velikosti vnitřních orgánů. Před migračním letem nastává smrštění těch vnitřních orgánů, které během energeticky vyčerpávajícího letu jednoduše „nejsou potřeba“. K těm patří hlavně orgány zažívací soustavy, které se mohou smrsknout o více než 50 % (ledviny, játra, střeva).[133][134] Naopak u orgánů klíčových k letu (prsní svaly, srdce, plíce) může dojít k jejich roztažení. Orgány obnoví svou původní velikost po příletu do zimoviště nebo na odpočinkové stanoviště.[135][136][137]
Obnova orgánů na jejich původní velikost trvá i několik dní, přičemž hlavně při tahových zastávkách ptáci mohou kompenzovat zmenšené orgány trávicího ústrojí pomocí prodloužené doby zažívání.[133]

Spánek
Spánek ptáků, zvláště během dlouhých migračních letů, zůstává z velké části nevysvětlen.[93] Je potvrzeno, že celá řada ptáků dokáže spát za letu. U fregatek obecných bylo prokázáno, že dovedou spát během letu s jednou (tzv. spaní na půl mozku) nebo oběma hemisférami. Zatímco během pobytu na souši fregatky prospí kolem 12 hodin denně, během dlouhých letů spí v průměru pouze kolem 40 minut denně, aniž by to bylo poznat na jejich tělesném výkonu či pozornosti. Není úplně zřejmé, jak fregatky dokážou vystačit po dobu několika týdnů i měsíců s tak malým množstvím spánku; vysvětlení bude patrně souviset s neurochemickými procesy v mozku.[138]
.jpg.webp)
U strnadců bělopásých bylo pokusy vyzkoumáno, že strnadci dokážou během tažného období zredukovat množství svého spánku o dvě třetiny, aniž by se snížila přesnost jejich reakcí na předkládané úkoly. Naproti tomu v době mimo tažné období byla reakce strnadců na předkládané úkoly rapidně snížena již po jednom dni s redukovaným množstvím spánku.[99] Rorýsi velcí dokáží spát za letu a zůstat tak ve vzduchu po dobu nejméně půl roku.[139]
Přepeřování
U tažných druhů lze rozlišit dvě nejčastější přepeřovací strategie: k přepeřování dochází buď po období rozmnožování v oblasti hnízdišť před migračním letem, nebo (častěji) až po příletu do zimovišť. Obě strategie přitom mají své výhody: čerstvě přepeření jedinci před migračním letem mají nové peří, které pomáhá zefektivnit let; naopak post-migrační přepeření pomůže s obnovou peří, které se při tahu může značně obnosit. Nadto existuje celá řada dalších různých variací přepeřovací strategie. Někteří ptáci přepeřují hned dvakrát ročně (např. budníček větší[140]), řada druhů přepeřuje během migrace (např. rákosník velký, lesňáček lejsčí), přičemž některé z těchto druhů přepeřují jen některé letky (např. cvrčilka říční[141]).[142]
Migrace ptáků v Česku a Evropě

V Evropě žije kolem devadesáti stěhovavých druhů ptáků.[143] Kolem 68 druhů ptáků (z toho 55 pěvců), kteří hnízdí v Evropě, migrují každoročně do Afriky.[144] V Česku hnízdí desítky druhů, kteří na zimu odlétají do tzv. „teplých krajin“, naopak desítky druhů přilétají do Česka zazimovat. Zatímco některé český tažných ptáků táhnou pouze nepravidelně, částečně nebo výjimečně, jiné druhy táhnou do teplých krajin pravidelně.[145] K těm nejznámějším ptákům, kteří v Česku hnízdí, avšak pravidelně odlétají zimovat na jih, patří vlaštovka obecná, čáp bílý, drozd zpěvný, holub hřivnáč, skřivan polní, krutihlav obecný, čírka modrá, rákosník obecný, dudek chocholatý, rehek zahradní, rorýs obecný nebo rybák obecný.[44] Migrující druhy odlétají na jih nejčastěji na podzim (září, říjen) a vrací se na jaře (březen, duben).[66] K ptákům, kteří v Česku zimují, patří mj. brkoslav severní, pěnkava jikavec, čečetka zimní, káně rousná, morčáci, hoholi, turpani, hoholka lední nebo potáplice.[66]
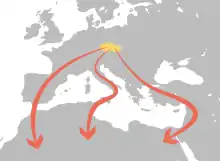
Ptáci nejčastěji odlétají do oblasti Středomoří, Pyrenejského a Apeninského poloostrova, některé druhy však odlétají dále do Afriky.[146] Ptáci hnízdící v Česku, potažmo střední Evropě, odlétají do svých zimovišť třemi hlavními tahovými cestami:
- jihozápadní cestou přes Pyrenejský poloostrov a Gibraltarský průliv;
- jihovýchodní cestou přes Balkánský poloostrov a Sýrii;
- apeninskou cestou vedoucí přes Apeninský poloostrov a Sicílii a „napřímo“ přes Středozemní moře.[147]
Zatímco první dvě cesty vedou podél pevniny a snaží se tak vyhnout otevřeným plochám, ta třetí překonává významnou geografickou bariéru – Středozemní moře. Díky relativní ustálenosti tahových cest i pravidelnosti migrace lze předvídat, kudy a kdy budou stěhovavé druhy Českem protahovat. Toho využívají odchytové stanice, které kroužkují ptáky za účelem vědeckého výzkumu a poznání migrace. Nejaktivnějším kroužkovacím stanovištěm v Česku je odchytová stanice na Červenohorském sedle, které tvoří přirozený trichtýř Jesenického masivu. Mezi lety 2010–2018 bylo na Červenohorském sedle odchyceno a okroužkováno kolem 100 jedinců 125 druhů ptáků.[148]
V Evropské unii včetně Česka jsou stěhovavé druhy chráněny mj. Bonnskou úmluvou,[149] nicméně i tak dochází v některých členských státech k odstřelům stěhovavých druhů v masových měřítkách pro „sport“. Na Maltě, kudy každoročně protahuje kolem 40 milionů ptáků, je každoročně pytláky zastřeleno kolem 2 milionů ptáků.[150] Podobně i na Kypru bylo jen v roce 2016 uloveno cca 2,3 milionu ptáků (především pěvců), kteří tvoří hlavní přísadu do místní „pochoutky“ zvané ambelopoulia.[151] Odhaduje se, že ve Středomoří každoročně hyne až 25 milionů ptáků následkem pytlačení.[152]
Ptačí migrace a člověk
Tichomořské plavby
_(14763687525).jpg.webp)
Zmínky o migraci ptáků se objevují v orálních i písemných tradicích lidstva již od pradávna. Ptačí migrace patrně hrály důležitou roli v navigaci plaveb Polynésanů a Mikronésanů, jimž již před několika tisíci lety poskytovaly indicie o existenci pevniny, což podporují mnohé legendy dochované v orálních tradicích tichomořských národů. Např. na ostrově Bougainville se traduje, že předci tamních obyvatel vydedukovali přítomnost dosud neznámého ostrova pozorováním tahu ptáků. Domorodci si vyhlédli hvězdu, která byla ve směru jejich letu, nasedli do kánoí a vypluli ve směru dané hvězdy, dokud nenarazili na ostrov, který poté osídlili.[153] Podobné zmínky se vyskytují i v dalších legendách. V samojské kultuře se traduje, že bůh Tagaloa seslal svou dceru Sinu na Zemi ve formě ptáka Tuli, aby našel pevninu.[154] Jako tuli se v samojštině označuje řada druhů brodivých ptáků, mj. kulík pacifický,[155] který migruje mezi arktickými oblastmi a Novým Zélandem.[156]
Vedle ostrovů a atolů v Tichomoří migrace ptáků patrně pomohly Polynésanům objevit i novozélandské souostroví, které představuje jednu z posledních velkých pevnin osídlených lidmi. Na Nový Zéland každoročně migruje přes 20 milionů ptáků, což by patrně jen těžko uniklo obyvatelům polynéských ostrovů, kteří jsou známí výbornými pozorovacími schopnostmi okolní přírody.[154] Jedním z nejčastěji zmiňovaných ptáků, který mohl sloužit jako ukazatel novozélandské pevniny, je kukačka dlouhoocasá, která každoročně migruje mezi hnízdišti na Novém Zélandu a zimovišti v jižním Tichomoří.[154][157] Podobně i kukačka nádherná každoročně migruje mezi novozélandskou pevninou a mikronéskými ostrovy.[157]
Západní tradice
|
I čáp na nebi zná svůj čas, hrdlička, vlaštovka a jeřáb dodržují čas svého příletu, ale můj lid nezná Hospodinovy řády. |
| Starý zákon, Kniha Jeremijáš 8,7[158] |
K nejstarším písemným zmínkám o migraci ptáků v tradici Západu patří Homérova Illiada z 8. století př. n. l., kde je trojská armáda přirovnávána k táhnoucímu hejnu jeřábů.[159] Zmínky o migraci ptáků lze nalézt i ve Starém zákoně, konkrétně v Knize Jób[160] nebo Knize Jeremijáš.[158] Ptačí migrací se zabývali starořečtí klasičtí myslitelé jako byli Hérodotos, Hésiodos a Aristotelés.[161] Aristotelés si mj. všiml, že ptáci před odletem ztloustnou. Tento filosof uvažoval o třech možných vysvětleních sezónního odletu ptáků: ptáci buď migrují do vzdálených končin, nebo se transformují do jiné podoby (tzv. teorie trasmutace), nebo hibernují. Aristotelés zavrhl migraci jako bizarní, avšak transmutací a hibernací se podrobněji zabýval.[162] Aristotelovo uvažování o transmutaci bylo motivováno tím, že zatímco jeden druh odlétá na sever, jiný nedlouho poté přilétá z jihu. Aristotelés proto uvažoval, že se jedná o tytéž ptáky, kteří se však přeměnili na jiný druh. Tento řecký velikán propagoval i teorii hibernace, kterou později šířili jeho následovníci. Podle hibernační teorie se tažní ptáci na chladné období uchylují do jeskyní, dutin stromů či dokonce do bahna močálů a jezer, kde v zimním období hibernují.[163][164]
Migrace vs hibernace vlaštovek

Aristotelova teorie hibernace ptáků se stala převažující teorií vysvětlení každoročního tahu vlaštovek na další 2 tisíce let. V roce 1555 teorii „posvětil“ uppsalský arcibiskup Olaus Magnus, který popsal, jak v severních mořích rybáři vytahují sítě plné ryb a vlaštovek.[165] Podle Magnuse stačilo vlaštovky přemístit do tepla vyhřátého domu, načež vlaštovky ožily, avšak nedlouho na to uhynuly.[161] Vedle hibernace jako možného vysvětlení tahu ptáků se uvažovalo občas i o jiných teoriích. Svérázně se s otázkou vypořádal novoanglický učenec Charles Morton, který v roce 1694 sice správně napsal, že motivací pro migraci ptáků jsou změny v okolním prostředí a v dostupnost potravy, nicméně navrhl, že ptáci odlétají na Měsíc.[166]
První náznaky vyřešení „záhady mizení vlaštovek“ se objevily na přelomu 18. a 19. století. Anglický učenec Thomas Bewick ve svém zásadním díle A History of British Birds (Historie britských ptáků) z roku 1797 zmiňuje výpověď „velice inteligentního lodního kapitána“, který mezi ostrovy Mallorca a Menorca pozoroval velká hejna vlaštovek mířících na sever. Bewick dále zavrhl možnost hibernace vlaštovek ve vodě, což je podle něj chybné tvrzení založené na skutečnosti, že se vlaštovky stahují na noc k vodním plochám, kde hřadují. Snad největší přínos Bewicka k vyřešení otázky mizení vlaštovek bylo to, že několik vlaštovek odchytil a udržoval je v zajetí v teple a suchu po několik britských zim, čímž dokázal, že u nich nedochází k hibernaci. Bewick uzavřel otázku migrace tím, že vlaštovky opouští Británii jednoduše poté, co dojde k vyčerpání jejich potravních zdrojů.[164][167]

Možnost hibernace vlaštovek se dlouho jevila jako logická, protože byla známa u některých jiných zvířat jako jsou obojživelníci a plazi. Některé kultury si vysvětlovaly mizení ptáků po svém. Dodnes se u některých jihoevropských národů traduje, že malé ptáky přenáší přes Středozemní moře větší druhy.[168][169] Podobně se v tradicích některých kmenů původních obyvatel amerického kontinentu traduje, že aljašští kolibříci se přemisťují na zimu na jih na hřbetech bernešek velkých, jimž se zavrtají do peří.[168][170] Přesvědčení o hibernaci vlaštovek však bylo mezi přírodovědci nadále silně zakořeněno. Švédský přírodovědec Carl Linné v roce 1845 vypsal odměnu tomu, kdo vlaštovky v bahně objeví.[170] Americký ornitolog Elliott Coues roce 1878 publikoval seznam hned 182 článků, které se zabývaly hibernací vlaštovek. Sám Coues se k hibernaci stavěl poněkud odměřeně, když napsal: „Nikdy jsem nic takového neviděl, ani neznám nikoho, kdo to [hibernaci] viděl [...]“.[162]
Pfeilstorch a první studia migrace

Významným posunem v rozluštění „záhady“ mizení ptáků byl nález čápa bílého s šípem zabodnutým v krku. Tento jedinec byl odchycen v městě Klütz v dnešním severním Německu v roce 1822 a poskytuje jeden z prvních solidních důkazů dálkové migrace ptáků.[171] Šíp měl na délku kolem 80 cm a byl vyroben z afrického dřeva, čímž bylo jasné, že čáp byl zasažen šípem na území Afriky, odkud vykonal několika tisícikilometrový let až k Baltskému moři, kde byl odchycen.[172] Tento čáp byl nazván pfeilstorch (z německého pfeil, čili „šíp“, a storch, „čáp“). Jedinec byl vypreparován a stal se součástí zoologické sbírky Roztocké univerzity.[171] K roku 2003 bylo evidováno kolem 25 případů pfeilstorch.[172]
V průběhu 19. století se názor vědců na každoroční odlet ptáků postupně začal přiklánět na stranu migrace. Americký ornitolog George Lowery kolem poloviny 20. století vyvinul techniku zaznamenávání nočních migrantů skrze teleskopy namířené k úplňku a počítání proletěných siluet.[173] Klíčovým krokem ke studiu migrace ptáků byl vynález kroužkování, se kterým přišel dánský ornitolog Hans Mortensen na přelomu 19. a 20. století. V prvních desetiletích 20. století začaly vycházet první studie publikující výsledky kroužkovacích projektů.[174]
Moderní metody studia migrace
Ke klíčovým moderním vědeckým metodám studia migrace stále patří kroužkování ptáků. Jedná se o levnou a přístupnou metodu, jak získat důležitá data o migraci. Kroužkuje se pomocí kovových kroužků (nejčastěji hliníkových), které se umístí na běhák ptáka. Ptáci ke kroužkování se nejčastěji chytají v hnízdištích nebo za letu do ptačích sítí.[175]
I když kroužkování přineslo odpovědi na řadu základních otázek spojených s migrací, neposkytuje informace jako např. kde se jedinec pohyboval mezi místem, kde byl okroužkován, a odchytovým místem. Dalším problém kroužkování je nízká návratnost kroužků, které se může pohybovat jen v desetinách procent. Průlom nastal v 90. letech 20. století s příchodem telemetrických zařízení jako jsou satelitní vysílače.[176]

Obecně platí, že telemetrické zařízení umístěné na ptákovi by nemělo přesahovat 3 % jeho hmotnosti.[177] U velkých ptáků je možné používat pokročilá a poměrně těžká zařízení vybavená solárními panely přijímajícími signál satelitní navigace. Tato zařízení se mohou například připevňovat jako obojky na krky nebo na rýdovací pera.[175] Lety velkých ptáků tak lze sledovat i v reálném čase.[178] U menších druhů se používají hlavně ultralehké zařízení zvané geolokátory. Tato miniaturní zařízení mohou vážit i méně než 1 gram a unesou je i tak malé druhy jako vlaštovky. Zařízení získává v určeném časovém intervalu každodenní data o intenzitě osvětlení, z čehož se dají vyvodit zeměpisné souřadnice. Největší nevýhody geolokátorů jsou nutnost odchycení jedinců pro stáhnutí dat[176] a zeměpisné souřadnice odvozené z geolokátorů mají rozptyl ±150 km u zeměpisné šířky a ±200 km u zeměpisné délky.[179] V roce 2015 došlo k prvnímu testu miniaturních zařízení, která dokáží komunikovat se satelitem v určených intervalech (v tomto případě každý 28 dní) a přesto váží kolem 1 g. Tato zařízení byla připevněna na hřbety lesňáčků zlatohlavých (váha jedince kolem 20g). I když bylo ptáky nutné znovu odchytit pro stáhnutí dat, zařízení poskytla cenné informace o zimovištích druhu.[179]

K dalším metodám studia migrace patří radarová zařízení[180] nebo satelitní snímky.[181] Migraci ptáků pomáhají studovat dokonce i astronauti z Mezinárodní vesmírné stanice.[182] Vedle moderních technologií vědci používají i mnohem jednodušší zařízení jako je Emlenův trychtýř. Jedná se v podstatě o plechovou klec ve tvaru převráceného kuželu s malou plošinkou na dně potřenou inkoustem. V době migrace odchycený pták hopká po inkoustu a po stěnách plechové klece obalené papírem, čímž se pak dá rozpoznat, jakým směrem by se nejraději vydal. Stropní část bývá být zakryta jen drátěným pletivem, díky čemuž může pták pozorovat, co je nad ním. V případě potřeby tak lze např. provádět pokusy v planetáriu pomocí manipulace postavení hvězd.[183]
Další moderní metodou studia migrace ptáků je rozbor stabilních izotopů vodíků (deuterium) ve tkáních ptáků (např. v kůži nebo peří). Izotopové složení ptačích tkání odráží izotopové složení jejich potravy, která byla hmyzem či rostliny nasáknuta skrze dešťovou vodu. Obsah deuteria v dešťové vodě je jedinečný pro každou oblast, takže jeho analýzou lze určit místo původu ptáků.[184][185]
Ohrožení a ochrana

Největší ohrožení stěhovavých ptáků představuje člověk a jeho aktivity. Tahové cesty často vedou napříč státy, takže management a ochrana ptáků vyžaduje mezinárodní spolupráci. Pro tyto účely bylo vytvořeno několik mezinárodních dohod. K těm nejdůležitěším patří tzv. Bonnská úmluva,[149] v rámci které byla sjednána i Dohoda o ochraně africko-euroasijských stěhovavých vodních ptáků (AEWA).[186] K signatářům obou dohod patří i Evropská unie, potažmo Česko.[149][186]
Dlouhou dobu se mělo za to, že stěhovaví ptáci během migrace zastavují tam, kde se naskytne vhodná příležitost. Moderní výzkumy se sledovacími zařízeními nicméně ukázaly opak; stěhovaví ptáci často dělají pouze několik málo tahových zastávek na své migrační cestě a jsou tedy citelně závislí na svých odpočinkových stanovištích. To platí zejména v místech, která fungují jako přirozený trychtýř migračních cest jako je např. Střední Amerika, která představuje vzhledem k severoamerické pevnině jen úzký pás pevniny, který řada migračních ptáků využívá.[51] Tyto oblasti je tedy potřeba chránit, což je někdy problematické, protože ptáci svá odpočinková stanoviště často využívají jen několik dní v roce, a může být složité tato místa identifikovat.[187] Vedle ochrany stanovišť se snahy ochranářů soustřeďují mj. na potlačení pytláctví, omezení hazardních bariér (např. drátů elektrického vedení, do kterých tažní ptáci někdy naráží), vědecký výzkum nebo vzdělávání veřejnosti o problematice ochrany migračních druhů.[188]

Poměrně běžným způsobem ochrany ptačích druhů je pomocí metody reintrodukce (tzn. znovu uvedení druhu do míst, odkud byl v minulosti vyhuben). U tažných druhů to je nicméně problematické – jakmile totiž dojde k vyhubení určité populace, dojde i k zániku genů s informacemi o tahové cestě. Známý je případ jeřábů amerických, kteří byli kdysi rozšířeni po celém středozápadě Spojených států, avšak následkem lovu a drastického úbytku přirozeného prostředí se vyskytují již jen v několika málo oblastech. V rámci jednoho z reintrodukčních programů byla do Wisconsinu (sever Spojených států amerických) reintrodukována malá populace jeřábů, která byla již od malička učena následovat ultralehké letadlo. Toto letadlo nakonec jeřáby vedlo přes celé Spojené státy až do jejich zimovišť na Floridě (jih Spojených států amerických). Po skončení zimy se jeřábi dovedli sami vrátit zpět do Wisconsinu.[189]
Některé druhy stěhovavých ptáků jsou následkem působení člověka již vyhynulé. Jedním z nejznámějších případů je holub stěhovavý, jehož počty na severoamerickém kontinentu před příchodem Evropanů dosahovaly kolem 3–5 miliard. Nadměrný lov, odlesňování a dramatický úbytek biotopu však vedlo k vyhnutí druhu.[190] Mezi stěhovavými druhy je velké množství ohrožených i kriticky ohrožených druhů; velká část z nich náleží k trubkonosým.[191]
Na podporu šíření povědomí o potřebě ochrany stěhovavých druhů se každoročně koná Světový den stěhovavých ptáků.[192]
Změna klimatu
Probíhající změna klimatu mají na migraci ptáků dalekosáhlý vliv. Dochází ke změnám v ptačí fenologii, morfologii i načasování migrace. Následkem globálního oteplování nastává po celém světě k časnějším odletům ptáků do zimovišť, čímž se ptáci snaží přizpůsobit stále ranějšímu příchodu jara, se kterým je spojováno množství dostupné potravy jako je nektar, ovoce a hmyz. V průměru se doba odletu migrantů posouvá od pozdního 20. století o půl dne až několik dní vpřed každých deset let.[193][194][195][196]

Tyto změny načasování však mohou vést k prudkým populačním poklesům tažných druhů. Ptáci mohou např. přiletět do zimovišť ještě před tím, než dojde k rojení hmyzu, což vzhledem k vyčerpání ptáků po dlouhém letu může mít kritické následky.[196] V důsledku změny načasování dochází i ke změnám v hnízdních zvyklostech stěhovavých ptáků.[194] Např. doba hnízdění severoamerické vlaštovky stromové se jen mezi lety 1959–1991 posunula vpřed v průměru o 9 dní, což je vedle vyšších teplot spojováno i s ranější dobou květenství tamější vegetace.[197] U nizozemských populací lejsčíků černohlavých sice došlo k posunutí doby kladení vajec vpřed, takže ptáci mají dostatek potravy v době hnízdění, nicméně nedošlo k posunutí samotného migračního letu. Ptáci tak přilétají do Nizozemí příliš pozdě na to, aby stihli vrcholné období výskytu housenek, které představují jeden z jejich klíčových potravních zdrojů. Následkem toho některé nizozemské populace lejsčíků černohlavých poklesly mezi lety přibližně 1985–1995 až o 90 %.[198] Podle výsledků studie 20 druhů evropských migračních druhů se průměrná doba migrantů strávená v Evropě mezi lety 1992–2018 prodloužila o 16 dní. Autoři studie předpokládají, že některé druhy jako konipasi, slavík obecný či budníček větší mohou úplně přestat migrovat.[199][200]
U řady druhů dochází ke zkracování délky migrace, protože ptáci již nemusí létat tak daleko, aby našli vhodné podmínky pro přezimování.[201] Příkladem z českého ptactva mohou být čápi bílí, kteří někdy hnízdí v Řecku nebo Španělsku místo v Africe, nebo některé populace pěnic černohlavých, které zimují ve Velké Británii namísto v Africe.[143] S vyššími teplotami jsou spojovány i změny v morfologii ptáků – u migrujících druhů dochází k postupnému prodlužování křídel a zmenšování těl, což je spojováno s efektivnější migrací. Menší těla totiž lépe odvádí teplo z rozehřátých těl letících ptáků a delší křídla zefektivňují létání.[202][203]
Nejslibnější obranou ptáků proti změnám klimatu je flexibilita migrace. Četné výzkumy ukazují, že úprava načasování migrace vzhledem ke klimatickým změnám má pozitivní vliv na populaci druhu.[204] Zdá se, že migranti na krátkou vzdálenost dokáží reagovat na změny klimatu pružněji než migranti na dlouhou vzdálenost. Dálkoví migranti totiž časují svůj tah hlavně na základě množství denního světla a nemohou na dálku odhadnout podmínky destinace. Naproti tomu migranti táhnoucí krátké vzdálenosti mohou lépe vytušit podmínky v zimovišti / hnízdišti a mohou tak dokonce profitovat z probíhajících změn klimatu (např. v případě časného oteplení hnízdiště se tam mohou vypravit dříve, což jim může poskytnout konkurenční výhodu).[205] Nicméně úprava načasování migrace je u každého druhu jiná a ne vždy se děje dostatečně rychle nebo pružně na to, aby se vyrovnala s následky klimatických změn.[196]
Odraz v kultuře
Migrační druhy ptáků zanechaly svou stopu v mnohých kulturách v podobě mýtů, legend či tradičních domorodých tanců.[206] Ve Starověkém Egyptě bylo na každoroční přílet a odlet migračních druhů nahlíženo jako na součást cyklického bytí světa.[207] Ve staročínské sbírce textů Chuaj-nan-c' je každoroční odlet ptáků přičítán na vrub metamorfóze, čili přeměně ptačích druhů na jiné bytosti. Ve sbírce se mj. píše, že vlaštovky na podzim nalétají do moře a přemění se ve škeble. Podobně i podle starších čínských kalendářů se jestřábi na jaře mění v holuby a myši v křepelky.[207] Migrace ptáků je často spojována se změnou ročních dob.[206] Např. v Česku se vlaštovky a jiřičky přezdívají „poslové jara“.[208] Někdy se za posly jara označují i čápi bílí,[209] kteří jsou hojně spojováni i s nošením dětí.[210] Nošení dětí čápy zvěčnil dánský klasik Hans Christian Andersen ve své poněkud potemnělé pohádce „Čápi“. Švédská spisovatelka Selma Lagerlöf v knize Podivuhodná cesta Nilse Holgerssona Švédskem popsala příběh chlapce Nilse, který cestuje Švédskem na hřbetu husy.[211]
Stěhování ptáků se stalo námětem řady dokumentárních filmů jako je např. Ptačí svět[212] nebo několika dílů dokumentární minisérie Migrations secrètes (Záhady migrace).[213] O zmíněném vedení hus a jeřábů ultralehkým letadlem byl natočen film Cesta domů (1996).[214]
Odkazy
Reference
- Veselovský 2001, s. 306–307.
- ROLLAND, Jonathan; JIGUET, Frédéric; JØNSSON, Knud Andreas; CONDAMINE, Fabien L.; MORLON, Hélène. Settling down of seasonal migrants promotes bird diversification. S. 20140473. Proceedings of the Royal Society B: Biological Sciences [online]. 2014-06-07 [cit. 2021-12-11]. Roč. 281, čís. 1784, s. 20140473. DOI 10.1098/rspb.2014.0473. (anglicky)
- SOMVEILLE, Marius; MANICA, Andrea; BUTCHART, Stuart H. M.; RODRIGUES, Ana S. L. Mapping Global Diversity Patterns for Migratory Birds. S. e70907. PLoS ONE [online]. 2013-08-07 [cit. 2021-12-20]. Roč. 8, čís. 8, s. e70907. DOI 10.1371/journal.pone.0070907. (anglicky)
- The Basics of Bird Migration: How, Why, and Where [online]. The Cornell Lab, 2021-08-01 [cit. 2021-12-11]. Dostupné online. (anglicky)
- BOERE, Gerard C.; STROUD, David A. The flyway concept: what it is and what it isn’t. In: BOERE, G. C.; GALBRAITH, C. A.; STROUD, D. A. Waterbirds around the world: a global overview of the conservation, management and research of the world's waterbird flyways. Edinburgh: The Stationery Office, 2006. Dostupné online. ISBN 0114973334. (anglicky)
- Rappole 2013, s. 1.
- Veselovský 2001, s. 307.
- SORIANO-REDONDO, Andrea; GUTIÉRREZ, Jorge S.; HODGSON, Dave; BEARHOP, Stuart. Migrant birds and mammals live faster than residents. S. 5719. Nature Communications [online]. 2020-12 [cit. 2021-12-16]. Roč. 11, čís. 1, s. 5719. DOI 10.1038/s41467-020-19256-0. (anglicky)
- Newton 2008, s. 396.
- WILBER, Sarah. Aptenodytes forsteri (emperor penguin) [online]. Animal Diversity Web [cit. 2021-12-14]. Dostupné online. (anglicky)
- Newton 2008, s. 228.
- Newton 2008, s. 64.
- DALY, Jon. 'It's unusual': Emus seek greener pastures in long migration south [online]. ABC News, 2021-09-20 [cit. 2021-12-14]. Dostupné online. (anglicky)
- Lincoln 1935, s. 18.
- PANOV, Ilya N. Overlap between moult and autumn migration in passerines in northern taiga zone of Eastern Fennoscandia. S. 33–64. Avian Ecology and Behaviour [online]. 2011 [cit. 2021-12-12]. Roč. 19, s. 33–64. Dostupné online. (anglicky)
- MEISSNER, Włodzimierz. Immature dunlins Calidris alpina migrate towards wintering grounds later than adults in years of low breeding success. S. 47–53. Journal of Ornithology [online]. 2015-01 [cit. 2021-12-12]. Roč. 156, čís. 1, s. 47–53. DOI 10.1007/s10336-014-1132-y. (anglicky)
- MEISSNER, Włodzimierz; WŁODARCZAK-KOMOSIŃSKA, Anna; GÓRECKI, Dariusz; WÓJCIK, Cezary; ŚCIBORSKI, Mateusz; KRUPA, Robert; ZIĘCIK, Piotr. Autumn Migration of Waders (Charadrii) at the Reda Mouth (N Poland). S. 23–39. RING [online]. 2009-01-01. Roč. 31, čís. 1, s. 23–39. DOI 10.2478/v10050-008-0046-6. (anglicky)
- KETTERSON, E. D.; NOLAN, V. Intraspecific variation in avian migration: evolutionary and regulatory aspects. In: RANKIN, M. A. Migration: mechanisms and adaptive significance. University of Texas, Austin: University of Texas Press, 1985. S. 553–579.
- DOBRUSKÁ, Gabriela. Nové poznatky o úžasném letci rorýsovi • Česká společnost ornitologická [online]. Česká společnost ornitologická, 2021-05-25 [cit. 2021-12-14]. Dostupné online. (česky)
- Do hummingbirds migrate in flocks? [online]. The Cornell Lab, 2009-04-01 [cit. 2021-12-13]. Dostupné online. (anglicky)
- Newton 2008, s. 89–91.
- BEAUCHAMP, Guy. Long-distance migrating species of birds travel in larger groups. S. 692–694. Biology Letters [online]. 2011-10-23 [cit. 2021-12-14]. Roč. 7, čís. 5, s. 692–694. DOI 10.1098/rsbl.2011.0243. (anglicky)
- TAYLOR, Alan. The Murmurations of Starlings - The Atlantic. www.theatlantic.com [online]. [cit. 2021-12-29]. Dostupné online. (anglicky)
- Ten Million Starlings Swarm (7 Tonnes of Bird Poo) [online]. BBC Earth, 2021-04-25 [cit. 2021-12-29]. Dostupné online.
- Why Birds Fly in a V Formation [online]. Scientific American [cit. 2022-02-02]. Dostupné online. (anglicky)
- PORTUGAL, Steven J.; HUBEL, Tatjana Y.; FRITZ, Johannes; HEESE, Stefanie; TROBE, Daniela; VOELKL, Bernhard; HAILES, Stephen. Upwash exploitation and downwash avoidance by flap phasing in ibis formation flight. S. 399–402. Nature [online]. 2014-01 [cit. 2022-02-02]. Roč. 505, čís. 7483, s. 399–402. DOI 10.1038/nature12939. (anglicky)
- Newton 2008, s. 54.
- WEIMERSKIRCH, Henri; MARTIN, Julien; CLERQUIN, Yannick; ALEXANDRE, Peggy; JIRASKOVA, Sarka. Energy saving in flight formation. S. 697–698. Nature [online]. 2001-10 [cit. 2021-12-16]. Roč. 413, čís. 6857, s. 697–698. DOI 10.1038/35099670. (anglicky)
- Science Reference Section. Why do geese fly in a V? [online]. Library of Congress, 2019-11-19 [cit. 2021-12-16]. Dostupné online. (anglicky)
- Newton 2008, s. 78.
- Newton 2008, s. 79–80.
- SCOTT, Graham R.; HAWKES, Lucy A.; FRAPPELL, Peter B.; BUTLER, Patrick J.; BISHOP, Charles M.; MILSOM, William K. How Bar-Headed Geese Fly Over the Himalayas. S. 107–115. Physiology [online]. 2015-03 [cit. 2021-12-14]. Roč. 30, čís. 2, s. 107–115. DOI 10.1152/physiol.00050.2014. (anglicky)
- HAWKES, L. A.; BALACHANDRAN, S.; BATBAYAR, N.; BUTLER, P. J.; FRAPPELL, P. B.; MILSOM, W. K.; TSEVEENMYADAG, N. The trans-Himalayan flights of bar-headed geese (Anser indicus). S. 9516–9519. Proceedings of the National Academy of Sciences [online]. 2011-06-07 [cit. 2021-12-14]. Roč. 108, čís. 23, s. 9516–9519. DOI 10.1073/pnas.1017295108. (anglicky)
- SENNER, Nathan R.; STAGER, Maria; VERHOEVEN, Mo A.; CHEVIRON, Zachary A.; PIERSMA, Theunis; BOUTEN, Willem. High-altitude shorebird migration in the absence of topographical barriers: avoiding high air temperatures and searching for profitable winds. S. 20180569. Proceedings of the Royal Society B: Biological Sciences [online]. 2018-06-27 [cit. 2021-12-14]. Roč. 285, čís. 1881, s. 20180569. DOI 10.1098/rspb.2018.0569. (anglicky)
- Newton 2008, s. 80.
- Čapí migrace [online]. Česká společnost ornitologická [cit. 2021-12-14]. Dostupné online. (česky)
- CORSO, Andrea. European Birding Hot Spot: The Strait of Messina, southern Italy. [online]. Surfbirds [cit. 2021-12-14]. Dostupné online. (anglicky)
- Newton 2008, s. 14.
- Kolektiv autorů. Strategie ochrany biologické rozmanitosti České republiky 2016–2025. [s.l.]: Ministerstvo životního prostředí, 2016. Dostupné online. ISBN 978-80-7212-609-5. S. 112.
- Newton 2008, s. 14–15.
- Newton 2008, s. 259.
- RUEGG, Kristen C.; SMITH, Thomas B. Not as the crow flies: a historical explanation for circuitous migration in Swainson's thrush (Catharus ustulatus). S. 1375–1381. Proceedings of the Royal Society of London. Series B: Biological Sciences [online]. 2002-07-07 [cit. 2021-12-13]. Roč. 269, čís. 1498, s. 1375–1381. DOI 10.1098/rspb.2002.2032. (anglicky)
- KLVAŇA, Petr. Podzimní cesty našich tažných ptáků. S. 11–15. Ptačí svět [online]. Česká společnost ornitologická, 2017 [cit. 2021-12-16]. Roč. XXIV, čís. 3, s. 11–15.
- PROCHÁZKA, Pavel. Podzimní tahové cesty našich migrantů [online]. ČSO, 2017 [cit. 2022-01-27]. Dostupné online. (anglicky)
- Atlas migrace ptáků. Vltava [online]. 2008-12-11 [cit. 2022-02-01]. Dostupné online. (česky)
- Newton 2008, s. 3.
- RAMACHANDRA, T. V.; MAHAPATRA, Durga Madhab; RAO, Sankara Kolluri; NATHAN, M. Bhoomi. Environmental Impact Assessment of the National Large Solar Telescope Project and Its Ecological Impact in Merak Area [online]. Energy & Wetland Research Group Centre for Ecological Sciences, Indian Institute of Science, Bengalore, India, 2013 [cit. 2021-12-11]. S. 69. Dostupné online. (anglicky)
- Newton 2008, s. 8.
- JAHN, Alex E.; BRAVO, Susana P.; CUETO, Víctor R. Patterns of partial avian migration in northern and southern temperate latitudes of the New World. Emu - Austral Ornithology. 2012-03-01, roč. 112, čís. 1, s. 17–22. Dostupné online [cit. 2021-12-12]. ISSN 0158-4197. DOI 10.1071/MU10091. (anglicky)
- What We Do [online]. Birdlife International [cit. 2021-12-14]. Dostupné online. (anglicky)
- Stopover Habitats and What Birds Need on Migration. American Bird Conservancy [online]. 2017-04-27 [cit. 2021-12-14]. Dostupné online. (anglicky)
- FERRETTI, Andrea; MAGGINI, Ivan; LUPI, Sara. The amount of available food affects diurnal locomotor activity in migratory songbirds during stopover. Scientific Reports. 2019-12, roč. 9, čís. 1, s. 19027. Dostupné online [cit. 2021-12-14]. ISSN 2045-2322. DOI 10.1038/s41598-019-55404-3. PMID 31836848. (anglicky)
- Černý 1980, s. 11.
- Vrcholí migrace ptáků – pomozme jim v letu o život! • Česká společnost ornitologickáVrcholí migrace ptáků – pomozme jim v letu o život! [online]. Česká společnost ornitologická, 2021-10-19 [cit. 2021-12-14]. Dostupné online. (anglicky)
- The ecology, behavior, and conservation of migratory birds. www.usgs.gov [online]. U.S. Geological Survey [cit. 2022-01-26]. Dostupné online. (anglicky)
- NUWER, Rachel. Fish Eggs Survive Journey through a Duck [online]. Scientific American, 2020-09-01 [cit. 2022-01-26]. Dostupné online. (anglicky)
- LENNOX, Robert J.; CHAPMAN, Jacqueline M.; SOULIERE, Christopher M.; TUDORACHE, Christian; WIKELSKI, Martin; METCALFE, Julian D.; COOKE, Steven J. Conservation physiology of animal migration. Conservation Physiology [online]. 2016 [cit. 2022-01-26]. Roč. 4, čís. 1. DOI 10.1093/conphys/cov072. (anglicky)
- FRITZSCHE MCKAY, Alexa; HOYE, Bethany J. Are Migratory Animals Superspreaders of Infection?. S. 260–267. Integrative and Comparative Biology [online]. 2016-08-01 [cit. 2022-01-25]. Roč. 56, čís. 2, s. 260–267. DOI 10.1093/icb/icw054. (anglicky)
- COHEN, Emily B.; AUCKLAND, Lisa D.; MARRA, Peter P.; HAMER, Sarah A. Avian Migrants Facilitate Invasions of Neotropical Ticks and Tick-Borne Pathogens into the United States. S. 8366–8378. Applied and Environmental Microbiology [online]. 2015-12-15 [cit. 2022-01-25]. Roč. 81, čís. 24, s. 8366–8378. DOI 10.1128/AEM.02656-15. (anglicky)
- VERHAGEN, Josanne H.; HERFST, Sander; FOUCHIER, Ron A. M. How a virus travels the world. S. 616–617. Science [online]. 2015-02-06 [cit. 2022-01-25]. Roč. 347, čís. 6222, s. 616–617. DOI 10.1126/science.aaa6724. (anglicky)
- Newton 2008, s. 5.
- Rappole 2013, s. 15.
- Rappole 2013, s. 6.
- Song Sparrow Range Map [online]. The Cornell Lab of Ornithology [cit. 2021-12-16]. Dostupné online. (anglicky)
- RYCHLÍKOVÁ, H.; FORMÁNEK, J.; ŠKOPEK, J. Pták roku 2001: Kavka Obecná. [s.l.]: Česká společnost ornitologická, 2001. Dostupné online.
- Velká podzimní migrace [online]. Česká společnost ornitologická [cit. 2021-12-16]. Dostupné online.
- TURNER, Angela K. Swallows & martins: an identification guide and handbook. Boston: Houghton Mifflin, 1989. Dostupné online. ISBN 0395511747. S. 164–169. (anglicky)
- VAN GILS, Jan A; BATTLEY, Phil F; PIERSMA, Theunis; DRENT, Rudi. Reinterpretation of gizzard sizes of red knots world-wide emphasises overriding importance of prey quality at migratory stopover sites. S. 2609–2618. Proceedings of the Royal Society B: Biological Sciences [online]. 2005-12-22 [cit. 2021-12-14]. Roč. 272, čís. 1581, s. 2609–2618. DOI 10.1098/rspb.2005.3245. (anglicky)
- Longest non-stop migration by a bird [online]. Guinness World Records [cit. 2021-04-30]. Dostupné online. (anglicky)
- HROMÁDKOVÁ, T; PAVEL, V; FLOUSEK, J; BRIEDIS, M. Seasonally specific responses to wind patterns and ocean productivity facilitate the longest animal migration on Earth. S. 1–12. Marine Ecology Progress Series [online]. 2020-03-19 [cit. 2021-12-11]. Roč. 638, s. 1–12. DOI 10.3354/meps13274. (anglicky)
- Čápi a vlaštovky se s tím moc nepářou, do Afriky doletí na jeden zátah [online]. Český rozhlas, 2017-10-17 [cit. 2021-12-20]. Dostupné online.
- BAIRLEIN, Franz; NORRIS, D. Ryan; NAGEL, Rolf; BULTE, Marc; VOIGT, Christian C.; FOX, James W.; HUSSELL, David J. T. Cross-hemisphere migration of a 25 g songbird. S. 505–507. Biology Letters [online]. 2012-08-23. Roč. 8, čís. 4, s. 505–507. DOI 10.1098/rsbl.2011.1223. (anglicky)
- Featherweight songbird is a long-distance champ [online]. Phys.org [cit. 2021-12-14]. Dostupné online. (anglicky)
- PYLE, Peter. Seabirds [online]. USGS - Circular 1198, 2001 [cit. 2021-12-14]. S. 154. Dostupné online. (anglicky)
- SHAFFER, S. A.; TREMBLAY, Y.; WEIMERSKIRCH, H.; SCOTT, D.; THOMPSON, D. R.; SAGAR, P. M.; MOLLER, H. Migratory shearwaters integrate oceanic resources across the Pacific Ocean in an endless summer. S. 12799–12802. Proceedings of the National Academy of Sciences [online]. 2006-08-22 [cit. 2021-05-18]. Roč. 103, čís. 34, s. 12799–12802. DOI 10.1073/pnas.0603715103. (anglicky)
- Rappole 2013, s. 12.
- BARÇANTE, Luciana; M. VALE, Mariana; S. ALVES, Maria Alice. Altitudinal migration by birds: a review of the literature and a comprehensive list of species. S. 321–335. Journal of Field Ornithology [online]. 2017-12 [cit. 2021-12-16]. Roč. 88, čís. 4, s. 321–335. DOI 10.1111/jofo.12234. (anglicky)
- BOYLE, W. Alice. Altitudinal bird migration in North America. S. 443–465. The Auk [online]. 2017-04 [cit. 2021-12-16]. Roč. 134, čís. 2, s. 443–465. DOI 10.1642/AUK-16-228.1. (anglicky)
- Newton 2008, s. 379.
- BOYLE, W. Alice; CONWAY, Courtney J.; BRONSTEIN, Judith L. Why do some, but not all, tropical birds migrate? A comparative study of diet breadth and fruit preference. S. 219–236. Evolutionary Ecology [online]. 2011-01 [cit. 2021-12-16]. Roč. 25, čís. 1, s. 219–236. DOI 10.1007/s10682-010-9403-4. (anglicky)
- HOBSON, Keith A.; WASSENAAR, Len I.; MILÁ, Borja; LOVETTE, Irby; DINGLE, Caroline; SMITH, Thomas B. Stable isotopes as indicators of altitudinal distributions and movements in an Ecuadorean hummingbird community. S. 302–308. Oecologia [online]. 2003-07-01 [cit. 2021-12-16]. Roč. 136, čís. 2, s. 302–308. DOI 10.1007/s00442-003-1271-y. (anglicky)
- Rappole 2013, s. 14.
- Shortest bird migration [online]. Guinness World Records [cit. 2021-12-16]. Dostupné online. (anglicky)
- CHAN, Ken. Partial migration in Australian landbirds: a review. S. 281–292. Emu - Austral Ornithology [online]. 2001-12 [cit. 2021-12-12]. Roč. 101, čís. 4, s. 281–292. DOI 10.1071/MU00034. (anglicky)
- DINGLE, Hugh. Bird migration in the southern hemisphere: a review comparing continents. S. 341–359. Emu - Austral Ornithology [online]. 2008-12 [cit. 2021-12-12]. Roč. 108, čís. 4, s. 341–359. DOI 10.1071/MU08010. (anglicky)
- LUNDBERG, Per. The evolution of partial migration in Birds. S. 172–175. Trends in Ecology & Evolution [online]. 1988-07 [cit. 2021-12-12]. Roč. 3, čís. 7, s. 172–175. DOI 10.1016/0169-5347(88)90035-3. (anglicky)
- BOLAND, John M. Leapfrog Migration in North American Shorebirds: Intra- and Interspecific Examples. The Condor [online]. Oxford University Press, 1990-05 [cit. 2021-12-12]. Roč. 92, čís. 2. DOI 10.2307/1368226. (anglicky)
- BELL, Christopher P. Leap-Frog Migration in the Fox Sparrow: Minimizing the Cost of Spring Migration. S. 470–477. The Condor [online]. 1997-05 [cit. 2021-12-12]. Roč. 99, čís. 2, s. 470–477. DOI 10.2307/1369953. (anglicky)
- ÅKESSON, Susanne; ATKINSON, Phil W.; BERMEJO, Ana; PUENTE, Javier; FERRI, Mauro; HEWSON, Chris M.; HOLMGREN, Jan. Evolution of chain migration in an aerial insectivorous bird, the common swift Apus apus. S. 2377–2391. Evolution [online]. 2020-10 [cit. 2021-12-12]. Roč. 74, čís. 10, s. 2377–2391. DOI 10.1111/evo.14093. (anglicky)
- Newton 2008, s. 87.
- ALERSTAM, Thomas. Flight by night or day? Optimal daily timing of bird migration. Journal of Theoretical Biology. 2009-06, roč. 258, čís. 4, s. 530–536. Dostupné online [cit. 2021-12-16]. DOI 10.1016/j.jtbi.2009.01.020. (anglicky)
- Newton 2008, s. 85–89.
- MOORE, Ashli; BARTELL, Paul. Avian Migration: The Ultimate Red-Eye Flight [online]. American Scientist, 2017-02-06 [cit. 2021-12-15]. Dostupné online. (anglicky)
- Newton 2008, s. 87–88.
- Eleonora's Falcons imprison live birds for a later meal [online]. 2015-08-26 [cit. 2021-12-11]. Dostupné online. (anglicky)
- QNINBA, A.; BENHOUSSA, A.; RADI, M.; EL IDRISSI, A.; BOUSADIK, H.; BADAOUI, B.; EL AGBANI, M. A. Mode de prédation très particulier du Faucon d’Éléonore (Falco eleonorae) sur l’Archipel d’Essaouira, Maroc Atlantique. S. 149–150. Alauda [online]. 2015 [cit. 2021-12-11]. Roč. 83, s. 149–150. (francouzsky)
- IBANEZ, C.; JUSTE, J.; GARCIA-MUDARRA, J. L.; AGIRRE-MENDI, P. T. Bat predation on nocturnally migrating birds. S. 9700–9702. Proceedings of the National Academy of Sciences [online]. 2001-08-14 [cit. 2021-12-11]. Roč. 98, čís. 17, s. 9700–9702. DOI 10.1073/pnas.171140598. (anglicky)
- Newton 2008, s. 86.
- RATTENBORG, Niels C; MANDT, Bruce H; OBERMEYER, William H; WINSAUER, Peter J; HUBER, Reto; WIKELSKI, Martin; BENCA, Ruth M. Migratory Sleeplessness in the White-Crowned Sparrow (Zonotrichia leucophrys gambelii). S. e212. PLoS Biology [online]. 2004-07-13 [cit. 2021-12-20]. Roč. 2, čís. 7, s. e212. DOI 10.1371/journal.pbio.0020212. (anglicky)
- FARNSWORTH, Andrew. Flight Calls and Their Value for Future Ornithological Studies and Conservation Research. S. 733–746. The Auk [online]. 2005-07-01 [cit. 2021-12-15]. Roč. 122, čís. 3, s. 733–746. DOI 10.1093/auk/122.3.733. (anglicky)
- GRIFFITHS, Emily T.; KEEN, Sara C.; LANZONE, Michael; FARNSWORTH, Andrew. Can Nocturnal Flight Calls of the Migrating Songbird, American Redstart, Encode Sexual Dimorphism and Individual Identity?. S. e0156578. PLOS ONE [online]. 2016-06-10 [cit. 2021-12-15]. Roč. 11, čís. 6, s. e0156578. DOI 10.1371/journal.pone.0156578. (anglicky)
- PULIDO, Francisco. The Genetics and Evolution of Avian Migration. S. 165–174. BioScience [online]. 2007-02-01 [cit. 2021-12-12]. Roč. 57, čís. 2, s. 165–174. DOI 10.1641/B570211. (anglicky)
- CHERNETSOV, Nikita; BERTHOLD, Peter; QUERNER, Ulrich. Migratory orientation of first-year white storks ( Ciconia ciconia ): inherited information and social interactions. S. 937–943. Journal of Experimental Biology [online]. 2004-02-22 [cit. 2021-12-14]. Roč. 207, čís. 6, s. 937–943. DOI 10.1242/jeb.00853. (anglicky)
- TOEWS, David P. L.; TAYLOR, Scott A.; STREBY, Henry M.; KRAMER, Gunnar R.; LOVETTE, Irby J. Selection on VPS13A linked to migration in a songbird. S. 18272–18274. Proceedings of the National Academy of Sciences [online]. 2019-09-10 [cit. 2021-12-20]. Roč. 116, čís. 37, s. 18272–18274. DOI 10.1073/pnas.1909186116. (anglicky)
- PULIDO, F.; BERTHOLD, P.; MOHR, G.; QUERNER, U. Heritability of the timing of autumn migration in a natural bird population. S. 953–959. Proceedings of the Royal Society of London. Series B: Biological Sciences [online]. 2001-05-07 [cit. 2021-12-12]. Roč. 268, čís. 1470, s. 953–959. DOI 10.1098/rspb.2001.1602. (anglicky)
- BERTHOLD, P.; HELBIG, A. J.; MOHR, G.; QUERNER, U. Rapid microevolution of migratory behaviour in a wild bird species. S. 668–670. Nature [online]. 1992-12. Roč. 360, čís. 6405, s. 668–670. DOI 10.1038/360668a0. (anglicky)
- YOUNG, Ed. On The Evolution of Migration [online]. National Geographic, 2014-08-04 [cit. 2021-12-12]. Dostupné online. (anglicky)
- Origin and evolution of migration [online]. Britannica [cit. 2021-12-12]. Dostupné online. (anglicky)
- WINGER, Benjamin M.; AUTERI, Giorgia G.; PEGAN, Teresa M.; WEEKS, Brian C. A long winter for the Red Queen: rethinking the evolution of seasonal migration. S. 737–752. Biological Reviews [online]. 2019-06 [cit. 2021-12-13]. Roč. 94, čís. 3, s. 737–752. DOI 10.1111/brv.12476. (anglicky)
- SALEWSKI, Volker; BRUDERER, Bruno. The evolution of bird migration—a synthesis. S. 268–279. Naturwissenschaften [online]. 2007-03-20 [cit. 2021-12-13]. Roč. 94, čís. 4, s. 268–279. DOI 10.1007/s00114-006-0186-y. (anglicky)
- The Evolution of Bird Migration [online]. The Cornell Lab, 2017-04-11 [cit. 2021-12-13]. Dostupné online. (anglicky)
- ZINK, Robert M.; GARDNER, Aubrey S. Glaciation as a migratory switch. S. e1603133. Science Advances [online]. 2017-09-29 [cit. 2021-12-13]. Roč. 3, čís. 9, s. e1603133. DOI 10.1126/sciadv.1603133. (anglicky)
- GWINNER, E. Circadian and circannual programmes in avian migration. S. 39–48. Journal of Experimental Biology [online]. 1996-01-01 [cit. 2021-12-20]. Roč. 199, čís. 1, s. 39–48. DOI 10.1242/jeb.199.1.39. (anglicky)
- POKROVSKY, Ivan; KÖLZSCH, Andrea; SHERUB, Sherub; FIEDLER, Wolfgang; GLAZOV, Peter; KULIKOVA, Olga; WIKELSKI, Martin. Longer days enable higher diurnal activity for migratory birds. S. 2161–2171. Journal of Animal Ecology [online]. 2021-09 [cit. 2021-12-20]. Roč. 90, čís. 9, s. 2161–2171. DOI 10.1111/1365-2656.13484. (anglicky)
- WILTSCHKO, Roswitha; WILTSCHKO, Wolfgang. Chapter Seven - Avian Navigation: A Combination of Innate and Learned Mechanisms. Příprava vydání Marc Naguib, H. Jane Brockmann, John C. Mitani, Leigh W. Simmons, Louise Barrett, Sue Healy, Peter J. B. Slater. Svazek 47. [s.l.]: Academic Press, 2015. Dostupné online. DOI 10.1016/bs.asb.2014.12.002. S. 229–310. (anglicky)
- REYNOLDS, Andrew M.; CECERE, Jacopo G.; PAIVA, Vitor H. Pelagic seabird flight patterns are consistent with a reliance on olfactory maps for oceanic navigation. Proceedings of the Royal Society B: Biological Sciences. 2015-07-22, roč. 282, čís. 1811, s. 20150468. Dostupné online [cit. 2021-12-22]. ISSN 0962-8452. DOI 10.1098/rspb.2015.0468. PMID 26136443. (anglicky)
- The Basics: How Birds Navigate When They Migrate. All About Birds [online]. 2008-04-01 [cit. 2021-12-22]. Dostupné online. (anglicky)
- Newton 2008, s. 85.
- LÁZŇOVSKÝ, Matouš. Cestování podle siločar v oku [online]. Lidovky, 2008-05-02 [cit. 2021-12-15]. Dostupné online.
- HEYERS, Dominik; MANNS, Martina; LUKSCH, Harald; GÜNTÜRKÜN, Onur; MOURITSEN, Henrik. A Visual Pathway Links Brain Structures Active during Magnetic Compass Orientation in Migratory Birds. S. e937. PLoS ONE [online]. 2007-09-26 [cit. 2021-12-15]. Roč. 2, čís. 9, s. e937. DOI 10.1371/journal.pone.0000937. (anglicky)
- THORUP, Kasper. Reverse migration as a cause of vagrancy. Bird Study. 2004-10-01, roč. 51, čís. 3, s. 228–238. Dostupné online [cit. 2021-12-28]. ISSN 0006-3657. DOI 10.1080/00063650409461358. (anglicky)
- OGDEN, Lesley Evans. Vagrant Birds May Portend Species Distribution in Climate-Changed World. Scientific American [online]. 2016-08-19 [cit. 2021-12-28]. Dostupné online. (anglicky)
- THORUP, Kasper; ALERSTAM, Thomas; HAKE, Mikael. Bird orientation: compensation for wind drift in migrating raptors is age dependent. Proceedings of the Royal Society of London. Series B: Biological Sciences. 2003-08-07, roč. 270, čís. suppl_1, s. S8–S11. Dostupné online [cit. 2021-12-28]. DOI 10.1098/rsbl.2003.0014. PMID 12952622. (anglicky)
- ELKINS, Norman. Weather and bird migration. S. 238–256. British Birds [online]. 2005 [cit. 2021-12-28]. Roč. 98, s. 238–256. (anglicky)
- SANDBERG, Roland. Interaction of body condition and magnetic orientation in autumn migrating robins, Erithacus rubecula. S. 679–686. Animal Behaviour [online]. 1994-03-01 [cit. 2021-12-28]. Roč. 47, čís. 3, s. 679–686. DOI 10.1006/anbe.1994.1092. (anglicky)
- GILROY, James; LEES, Alexander Charles. Vagrancy theories: Are autumn vagrants really reverse migrants?. S. 427–438. British Birds [online]. 2003 [cit. 2021-12-28]. Roč. 96, čís. 9, s. 427–438. Dostupné online. (anglicky)
- LEES, Alexander C.; GILROY, James J. Bird migration: When vagrants become pioneers. S. R1568–R1570. Current Biology [online]. 2021-12-20 [cit. 2021-12-28]. Roč. 31, čís. 24, s. R1568–R1570. DOI 10.1016/j.cub.2021.10.058. (anglicky)
- DUFOUR, Paul; DE FRANCESCHI, Christophe; DONIOL-VALCROZE, Paul; JIGUET, Frédéric; GUÉGUEN, Maya; RENAUD, Julien; LAVERGNE, Sébastien. A new westward migration route in an Asian passerine bird. S. 5590–5596.e4. Current Biology [online]. 2021-12. Roč. 31, čís. 24, s. 5590–5596.e4. DOI 10.1016/j.cub.2021.09.086. (anglicky)
- GUGLIELMO, Christopher G. Obese super athletes: fat-fueled migration in birds and bats. Journal of Experimental Biology. 2018-03-07, roč. 221, čís. Suppl_1, s. jeb165753. Dostupné online [cit. 2021-12-27]. ISSN 0022-0949. DOI 10.1242/jeb.165753. (anglicky)
- Bird Migration | How Do Birds Prepare For Migration?. The RSPB [online]. [cit. 2021-12-28]. Dostupné online. (anglicky)
- GOYMANN, Wolfgang; LUPI, Sara; KAIYA, Hiroyuki. Ghrelin affects stopover decisions and food intake in a long-distance migrant. Proceedings of the National Academy of Sciences. 2017-02-21, roč. 114, čís. 8, s. 1946–1951. PMID 28167792. Dostupné online [cit. 2021-12-28]. ISSN 0027-8424. DOI 10.1073/pnas.1619565114. PMID 28167792. (anglicky)
- ARAÚJO, Pedro M.; VIEGAS, Ivan; ROCHA, Afonso D. Understanding how birds rebuild fat stores during migration: insights from an experimental study. Scientific Reports. 2019-07-11, roč. 9, čís. 1, s. 10065. Dostupné online [cit. 2021-12-27]. ISSN 2045-2322. DOI 10.1038/s41598-019-46487-z. PMID 31296911. (anglicky)
- BAUCHINGER, Ulf; KOLB, Harald; AFIK, Danny. Blackcap Warblers Maintain Digestive Efficiency by Increasing Digesta Retention Time on the First Day of Migratory Stopover. Physiological and Biochemical Zoology. 2009-09, roč. 82, čís. 5, s. 541–548. Dostupné online [cit. 2022-01-03]. ISSN 1522-2152. DOI 10.1086/603638. (anglicky)
- HUME, I. D.; BIEBACH, H. Digestive tract function in the long-distance migratory garden warbler, Sylvia borin. Journal of Comparative Physiology B. 1996-10, roč. 166, čís. 6, s. 388–395. Dostupné online [cit. 2022-01-03]. ISSN 0174-1578. DOI 10.1007/BF02336922. (anglicky)
- PIERSMA, Theunis; GILL, Robert E. Guts Don't Fly: Small Digestive Organs in Obese Bar-Tailed Godwits. S. 196–203. The Auk [online]. 1998 [cit. 2021-05-01]. Roč. 115, čís. 1, s. 196–203. DOI 10.2307/4089124. (anglicky)
- BATTLEY, Phil F.; PIERSMA, Theunis. Body Composition and Flight Ranges of Bar-Tailed Godwits (Limosa lapponica baueri) from New Zealand (Composition Corporelle et Distance de Vol chez Limosa lapponica baueri de Nouvelle-Zélande). S. 922–937. The Auk [online]. 2005 [cit. 2021-05-01]. Roč. 122, čís. 3, s. 922–937. Dostupné online. ISSN 0004-8038. (anglicky)
- LANDYS-CIANNELLI, M. M.; PIERSMA, T.; JUKEMA, J. Strategic size changes of internal organs and muscle tissue in the Bar-tailed Godwit during fat storage on a spring stopover site. Functional Ecology. 2003-04, roč. 17, čís. 2, s. 151–159. Dostupné online [cit. 2022-01-03]. ISSN 0269-8463. DOI 10.1046/j.1365-2435.2003.00715.x. (anglicky)
- RATTENBORG, Niels C.; VOIRIN, Bryson; CRUZ, Sebastian M. Evidence that birds sleep in mid-flight. Nature Communications. 2016-08-03, roč. 7, čís. 1, s. 12468. Dostupné online [cit. 2021-12-28]. ISSN 2041-1723. DOI 10.1038/ncomms12468. PMID 27485308. (anglicky)
- LIECHTI, Felix; WITVLIET, Willem; WEBER, Roger. First evidence of a 200-day non-stop flight in a bird. Nature Communications. 2013-10-08, roč. 4, čís. 1, s. 2554. Dostupné online [cit. 2021-12-28]. ISSN 2041-1723. DOI 10.1038/ncomms3554. (anglicky)
- UNDERHILL, L. G.; PRYS-JONES, R. P.; DOWSETT, R. J. The biannual primary moult of Willow Warblers Phylloscopus trochilus in Europe and Africa. Ibis. 1992, roč. 134, čís. 3, s. 286–297. Dostupné online [cit. 2021-12-27]. ISSN 1474-919X. DOI 10.1111/j.1474-919X.1992.tb03811.x. (anglicky)
- PEARSON, D. J.; BACKHURST, G. C. Moult in the river Warbler Locustella fluviatilis. Ringing & Migration. 1983-10-01, roč. 4, čís. 4, s. 227–230. Dostupné online [cit. 2021-12-27]. ISSN 0307-8698. DOI 10.1080/03078698.1983.9673810. (anglicky)
- HEDENSTRÖM, Anders. Adaptations to migration in birds: behavioural strategies, morphology and scaling effects. Philosophical Transactions of the Royal Society B: Biological Sciences. 2008-01-27, roč. 363, čís. 1490, s. 287–299. Dostupné online [cit. 2021-12-27]. DOI 10.1098/rstb.2007.2140. PMID 17638691. (anglicky)
- Miliardy ptáků se vracejí z teplých krajin, někteří překonají i tisíce kilometrů. ČT24 [online]. [cit. 2022-01-27]. Dostupné online. (česky)
- HAHN, Steffen; BAUER, Silke; LIECHTI, Felix. The Natural Link between Europe and Africa: 2.1 Billion Birds on Migration. Oikos. 2009, roč. 118, čís. 4, s. 624–626. Dostupné online [cit. 2022-01-27]. ISSN 0030-1299. (anglicky)
- VERMOUZEK, Zdeněk; ŠÍREK, Jiří. Seznam ptáků Česka. [s.l.]: Česká společnost ornitologická, 2016. Dostupné online. ISBN 978-80-87572-24-5.
- ŠIMURDA, Jakub. Kam mizí ptáci na zimu? [online]. Priroda.cz, 2010-11-01 [cit. 2022-01-27]. Dostupné online.
- Cepák et al., s. 11.
- LUČAN, Radek K.; LUČANOVÁ, Anna; VAVŘÍK, Martin. Červenohorské sedlo: historie a současnost sledování (nejen) ptačí migrace na našich horách. Časopis Ochrana Přírody [online]. [cit. 2022-01-27]. Dostupné online.
- Úmluva o ochraně stěhovavých druhů volně žijících živočichů (Bonnská úmluva) [online]. Ministerstvo životního prostředí [cit. 2021-12-30]. Dostupné online.
- Malta – poslední zastávka tažných ptáků? [online]. Ekolist, 2010-04-14 [cit. 2022-01-27]. Dostupné online.
- Cyprus poachers kill 2.3 million songbirds in 2016. phys.org [online]. [cit. 2022-01-27]. Dostupné online. (anglicky)
- Ve Středomoří pytláci pozabíjí ročně 25 milionů ptáků. Čeští experti se snaží situaci zvrátit. ct24.ceskatelevize.cz [online]. ČT24 [cit. 2022-01-28]. Dostupné online. (česky)
- LEWIS, David. We the Navigators: The Ancient Art of Lanfinding in the Pacific. Honolulu, Hawaii: Universitz of Hawaii Press, 1994. ISBN 9780824158231. S. 214–216. (anglicky)
- CROWE, Andrew. Pathway of the birds: the voyaging achievements of Māori and their Polynesian ancestors. Auckland, New Zealand: David Bateman LTD, 2019. ISBN 9781869539610. S. 142–152. (anglicky)
- RICHTER-GRAVIER, Raphael. Manu narratives of Polynesia A comparative study of birds in 300 traditional Polynesian stories. Dunedin, New Zealand, 2019 [cit. 2021]. PhD Thesis. University of Otago. Vedoucí práce Michael Reilly. Dostupné online.
- SZABO, M. J. Pacific golden plover [online]. New Zealand Birds Online, 2013 [cit. 2021-12-09]. Dostupné online. (anglicky)
- Canoe Navigation, Locating land [online]. Te Ara: The Encyclopedia of New Zealand [cit. 2021-12-09]. Dostupné online. (anglicky)
- Jer 8, 7 (Kral, ČEP)
- Homérova Ilias, zpěv třetí.
- Job 39, 26 (Kral, ČEP)
- Lincoln 1935, s. 4.
- POMEROY, Ross. The Weirdest Ideas About Bird Migration [online]. 2018-02-19 [cit. 2021-12-10]. Dostupné online. (anglicky)
- Lincoln 1935, s. 4–5.
- FONE, Martin. Curious Questions: How did a stork with a spear through its neck solve the mystery of the migration of birds? [online]. Country Life, 2020-01-11 [cit. 2021-12-10]. Dostupné online. (anglicky)
- Review: The Wisdom of Birds by Tim Birkhead [online]. 2008-10-17 [cit. 2021-12-10]. Dostupné online. (anglicky)
- HARRISON, Thomas P. Birds in the Moon. S. 323–330. Isis [online]. 1954-12 [cit. 2021-12-10]. Roč. 45, čís. 4, s. 323–330. DOI 10.1086%2F348354. (anglicky)
- BEWICK, Thomas. History of British Birds. Newcastle: Beilby and Bewick, 1847. S. 300–303. (anglicky)
- Lincoln 1935, s. 5.
- BROSNAN, Elise. Bird of the Month: The Pfeilstorch [online]. Audubon Society of Northern Virginia, 2019-09-01 [cit. 2021-12-10]. Dostupné online. (anglicky)
- Veselovský 2001, s. 306.
- RICHTER, Stefan; BICK, Andreas. Zoological Collection Chapter 49. Zoological Collections [online]. Springer International Publishing, 2018. DOI 0.1007/978-3-319-44321-8_49. (anglicky)
- Der Pfeilstorch in der Zoologischen Sammlung [online]. University of Rostock, 2003 [cit. 2021-12-10]. S. 9–10. (německy)
- Instructions for observing nocturnal bird migration by moon-watching [online]. University of Arizona, 2004 [cit. 2021-12-29]. Dostupné online. (anglicky)
- PREUSS, Niels Otto. Hans Christian Cornelius Mortensen: Aspects of his life and of the history of bird ringing [online]. Duglebjerg, Denmark: 2000 [cit. 2021-12-11]. Dostupné online. (anglicky)
- OVČIARIKOVÁ, S.. Metody sledování migrace zvířat se zaměřením na telemetrii. , 2020. . FAKULTA VETERINÁRNÍ HYGIENY A EKOLOGIE: Ústav biologie a chorob volně žijících zvířat. . Dostupné online.
- KLVAŇA, Petr; CEPÁK, Jaroslav. Kudy a kam ptáci létají. Vesmir.cz [online]. 2018 [cit. 2021-12-29]. Dostupné online.
- Frequently Asked Questions about GPS Transmitter (NC99) [online]. The Wildlife Center of Virginia [cit. 2022-01-28]. Dostupné online. (anglicky)
- BREZINA, Ivan. Migrující ptáci chytře využívají každou příležitost k přežití. I kosi. iDNES.cz [online]. 2019-03-22 [cit. 2021-12-29]. Dostupné online.
- HALLWORTH, Michael T.; MARRA, Peter P. Miniaturized GPS Tags Identify Non-breeding Territories of a Small Breeding Migratory Songbird. Scientific Reports. 2015-06-09, roč. 5, čís. 1, s. 11069. Dostupné online [cit. 2022-01-27]. ISSN 2045-2322. DOI 10.1038/srep11069. PMID 26057892. (anglicky)
- NILSSON, Cecilia; DOKTER, Adriaan M.; SCHMID, Baptiste; SCACCO, Martina; VERLINDEN, Liesbeth; BÄCKMAN, Johan; HAASE, Günther. Field validation of radar systems for monitoring bird migration. S. 2552–2564. Journal of Applied Ecology [online]. 2018 [cit. 2021-12-29]. Roč. 55, čís. 6, s. 2552–2564. DOI 10.1111/1365-2664.13174. (anglicky)
- BORGMANN, Kathi. Ornithology lab releases high-resolution migration maps [online]. Cornell University, 2020-03-03 [cit. 2021-12-29]. Dostupné online. (anglicky)
- JOHNSON, Michael. International Space Station Images Trace Bird Migrations [online]. NASA, 2021-03-01 [cit. 2021-12-29]. Dostupné online. (anglicky)
- EMLEN, Stephen T.; EMLEN, John T. A Technique for Recording Migratory Orientation of Captive Birds. The Auk. 1966-07-01, roč. 83, čís. 3, s. 361–367. Dostupné online [cit. 2021-12-29]. ISSN 1938-4254. DOI 10.2307/4083048. (anglicky)
- PROCHÁZKA, Petr. Analýza stabilních izotopů – alternativní metoda studia migrace ptáků. S. 3–21. Sylvia [online]. ČSO, 2006. Roč. 42, s. 3–21. Dostupné online.
- DUFKOVÁ, Marie. Medicína a přírodověda [online]. Třípól, 2021-05-31 [cit. 2021-01-18]. Dostupné online.
- Dohoda o ochraně africko-euroasijských stěhovavých vodních ptáků (AEWA) [online]. Ministerstvo životního prostředí [cit. 2021-12-30]. Dostupné online.
- Prioritizing Stopover Habitats [online]. Conservation Gateway: The Nature Conservancy [cit. 2021-12-30]. Dostupné online.
- What we are doing for Europe's migratory birds [online]. Euronatur.org [cit. 2021-12-30]. Dostupné online.
- Wisconsin Whooping Crane Management Plan [online]. Wisconsin Department of Natural Resources, 2006 [cit. 2021-12-22]. Dostupné online. (anglicky)
- The Passenger Pigeon. Smithsonian Institution [online]. [cit. 2021-12-30]. Dostupné online. (anglicky)
- Critically Endangered Migratory Birds. www.worldmigratorybirdday.org [online]. [cit. 2021-12-30]. Dostupné online.
- World Migratory Bird Day [online]. [cit. 2021-12-30]. Dostupné online. (anglicky)
- HORTON, Kyle G.; LA SORTE, Frank A.; SHELDON, Daniel. Phenology of nocturnal avian migration has shifted at the continental scale. Nature Climate Change. 2020-01, roč. 10, čís. 1, s. 63–68. Dostupné online [cit. 2021-12-28]. ISSN 1758-6798. DOI 10.1038/s41558-019-0648-9. (anglicky)
- JENNI, Lukas; KÉRY, Marc. Timing of autumn bird migration under climate change: advances in long–distance migrants, delays in short–distance migrants. Proceedings of the Royal Society of London. Series B: Biological Sciences. 2003-07-22, roč. 270, čís. 1523, s. 1467–1471. Dostupné online [cit. 2021-12-28]. ISSN 0962-8452. DOI 10.1098/rspb.2003.2394. PMID 12965011. (anglicky)
- ORELLANA-MACÍAS, José; BAUTISTA, Luis; MERCHÁN, Daniel. Shifts in crane migration phenology associated with climate change in southwestern Europe. Avian Conservation and Ecology. 2020-05-08, roč. 15, čís. 1. Dostupné online [cit. 2021-12-28]. ISSN 1712-6568. DOI 10.5751/ACE-01565-150116. (anglicky)
- Climate change may be why birds are migrating earlier across the United States [online]. 2019-12-17 [cit. 2022-01-18]. Dostupné online. (anglicky)
- DUNN, Peter O; WINKLER, David W. Climate change has affected the breeding date of tree swallows throughout North America. Proceedings of the Royal Society of London. Series B: Biological Sciences. 1999-12-22, roč. 266, čís. 1437, s. 2487–2490. Dostupné online [cit. 2022-01-18]. DOI 10.1098/rspb.1999.0950. PMID 10693819. (anglicky)
- BOTH, Christiaan; BOUWHUIS, Sandra; LESSELLS, C. M. Climate change and population declines in a long-distance migratory bird. Nature. 2006-05, roč. 441, čís. 7089, s. 81–83. Dostupné online [cit. 2021-12-28]. ISSN 0028-0836. DOI 10.1038/nature04539. (anglicky)
- LAWRENCE, Kieran B.; BARLOW, Clive R.; BENSUSAN, Keith. Phenological trends in the pre- and post-breeding migration of long-distance migratory birds. Global Change Biology. 2022, roč. 28, čís. 2, s. 375–389. Dostupné online [cit. 2022-01-27]. ISSN 1365-2486. DOI 10.1111/gcb.15916. (anglicky)
- Tažní ptáci tráví v Evropě více času. Někteří mohou přestat migrovat, tvrdí studie - Novinky.cz. www.novinky.cz [online]. [cit. 2022-01-27]. Dostupné online.
- VISSER, Marcel E.; PERDECK, Albert C.; BALEN, JOHAN H. Van. Climate change leads to decreasing bird migration distances. Global Change Biology. 2009, roč. 15, čís. 8, s. 1859–1865. Dostupné online [cit. 2022-01-18]. ISSN 1365-2486. DOI 10.1111/j.1365-2486.2009.01865.x. (anglicky)
- ZIMOVA, Marketa; WILLARD, David E.; WINGER, Benjamin M. Widespread shifts in bird migration phenology are decoupled from parallel shifts in morphology. Journal of Animal Ecology. 2021, roč. 90, čís. 10, s. 2348–2361. Dostupné online [cit. 2021-12-28]. ISSN 1365-2656. DOI 10.1111/1365-2656.13543. (anglicky)
- Smaller bodies, longer wings, earlier migrations: Untangling the multiple impacts of climate warming. ScienceDaily [online]. [cit. 2022-01-18]. Dostupné online. (anglicky)
- PANUCCIO, Michele; MARTÍN, Beatriz; MORGANTI, Michelangelo; ONRUBIA, Alejandro; FERRER, Miguel. Long-term changes in autumn migration dates at the Strait of Gibraltar reflect population trends of soaring birds. S. 55–65. Ibis [online]. 2017-01 [cit. 2021-12-15]. Roč. 159, čís. 1, s. 55–65. DOI 10.1111/ibi.12420. (anglicky)
- LA SORTE, Frank A.; HOCHACHKA, Wesley M.; FARNSWORTH, Andrew; DHONDT, André A.; SHELDON, Daniel. The implications of mid‐latitude climate extremes for North American migratory bird populations. Ecosphere [online]. 2016-03. Roč. 7, čís. 3. DOI 10.1002/ecs2.1261. (anglicky)
- Migratory Birds in Culture [online]. World Migratory Birds Day (UNEP/CMS) [cit. 2022-02-19]. Dostupné online. (anglicky)
- SAX, Boria. Bird Migration and Human Culture [online]. [cit. 2022-02-19]. Dostupné online. (anglicky)
- Jiřičky stavějící si hnízdo [online]. Česká společnost ornitologická [cit. 2022-02-19]. Dostupné online.
- Ne všichni čápi odlétají do Afriky [online]. Česká společnost ornitologická, 2017-12-12 [cit. 2022-02-19]. Dostupné online.
- Čáp bílý – důstojný pták opředený mnoha pověrami [online]. Mikroregion Slezská Harta [cit. 2022-02-19]. Dostupné online.
- Podivuhodná cesta Nilse Holgerssona Švédskem [online]. Databazeknih.cz [cit. 2022-02-19]. Dostupné online.
- Ptačí svět (2001) [online]. CSFD.cz [cit. 2022-02-19]. Dostupné online.
- Migrations secrètes: Odyssées aériennes des animaux migrateurs [online]. IMDB.ecom [cit. 2022-02-19]. Dostupné online. (anglicky)
- Cesta domů (1996) [online]. CSFD.cz [cit. 2022-02-19]. Dostupné online.
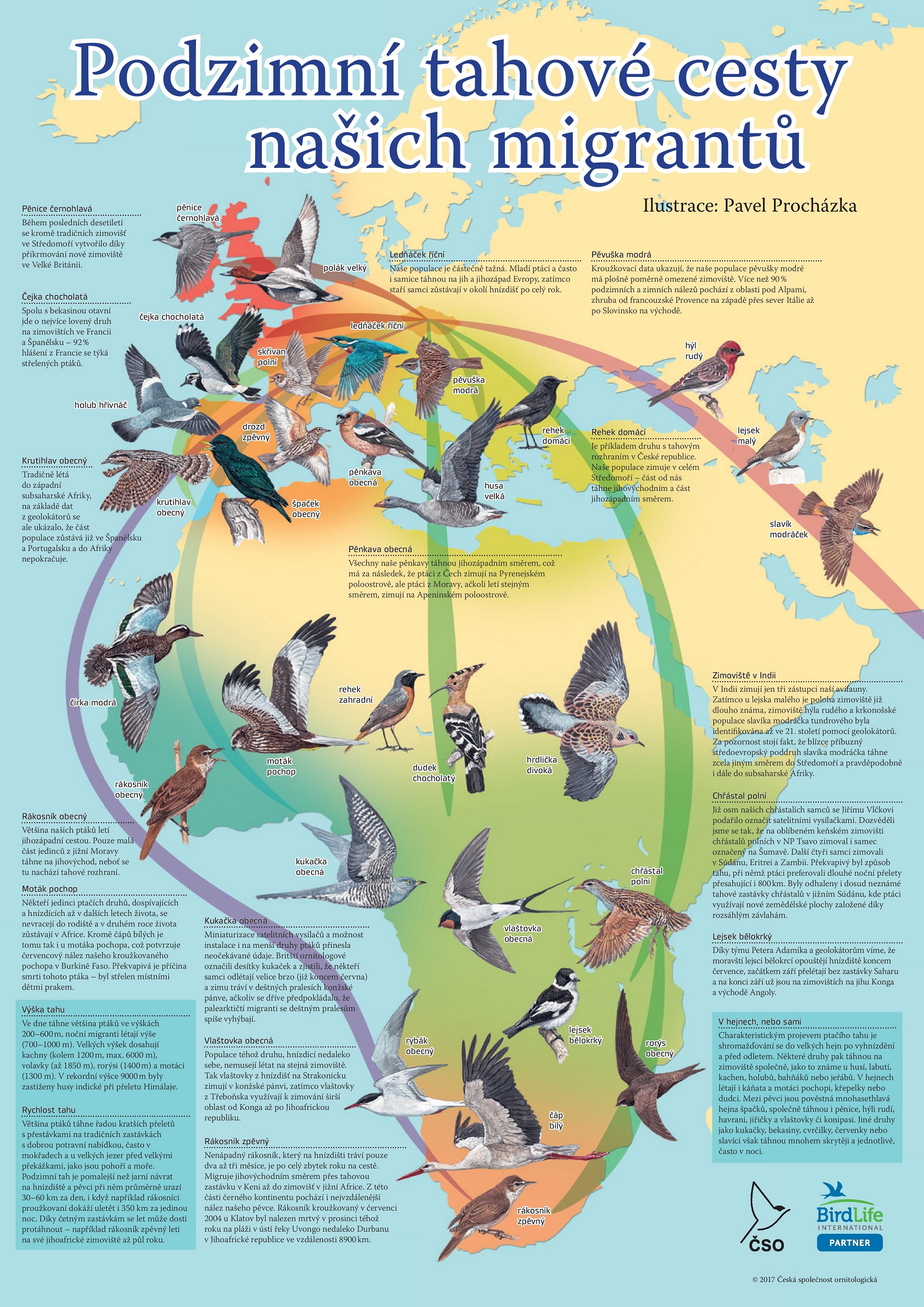{kind=link}
Literatura
- česky
- CEPÁK, Jaroslav, et al., 2008. Atlas migrace ptáků České a Slovenské republiky = Czech and Slovak bird migration atlas. Praha: Aventinum. ISBN 978-808-6858-876.
- CLOUDSLEY-THOMPSON, John, 1988. Migrace zvířat. Praha: Albatros.
- ČERNÝ, Walter, 1980. Ptáci. Praha: Artia.
- VESELOVSKÝ, Zdeněk. Obecná ornitologie. Praha: Academia, 2001. ISBN 8020008578.
- anglicky
- BERTHOLD, P., 2001. Bird migration: a general survey. Oxford: Oxford University Press. ISBN 9780198507871. (anglicky)
- LINCOLN, Frederick C., 1935. Migration of Birds: Circular 16. Manhattan, Kansas, US: U.S. Fish and Wildlife Service. Dostupné online. (anglicky)
- NEWTON, Ian, 2008. The migration ecology of birds. Amsterdam: Elsevier-Academic Press. ISBN 978-0-12-517367-4. (anglicky)
- RAPPOLE, John H., 2013. The avian migrant: the biology of bird migration. New York: Columbia University Press. ISBN 978-0-231-14678-4. (anglicky)
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu Migrace ptáků na Wikimedia Commons
Obrázky, zvuky či videa k tématu Migrace ptáků na Wikimedia Commons - BirdCast: Migration tools (anglicky)
- Bird Migration (anglicky) – dokument o migraci ptáků přes severozápadní Anglii
- Vrcholí migrace ptáků – pomozme jim v letu o život! – článek od ČSO