Eukaryota
Eukaryota (též Eukarya či česky jaderní) je doména, kam patří jednak veškeré známé mnohobuněčné formy života[pozn. 1], jako jsou živočichové[pozn. 2], rostliny[pozn. 3], houby nebo stramenopilní řasy (chaluhy, řasovky), a všechny jednobuněčné organismy řazené mezi protisty, které jsou co do počtu vývojových linií mezi eukaryoty ještě mnohem bohatší a různorodější. Eukaryotická buňka na rozdíl od prokaryotní obsahuje pravé buněčné jádro a množství dalších organel oddělených membránou od okolí. Tyto struktury rozdělují buňku na mnoho menších oddílů (kompartmentů). Eukaryontní organismy mají množství jiných unikátních znaků, jako je například specifická struktura bičíků a řasinek, existence cytoskeletu, určité odchylky ve struktuře genomu (např. DNA rozdělená do jednotlivých chromozomů, přítomnost intronů ve větším množství), schopnost pohlavního rozmnožování a mnohá další metabolická a biochemická specifika.
 | |
| Vědecká klasifikace | |
| Doména | Eukaryota Whittaker & Margulis, 1978 |
| Superskupiny | |
| |
| Sesterská skupina | |
| Archea: Heimdallarchaeota | |
| Některá data mohou pocházet z datové položky. | |
K eukaryontním organismům se řadí všechny buněčné organismy vyjma bakterií a archeí, tedy prokaryot.[pozn. 4] Klasifikace eukaryot se v poslední době radikálně mění. Tradiční systém říší rostliny, houby, živočichové a protista (poslední jmenovaná zahrnovala veškeré ostatní eukaryotické organismy) byl v současnosti překonán konceptem fylogeneticky přirozených superskupin. K roku 2020 je vymezeno sedm superskupin, Amorphea, Archaeplastida, CRuMs, TSAR, Cryptista, Haptista a Hemimastigophora, kromě kterých existuje spousta dosud nezařazených taxonů, včetně zástupců bývalé říše Excavata.[2] Odhady týkající se počtu druhů eukaryot se pohybují mezi 5 a 30 miliony,[3][4] popsáno bylo méně než dva miliony druhů.[4]
Vznik a vývoj
Datace a fosilní nálezy

Vznik eukaryotických buněk (a potažmo tedy eukaryot) je významný milník v evoluční historii života, je však zahalen nejistotou. Někdy se datuje do doby před 1,8 – 1,3 miliardami lety,[5] tedy o mnoho dříve před kambrijskou a ediakarskou diverzifikací života. Jindy se dokonce uvádí doba před dvěma miliardami let[7] či více.[8] Z těchto období naší geologické historie je k dispozici jen málo fosilních nálezů eukaryot, i když výjimky existují: byly popsány starohorní fosilní nálezy protistů, které jsou označovány za časné zástupce eukaryot.[5] Také některé nálezy řas (Grypania) jsou podle jejich objevitelů až 2,1 miliardy let staré.[6] Stopy po existenci eukaryot byly dokonce nalezeny i v australských břidlicích starých 2,7 miliardy let.[9] I přes tyto nálezy se hlavní současné linie eukaryot pravděpodobně oddělily až ke konci starohor, konkrétně v období mezi 1,3 – 0,72 miliardami let. Fosílie časných eukaryot, jako je například Shuiyousphaeridium macroreticulatum, může být obtížné odlišit od prokaryotických buněk. U tohoto domnělého eukaryota se při jeho řazení například vycházelo z velkých rozměrů buňky, pevné stěny a výrůstků na jeho povrchu.[5] Většinu nálezů ze starohor však nejsou vědci schopni zařadit do konkrétnější skupiny eukaryot. Kvalitní fosílie eukaryot se začínají objevovat až v ediakaru a kambriu, tedy v období asi před půl miliardou let. V ediakaru byly nalezeny vyjma zelených řas také zvláštní planktonní organismy ze skupiny Acritarcha.[10]
Vznik eukaryotické buňky
Otázka vzniku eukaryotické buňky (eukaryogeneze), která je charakteristická množstvím vnitřních membrán a složitými organelami (mitochondrie), není zcela vyjasněna. Dnes se zdá, že ke vzniku eukaryot přispěly jak archeální, tak bakteriální genomy.[11] Jedním ze světlých míst je vznik mitochondrií. V této organele byla totiž nalezena DNA, která vykazuje značnou příbuznost s bakteriemi z řádu Rickettsiales. Endosymbiotická teorie tak oprávněně tvrdí, že mitochondrie vznikly právě pohlcením Rickettsií jinou buňkou a mitochondrie jsou potomci těchto bakterií.[11]
To však neznamená, že pouhý vznik mitochondrií znamenal vznik eukaryot v dnešním slova smyslu. Eukaryogenezi se snaží dále vysvětlit více než 20 hypotéz.[12] Podle archeální hypotézy vznikla protoeukaryotická buňka z buňky jisté archebakterie a mitochondrie se vyvinuly až za určitý čas, a to pohlcením bakteriálního symbionta.[11] Detailněji se okolnostmi archeální hypotézy zabývá tzv. vodíková hypotéza, podle níž zmíněná archebakterie metabolizovala vodík, který ji právě poskytovaly v rámci své respirace endosymbiotické bakterie. Tyto bakterie následně prošly evolucí a změnily se na mitochondrie.[13]
Jiné studie však navrhují o krok složitější způsob vzniku eukaryotické buňky. Takzvané chimérické (čili fúzní) hypotézy předpokládají, že amitochondriální eukaryot (tedy předek eukaryot ještě bez mitochondrií) vznikl splynutím jedné buňky archeální a jedné bakteriální. Eukaryotický genom je podle této hypotézy rovněž mixem archeálních a bakteriálních genů. Až následně vznikly mitochondrie (a plastidy), a to dalšími endosymbiotickými událostmi.[14] Podobná teorie navrhuje jiný sled událostí: archebakterie strávila bakterii jako potravu a část bakteriální DNA se včlenila do archeální DNA.[15] Zcela jiný pohled nabízí teorie „Neomura“, která považuje archea i eukaryota za potomky určité složité grampozitivní bakterie, konkrétně jisté aktinobakterie. Tato aktinobakterie prošla značným vývojem a až později endosymbioticky získala mitochondrie.[16] Nedostatkem těchto teorií je fakt, že v současnosti neznáme žádné eukaryotické organismy s primární absencí mitochondrií, ačkoliv je tyto teorie předpokládají.[11]
Mnohobuněčnost


U eukaryot mnohobuněčnost vznikla několikrát nezávisle na sobě, zvlášť u živočichů, rostlin, hub a mnoha dalších eukaryotických taxonů.[18] Vznik (respektive vzniky) mnohobuněčnosti byl značným evolučním úspěchem eukaryot, u něhož byla například vyřešena otázka, jak se buňky navzájem dorozumí a jak si jednotlivé buněčné typy rozdělí práci.[19] U mnohobuněčných živočichů jsou unikátní nejen geny zajišťující správný embryonální vývoj (např. Hox geny), ale i geny zajišťující komunikaci mezi buňkami. U mnohobuněčných se vyskytují i nové buněčné struktury (dezmozomy a jiné buněčné spoje).[20]
První mnohobuněční živočichové vznikli v průběhu období ediakara (někdy před 570 – 550 miliony lety),[20] jak napovídají i některá molekulárně biologická data. Zřetelnější fosilní záznam se však objevuje až v kambriu. Z této doby, označované také jako kambrická exploze druhů, pochází množství fosilních dokladů současných živočišných kmenů.[21][22] U rostlin (včetně mnohých ruduch a zelených řas) se mnohobuněčnost vyvinula hned několikrát, a to zřejmě v souvislosti s tím, že se fotosyntetizující organismy nemusí tolik pohybovat za potravou, a vznik mnohobuněčnosti je pro ně tudíž schůdnější.[23] Vývoj mnohobuněčného uspořádání se u rostlin ubíral třemi základními cestami. Některé kolonie zelených řas, jaké tvoří např. váleč koulivý (Volvox globator), se zformovaly z několika původních jedinců. Jindy se v původní buňce mnohonásobně dělilo jádro, čímž došlo ke vzniku jednobuněčných, ale vícejaderných stélek (např. rod Caulerpa). Konečně třetím způsobem vznikly mnohobuněčné rostliny tak, že se původní buňka mitoticky dělila a jednotlivé dceřiné buňky se specializovaly na určitou funkci.[24] U hub nelze pozorovat nějaký obecný trend k mnohobuněčnosti a není vlastně jasné, kolikrát v rámci hub mnohobuněčnost vznikla.[20]
Skupin, u nichž byla pozorována mnohobuněčnost, je však mnohem více. Příkladem jsou akrasie (Acrasidae), chaluhy (Phaeophyceae), řasovky (Oomycota), pravé hlenky (Eumycetozoa) a dokonce několik nálevníků (Ciliophora) tvořících plodničky.[18][23]
Eukaryotická buňka
Velikost
Buňky eukaryot mají v průměru desetkrát větší rozměry (lineární) než buňky prokaryotických organismů,[25] ačkoliv toto pravidlo platí jen zhruba. Nejmenší eukaryotické buňky má zelená řasa Ostreococcus tauri, a to přibližně jeden μm,[26] tzn. menší než např. buňka bakterie Escherichia coli.[27] Naopak známe mnohé obrovské eukaryotické buňky. Značných rozměrů dosahují zejména některé mnohojaderné buňky uvnitř těl velkých živočichů (jako např. některé nervové buňky), které jsou však neschopné samostatné existence. Také žloutek ve vejcích pštrose či vyhynulého ptáka Aepyornis dosahuje obrovských rozměrů, ačkoliv před oplozením představuje též jen jedinou buňku.[28] Pokud se však zaměříme na největší buňku schopnou samostatné existence, existují i mimořádně velké jednobuněčné organismy. Velmi velká (až jeden metr) je jednobuněčná zelená řasa rodu Caulerpa.[29] Mnohojaderná plazmodia prvoků, jako je Physarum polycephalum, mohou také dosahovat velikosti několika metrů – zaznamenáni byli jedinci uvnitř těl velkých živočichů s plochou 5,54 m2.[30]
Stavba buňky

Buňka eukaryot je radikálně odlišná od prokaryotické buňky nejen v otázce velikosti, ale vykazuje značné rozdíly i po stránce strukturní. Na rozdíl od prokaryot jsou komplexnější a tzv. kompartmentalizované, tedy rozdělené na jednotlivé kompartmenty („oddělení“). Obsahují například pravé jádro obklopené dvojitou membránou, jež odděluje DNA od okolních částí buňky. Dalším výrazným rysem je endomembránový systém, tedy skupina organel rovněž obalených membránou. K těmto patří zejména endoplazmatické retikulum, Golgiho aparát, lyzozomy či mitochondrie, případně funkční deriváty mitochondrií (hydrogenozomy, mitozomy). Evoluční novinkou je na rozdíl od prokaryot i velmi rozvinutý cytoskelet, jakási vnitřní kostra buňky tvořená především aktinem a tubulinem. Ten zajišťuje nejen pohyb buněk, ale i vnitřní transport a zastává též důležité funkce při dělení buněk. Neméně důležitou součástí jsou ribozomy, u eukaryot jsou tzv. 80S (toto číslo udává čas, za který proběhne sedimentace ribozomální makromolekuly při její ultracentrifugaci).
Mimo výše uvedené struktury, které jsou přítomné téměř ve všech eukaryotických buňkách, se v mnoha skupinách eukaryot vyvinuly i jiné specifické organely. Typickým příkladem jsou plastidy, semiautonomní organely, v nichž probíhá fotosyntéza a některé další pochody. Vyskytují se u rostlin a mnohých protistů. Vznikly pravděpodobně v evoluční historii až určitou dobu po vzniku samotné eukaryotické buňky a samotné eukaryogeneze se neúčastnily. Přesto jsou však dalším dokladem endosymbiotické teorie.[31] Na rozdíl od mitochondrií se plastidy vyvinuly u několika, často nepříbuzných skupin eukaryot. Tzv. primární plastidy však pochází ze sinic a vznikly pravděpodobně pouze jednou, a to u rostlin, které v širším pojetí zahrnují nejen zelené rostliny (Viridiplantae), ale i ruduchy (Rhodophyta) a glaukofyty (Glaucophyta).[32] Plastidy u jiných druhů fotosyntetizujících eukaryot vznikly především sekundární endosymbiózou, tedy pohlcením jedné ze skupin rostlin. Pohlcením ruduchy vznikly plastidy (někdy kvůli svému původu zvané rhodoplasty) např. u různých heterokont (Heterokonta) a rozsivek (Dinoflagellata), pohlcením zelené řasy vznikly plastidy u Chlorarachniophyta, některých krásnooček (Euglenoidea) a jedné obrněnky.[31]
Zvláště u různých jednobuněčných protistů je známo mnoho dalších unikátních organel: vystřelovací či vymršťovací útvary (tzv. extruzomy), přichycovací organely (různé stonky a přísavky), stažitelné, potravní a jiné vakuoly, a podobně. K pohybu zase slouží různé bičíky a brvy (viz kapitola pohyb). Zcela samostatnou kapitolou je anatomie mnohobuněčných organismů (viz anatomie rostlin, anatomie živočichů, anatomie hub).
Klasifikace
Historická klasifikace
| Linnaeus 1735 2 říše |
Haeckel 1866[33] 3 říše |
Chatton 1937[34] 2 říše |
Copeland 1956[35] 4 říše |
Whittaker 1969[36] 5 říší |
Woese et al. 1977[37] 6 říší |
Woese et al. 1990[38] 3 domény |
|---|---|---|---|---|---|---|
| Protista | Prokaryota | Monera | Monera | Eubacteria | Bacteria | |
| Archaebacteria | Archaea | |||||
| Eukaryota | Protista | Protista | Protista | Eukarya | ||
| Vegetabilia | Plantae | Fungi | Fungi | |||
| Plantae | Plantae | Plantae | ||||
| Animalia | Animalia | Animalia | Animalia | Animalia |
Historické snahy o klasifikaci živých organismů jsou z velké části právě klasifikací eukaryot. Na nejvyšší úrovni v průběhu času docházelo k velkým změnám. Zatímco Linné rozeznával jen dvě základní říše, rostliny a živočichové, postupně se ukázalo, že jsou si tyto skupiny (a dále někteří jednobuněční a houby) v mnoha rysech vzájemně podobné a zároveň diametrálně odlišné od bakterií. Pravděpodobně prvním, kdo si toto uvědomil, byl francouzský biolog Edouard Chatton. Ten v roce 1937 navrhl dělit všechny živé organismy na dvě říše, eukaryota a prokaryota. Jeho návrh však poněkud zapadl a znovu se k němu v roce 1962 vrátili Roger Stanier a Cornelius Van Niel. Ačkoliv je v současnosti taxon prokaryota považován za zastaralý a nerespektující fylogenetický vývoj, taxon eukaryota se běžně používá.[39] Podpořili ho v roce 1990 Carl Woese, Otto Kandler a Mark Wheelis, kteří rozčlenili veškerý život do tří domén, Archaea, Bacteria a Eucarya.[40]
Tradičně se eukaryota dělila na několik říší, například podle Whittakera[41] na říše živočichové (Animalia), rostliny (Plantae), houby (Fungi) a protisté (Protista). Cavalier-Smith dále rozdělil říši protistů na dvě dílčí, a to protozoa a chromista.[42] Dnes se však prosazují zcela jinak postavené systémy.[43]
Současný taxonomický přístup

Většina dnešních systémů uznává třídoménový systém z roku 1990 a s ním spojenou existenci domény Eukaryota.[44][45] Nejnovější poznatky dokonce ukazují, že Eukaryota jsou podle jaderného genomu ve skutečnosti vnitřní skupinou archeí, která se odvětvuje uvnitř kladu Asgard,[46][47] pravděpodobně jako sesterská k nově popsané skupině Heimdallarchaeota.[48][49][pozn. 6] Domény tak vlastně mohou být pouze dvě a arachea bez eukaryot by tak představovala parafylum.
Klasifikace této domény na nižších úrovních byla dlouho nejasná, avšak dnes je již systém říší, jako např, prvoci (Protozoa), vesměs překonán konceptem takzvaných superskupin ("supergroups").
Přístup založený na tradičních taxonech
Velká část klasifikačních systémů nadále spoléhá, v souladu s názvoslovnými normami, na tradiční taxonomické úrovně, přičemž se je snaží naplnit co nejpřirozeněji s ohledem na skutečnou příbuznost. Omezené množství klasifikačních úrovní však neumožňuje vždy vyjádřit podřízenost, a tak přetrvávají i taxony, kladoucí na stejnou úroveň více podřízených taxonů, než pouhé sesterské skupiny a často obsahují parafyletické, případně i polyfyletické taxony. Naopak se v těchto systémech zpravidla daří na některou úroveň umístit i skupiny nevyjasněného postavení, tzv. incertae sedis a skupiny vyhynulé. Využívají zpravidla tradičně zavedené říše, třebaže si nové poznatky vynutily jejich částečné přeskupení a pozměněné naplnění.
Systémy založené na tomto tradičním přístupu jsou vhodné pro katalogizaci organismů a proto se používají v mnohých internetových katalozích, jako Catalogue of Life, Species 2000, AlgaeBase, WoRMS apod. Z pedagogických důvodů jsou vhodné pro podání základního přehledu o eukaryotních organismech, naopak nevhodné pro problematiku evoluce a vzájemné příbuznosti.
Příklad ukazuje následující tabulka (podbarvení říší odpovídá podbarvení taxoboxů):
Fylogenetický přístup
- Podrobnější informace naleznete v článku Klasifikace eukaryot, oddílu Fylogenetické stromy.
Jiné klasifikační systémy využívají důsledněji fylogenetický přístup a jsou proto strukturovanější a lépe respektují přirozenost (monofylii) jednotlivých skupin a kladogramy získané z fylogenetických analýz – protože zpravidla nepoužívají tradiční taxony, mají možnost vyjádřit do veliké hloubky vzájemné příbuzenské vztahy. I z tohoto důvodu však oplývají velkým množstvím incertae sedis na všech úrovních a vzhledem k dosud nedostatečnému pokrytí molekulárně genetickými daty si mnohý nový poznatek vyžádá zásadnější revizi systému.
Konkrétně u eukaryot se při fylogenetickém přístupu podařilo nastolit i vyvrátit mnoho hypotéz o příbuznosti a přirozenosti vyšších skupin, což bylo zpravidla spojeno se změnou struktury systému. Lze uvést mnoho příkladů:[52]
- Byly vyvráceny původní představy o vymezení skupin Chromista a Chromalveolata (tehdy bez Rhizaria) a požadavek přirozenosti vedl nejprve ke sjednocení jejich obsahu (zahrnutí rhizarií), ale nakonec i ke zpochybnění celkové přirozenosti.[53][54] Naopak přibývající data dosud podporují přirozenost jejich základních skupin: Sar,[55] Haptista a Cryptista[52] (ale ne jako součásti dříve předpokládané společné přirozené skupiny Hacrobia[56]).
- Byly nastoleny buněčně strukturní základy pro vymezení skupin Unikonta a Bikonta,[55] poté byly vyvráceny a nově nastoleno základní členění neexkavátních eukaryot na superskupiny Diaphoretickes a Amorphea[57]
- Protože uvnitř exkavát se podle současných představ nachází kořen eukaryotického fylogenetického stromu, nelze je chápat jako přirozenou skupinu,[52] naopak to lze tvrdit o jejich základních částech: Discoba[58] a Metamonada.
- Na vývojové linii Amorphea byly potvrzeny jako přirozené větve měňavkovci (Amoebozoa) a Obazoa a byly identifikovány jednotlivé bazální linie odvětvující se před vlastním korunovým kladem živočichů (dohromady tvoří Holozoa) jakož i bazální linie odvětvující se před vlastním korunovým kladem hub (dohromady tvoří Holomycota).
Klasifikace důsledně založená na fylogenetickém přístupu (v nejpřísnější podobě je to kladistika) je používána ve většině odborných článků zabývajících se příbuzenskými vztahy organismů, jejich evolucí a fylogenetickou klasifikací; pro objasňování této problematiky je vhodná i z pedagogických důvodů, nehodí se však pro představení základního přehledu eukaryotních organismů.
Takto založené klasifikační systémy jsou zpravidla v textu prezentovány odrážkovou strukturou s proměnným odsazením, kde nejvyšší úroveň představují tzv. superskupiny, ale nejčastěji graficky pomocí fylogenetických stromů.
Současné (2020) představy o fylogenezi eukaryot pracují zpravidla s následujícími 7 superskupinami (bez ohledu na jejich početnost - důležité je fylogenetické postavení blízko předpokládaného kořene fylogenetického stromu eukaryot):[52][59][60]
- Amorphea – druhově nejpočetnější superskupina: houby, živočichové, trubénky, měňavkovci;
- CRuMs – nepočetná skupina drobných protist, fylogeneticky nejbližší ke skupině Amorphea;
- Hemimastigophora – nepočetná skupina bičíkatých predátorů (hluboká linie eukaryot);
- Archaeplastida – rostliny (zelené rostliny, ruduchy, Rhodelphidia a glaukofyty);
- Cryptista – rozšířené skrytěnky, nejspíš blízké k rostlinám;
- Haptista – rozšířená Haptophyta, nejspíš blízké k TSAR;
- TSAR – původní superskupina Sar rozšířená o Telonemia;
Zjednodušený fylogenetický strom eukaryot podle současných (tj. počátek r. 2019)[52] představ (záměrně není vyznačen kořen stromu, jehož poloha není dosud definitivně vyjasněna):
| Diaphoretickes |
| ||||||||||||||||||||||||||||||||||||||||||||||||
|
Discoba (krásnoočka, améboflageláti…) | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
Genetika
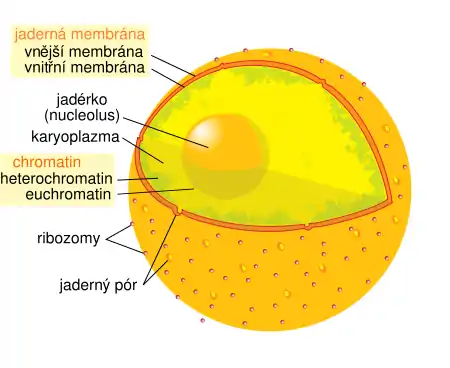
Genetický materiál v podobě DNA je uložen v jádře (či několika jádrech, jako u nálevníků) a v semiautonomních organelách, jako jsou mitochondrie a plastidy (vzácně též jinde, např. v nukleomorfu, zbytku jádra po endosymbiontovi). Tato DNA se následně, podobně jako u ostatních domén organismů, přepisuje do RNA a ta pak slouží buď k syntéze proteinů (mRNA), nebo má určitou katalytickou funkci jako taková (rRNA, tRNA a podobně).
V jádře tvoří DNA a některé proteiny (např. histony) tzv. chromatin, jehož jednotlivé oddělené části se nazývají chromozomy. Stáčením (spiralizací) a rozplétáním (despiralizací) chromatinu se značně mění jejich velikost. Velikost jaderného genomu (počet bází) je značně rozmanitá. Nejmenší eukaryotický genom, pouze 2,9 milionu párů bází, má vnitrobuněčná parazitická mikrosporidie jménem Encephalitozoon cuniculi,[61][62] a to proto, že u ní došlo k redukci genomu v rámci jejího parazitického způsobu života. Naopak největší genom ze všech známých eukaryot (670 miliard párů bází) má Amoeba dubia.[63] Oproti prokaryotním organizmům u eukaryot většina jejich DNA nekóduje žádný gen (a tedy ani protein). U lidí tvoří nekódující DNA asi 97 % genomu a její funkce, pokud nějaké vůbec má, není v drtivé většině případů dosud známa. Označuje se též jako junk DNA, související termíny jsou také introny či repetitivní DNA.[24]
Mitochondriální DNA a plastidová DNA jsou oproti jaderné DNA relativně malé a cirkulární a považují se za pozůstatek po endosymbiotických událostech, při nichž byl eukaryontní buňkou pohlcen prokaryotní organismus. Mitochondriální DNA má mnoho společných znaků s DNA alfa proteobakterií, plastidová vykazuje příbuznost s DNA cyanobakterií (sinic).
Životní projevy
Metabolismus
Metabolické pochody výrazně ovlivňuje už vyšší velikost eukaryontní buňky, která značně zvyšuje poměr objemu a plochy buňky.[24] V podstatě rozeznáváme u eukaryot dva druhy výživy: obligátní fotoautotrofii u rostlin a jiných fotosyntetizujících organismů a obligátní chemoheterotrofii u živočichů, hub a jiných organismů živících se organickým materiálem.[64] Pokud umí nějaký organismus užívat oba tyto základní typy výživy (např. mnohá krásnoočka), označuje se termínem mixotrof. Najdeme však drobné odchylky od klasických způsobů získávání živin a energie. Někteří prvoci a houby například dokáží metabolizovat i bez kyslíku.[65]
Rozmnožování
V reprodukci eukaryot hraje velkou roli nepohlavní rozmnožování, ale evolučně unikátní je zřejmě schopnost rozmnožovat se pohlavně. Poměrně často dochází k pravidelnému střídání pohlavního a nepohlavního rozmnožování (metageneze). Pouze u některých protistů, jako trubénky (Choanoflagellata), eugleny (Euglenoidea), trichomonády (Trichomonadida) a většina krytének (Arcellinida), nebyla sexuální reprodukce dosud nalezena a o její existenci můžeme jen spekulovat.[30]
Nepohlavně se množí většina protistů, ale i některé mnohobuněčné organismy. U jednobuněčných organismů je rozmnožování asociováno s mitózou. Před vlastní cytokinezí musí být replikována DNA a také se musí zmnožit organely, aby každá dceřiná buňka byla kompletní. Rozlišuje se několik typů nepohlavního rozmnožování, a to prosté dělení ve dva (u protistů nejběžnější), dále pučení (např. rournatky, Suctoria) či mnohonásobné dělení buňky za vzniku spousty dceřiných buněk (různí výtrusovci, Apicomplexa).[30] U mnohobuněčných organismů se vyskytuje široké spektrum různých druhů nepohlavního rozmnožování, jako je vegetativní rozmnožování (např. fragmentace u mnohých rostlin), tvorba spor nepohlavní cestou, partenogeneze, apomixie či též pučení.
I pohlavní rozmnožování je v rámci eukaryotické domény neobyčejně rozšířené (až na výše zmíněné výjimky). Typický scénář u mnoha prvoků i mnohobuněčných organismů vypadá takto: v procesu meiózy vznikají dva typy sexuálně diferencovaných buněk, tzv. gamety, obvykle označované jako + a – (či samčí a samičí). Pokud tyto buňky vypadají velmi podobně, pak rozmnožování je izogamické, pokud jsou odlišné, pak se mluví o anizogamii. Specifickým typem je pak oogamie, při níž je velká samičí pohlavní buňka oplodněna malou pohyblivou samčí pohlavní buňkou. Někdy splývají rovnou rodičovské buňky (gamonti), příkladem je konjugace nálevníků.[30] U některých mnohobuněčných organizmů se vyvinuly specializované pohlavní orgány.
Pohyb

(transmisní elektronový mikroskop)
Charakter pohybu eukaryotických organismů se do značné míry odvíjí od jejich tělní stavby. Jiný typ pohybu najdeme u jednobuněčných eukaryot, jinak se samozřejmě pohybují mnohobuněční.
U jednobuněčných organismů či jednobuněčných stádií mnohobuněčných organismů (př. spermie) hlavní roli hrají zejména řasinky či bičíky, struktury o průměru asi 0,2 mikrometru. Vlastní úzké tělo (axonema) je celé povlečeno plazmatickou membránou a nachází se tedy uvnitř buňky. Na příčném průřezu je vidět unikátní uspořádání mikrotubulů (9×2+2), při němž je jedna centrální dvojice mikrotubulů obklopena kruhovitě devíti dalšími. Tato struktura je velmi stabilní a liší se jen u několika skupin. Pohyb bičíků a řasinek spotřebovává ATP a je založen na činnosti tzv. ATPáz. Známe však i mnoho jiných struktur umožňujících pohyb samostatných buněk. Améboidní (měňavkovitý) pohyb eukaryot je založen na polymeraci aktinu a vyskytuje se například u zástupců říše Amoebozoa či také mnoha dírkonošců (Foraminifera). Charakteristický druh pohybu u krásnooček (Euglenida) zahrnující vlnivé změny těla se nazývá euglenoidní pohyb. Klouzavým pohybem, který je zřejmě založen na cíleném vylučování slizu, se mohou pohybovat například hromadinky (Gregarinidea) či kokcidie (Coccidea).[30] U mnohobuněčných organismů se rozvinuly zcela jiné druhy lokomoce, povětšinou založené na kontrakci speciálních typů buněk, a zahrnující plazení, chůzi, běh, plavání a podobně.
Rozšíření
Eukaryotické organismy žijí téměř všude, kde je dostatečný konstantní přívod energie, a to buď sluneční záření pro autotrofy, nebo dostatek živin pro výživu heterotrofů. Extrémofilní eukaryota, podobně jako mnohé bakterie a archea, žijí i na mnoha neobvyklých stanovištích s extrémními podmínkami. Omezujícím faktorem je pro eukaryotické organismy zejména teplota prostředí, která na rozdíl od bakterií a archeí (ani u většiny termofilních eukaryot) nesmí přesáhnout 70 °C.[66]
Výčet extrémních prostředí je poměrně dlouhý. Známe anaerobní eukaryota (mnozí protisté s hydrogenozomy, kvasinky), termofily (např. ruducha Cyanidium caldarium, termofilní houby z kompostů, někteří mnohoštětinatci), chladnomilná eukaryota (psychrofilní protisté z arktických a antarktických moří), acidofilové (Cyanidium caldarium a několik hub žijících v pH 0), alkalofilové (někteří protisté žijící v pH 10), halofilové (např. řasa Dunaliella salina), barofilové (hlubokomořští sumýši v 110 MPa) i xerofilové (mnohé pouštní houby a lišejníky).[66]
Odkazy
Poznámky
- vláknité stélky sinic i jiné komplexní bakteriální agregáty se v tomto ohledu považují za kolonie jednobuněčných organismů, i když některé zdroje je mohou chápat jako příklady tzv. agregativní mnohobuněčnosti
- ve smyslu Metazoa
- ve smyslu Archaeplastida, tedy včetně ruduch
- Parakaryota, představovaná jediným popsaným druhem Parakaryon myojinensis s unikátní buněčnou strukturou,[1] nelze považovat za samostatnou doménu, ale vzhledem k nedostatku molekulárních dat za buněčný organismus incertae sedis, u kterého mohou budoucí analýzy prokázat příslušnost k jedné ze známých domén
- nepopsaná šedá větev sousedící se Sar jsou telonemidi
- Nová analýza RNA polymeráz zpochybňuje příbuznost archeí kladu Asgard a eukaryot, Lokiarchaeota a příbuzné archeální linie klade do Euryarchaeot a podporuje přirozenost domény Archaea jako sesterské k eukaryotům,[50] její metodika však byla zásadním způsobem zpochybněna.[51]
Reference
- YAMAGUCHI, Masashi; MORI, Yuko; KOZUKA, Yoshimichi; OKADA, Hitoshi; UEMATSU, Katsuyuki; TAME, Akihiro; FURUKAWA, Hiromitsu. Prokaryote or eukaryote? A unique microorganism from the deep sea. S. 423–431. Journal of Electron Microscopy [online]. The Japanese Society of Microscopy in Oxford University Press, 2012-09-28 [cit. 2021-09-30]. Svazek 61, čís. 6, s. 423–431. Dostupné online. ISSN 2050-5701. DOI 10.1093/jmicro/dfs062. PMID 23024290. (anglicky)
- BURKI, Fabien; ROGER, Andrew J.; BROWN, Matthew W. The New Tree of Eukaryotes. Trends in Ecology & Evolution. 2020-01-01, roč. 35, čís. 1, s. 43–55. Dostupné online [cit. 2020-06-03]. ISSN 0169-5347. DOI 10.1016/j.tree.2019.08.008. (anglicky)
- HASSAN, Rashid M., Robert Scholes, Neville Ash. Ecosystems and human well-being : current state and trends : findings of the Condition and Trends Working Group. Washington, DC: Millennium Ecosystem Assessment (Program). Condition and Trends Working Group, 2005. (The millennium ecosystem assessment series, v. 1). ISBN 1559632275. (anglicky)
- Research Centre for Agricultural and Forest Environment; Polish Academy of Sciences. Globalisation and Biodiversity Protection [online]. Poznan: 2006 [cit. 2008-08-22]. Dostupné v archivu pořízeném dne 2007-07-13. (anglicky)
- KNOLL, Andrew H., Javaux, E. J., Hewitt, D., Cohen, P. Eukaryotic organisms in Proterozoic oceans. Philosophical Transactions of the Royal Society of London, Part B. 2006, roč. 361, čís. 1470, s. 1023–1038. Dostupné online. DOI 10.1098/rstb.2006.1843. PMID 16754612. (anglicky)
- HAN, T. M.; RUNNEGAR, B. Megascopic eukaryotic algae from the 2.1-billion-year-old negaunee iron-formation, Michigan. Science. 1992, roč. 257, čís. 5067, s. 232–5. PMID 1631544. (anglicky)
- VOLF, Petr; HORÁK, Petr. Paraziti a jejich biologie. Praha: Triton, 2007. (Vyd. 1). ISBN 978-80-7387-008-9. S. 318.
- https://phys.org/news/2017-03-complex-life-earth-billion-years.html - Study suggests complex life was present on Earth 2.33 billion years ago
- BROCKS, J. J., Logan, G. A., Buick, R. Summons, R. E. Archean Molecular Fossils and the Early Rise of Eukaryotes. Science. 13. August 1999, roč. 285, čís. 5430, s. 1033–1036. DOI 10.1126/science.285.5430.1033. PMID 10446042. (anglicky)
- University of Berkeley. Life of the Vendian [online]. Dostupné online. (anglicky)
- EMELYANOV, Victor V. Mitochondrial connection to the origin of the eukaryotic cell. European Journal of Biochemistry. Roč. 270, čís. 8, s. 1599–1618. Dostupné online. (anglicky)[nedostupný zdroj]
- PISANI, D., Cotton J. A., McInerney J. O. Supertrees disentangle the chimerical origin of eukaryotic genomes. Mol Biol Evol. Aug 2007, roč. 24, čís. 8, s. 1752–60. Dostupné online. PMID 17504772. (anglicky)
- Martin W, Müller M. The hydrogen hypothesis for the first eukaryote. Nature. Mar 1998, čís. 392, s. 37–41. DOI 10.1038/32033. PMID 9510239. (anglicky)
- BROWN, J. R.; DOOLITTLE, W. F. Archaea and the Prokaryote-to-Eukaryote Transition. Microbiology and Molecular Biology Rewiews. Prosinec 1997, roč. 61, čís. 4, s. 456–502. DOI 1092-2172/97/$04.0010. (anglicky)
- Doolittle W. F. You are what you eat: a gene transfer ratchet could account for bacterial genes in eukaryotic nuclear genomes. Trends in Genetics. 1998, roč. 14, čís. 8, s. 307–311. Dostupné online. DOI 10.1016/S0168-9525(98)01494-2. (anglicky)
- CAVALIER-SMITH, T. The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification.. Int J Syst Evol Microbiol. 52(Pt 1). 2002, s. 7–76. (anglicky)
- BALÁŽ, Vojtech, et al. Smrt jako součást života. Praha: NIDM, 2008. Dostupné v archivu pořízeném dne 2009-03-06. Archivováno 6. 3. 2009 na Wayback Machine
- BROOKE, Nina M., Holland, Peter W. H. The evolution of multicellularity and early animal genomes. Current Opinion in Genetics & Development. 2003-12, roč. 13, čís. 6, s. 599–603. Dostupné online. ISSN 0959-437X. DOI 14638321. (anglicky)
- RUIZ-TRILLO, Ińaki, Gertraud Burger, Peter W. H. Holland, Nicole King, B. Franz Lang, Andrew J. Roger, Michael W. Gray. The origins of multicellularity: a multi-taxon genome initiative. Trends in Genetics: TIG. 2007-03, roč. 23, čís. 3, s. 113–8. Dostupné online. ISSN 0168-9525. DOI S0168-9525(07)00023-6. (anglicky)
- Animals and Fungi: Common Origin, but Independent Approaches to Multicellularity [online]. Dostupné online. (anglicky)
- ZRZAVÝ, Jan. Fylogeneze živočišné říše. Praha: Scientia, 2006. (Vyd. 1). Dostupné v archivu pořízeném dne 2007-11-16. ISBN 80-86960-08-0. S. 255. Archivováno 16. 11. 2007 na Wayback Machine
- University of Berkeley. Cambrian: Life [online]. [cit. 2008-08-15]. Dostupné v archivu pořízeném dne 2008-08-01. (anglicky)
- ČEPIČKA, Ivan; NEUSTUPA, Jiří; HAMPL, Vladimír. Protistologie: záznam přednášek na internetu [online]. Univerzita Karlova, Přírodovědná fakulta UK: 2009. Dostupné online.
- CAMPBELL, N.A; REECE, J.B. Biologie. [s.l.]: Computer press, 2006.
- Journey into the Cell; Eukaryotic and Prokaryotic Cells [online]. Dostupné online. (anglicky)
- PALENIK, Brian, Jane Grimwood, Andrea Aerts, Pierre Rouzé, Asaf Salamov, Nicholas Putnam, Chris Dupont, Richard Jorgensen, Evelyne Derelle, Stephane Rombauts, Kemin Zhou, Robert Otillar, Sabeeha S Merchant, Sheila Podell, Terry Gaasterland, Carolyn Napoli, Karla Gendler, Andrea Manuell, Vera Tai, Olivier Vallon, Gwenael Piganeau, Séverine Jancek, Marc Heijde, Kamel Jabbari, Chris Bowler, Martin Lohr, Steven Robbens, Gregory Werner, Inna Dubchak, Gregory J Pazour, Qinghu Ren, Ian Paulsen, Chuck Delwiche, Jeremy Schmutz, Daniel Rokhsar, Yves Van de Peer, Hervé Moreau, Igor V Grigoriev. The tiny eukaryote Ostreococcus provides genomic insights into the paradox of plankton speciation. Proceedings of the National Academy of Sciences of the United States of America. 2007, roč. 104, čís. 18, s. 7705–10. Dostupné online. ISSN 0027-8424. PMID 17460045. (anglicky)
- KYSILKA, Jiří; KRMENČÍK, Pavel. Toxicon - Escherichia coli [online]. Dostupné online.
- ARMSTRONG, W.P. Physical Properties & Structure of Cells [online]. Dostupné online. (anglicky)
- JENSEN, Mari N. Caulerpa, The World's Largest Single-celled Organism? [online]. Dostupné online. (anglicky)
- HAUSMANN, Klaus; HOLZMANN, Norbert. Protozoologie. Praha: Academia, 2003.
- ČEPIČKA, Ivan; KOLÁŘ, Filip; SYNEK, Petr. Mutualismus, vzájemně prospěšná symbióza; Přípravný text – biologická olympiáda 2007–2008. Praha: NIDM ČR, 2007. S. 87.
- Hedges S. B., Blair J. E., Venturi M. L., Shoe J. L. A molecular timescale of eukaryote evolution and the rise of complex multicellular life.. BMC Evol Biol. Jan 2004, čís. 28;4:2.. Dostupné online. PMID 15005799. (anglicky)
- HAECKEL, Ernst. Generelle Morphologie der Organismen. Berlín: Reimer, 1866. Dostupné online. (německy)
- CHATTON, Edouard. Titres et travaux scientifiques. Sète: Imprimerie Sottano, 1937. (francouzsky)
- COPELAND, Herbert Faulkner. The Classification of Lower Organisms. Palo Alto: Pacific Books, 1956. 302 s. (anglicky)
- WHITTAKER, Robert Harding. New Concepts of Kingdoms of Organisms. Science. 1969, roč. 163, s. 150–160. PMID 5762760. (anglicky)
- WOESE, C. R.; BALCH, W. E.; MAGRUM, L. J.; FOX G. E.; WOLFE, R. S. An ancient divergence among the bacteria. Journal of Molecular Evolution. 1977, roč. 9, čís. 4, s. 305–311. PMID 408502. (anglicky)
- WOESE C.R.; KANDLER O.; WHEELIS M. L. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. U.S.A.. 1990, roč. 87, čís. 12, s. 4576–4579. Dostupné online. PMID 2112744. (anglicky)
- Sapp, Jan. The Prokaryote-Eukaryote Dichotomy: Meanings and Mythology. Microbiol Mol Biol Rev. June 2005, roč. 69, čís. 2, s. 292–305.. Dostupné online. (anglicky)
- Woese C. R., Kandler O., Wheelis M. L. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. U.S.A.. 1990, roč. 87, čís. 12, s. 4576–9. Dostupné online. PMID 2112744. (anglicky)
- R. H. Whittaker. New concepts of kingdoms of organisms. Science. 1969, roč. 163, s. 150–160. (anglicky)
- CAVALIER-SMITH, Thomas. Only six kingdoms of life. Proceedings. Biological Sciences / The Royal Society. 2004, roč. 271, čís. 1545, s. 1251–62. Dostupné online. ISSN 0962-8452. DOI 15306349. (anglicky)
- Simpson A. G., Roger A. J. The real 'kingdoms' of eukaryotes. Curr Biol. 2004, roč. 14, čís. 17, s. R693 - R696. PMID 15341755. (anglicky)
- Taxon: Domain Eukaryota [online]. Dostupné online. (anglicky)
- Biolib - Eukaryota (jaderní) [online]. Dostupné online.
- CAVALIER-SMITH, T. The origin of eukaryotic and archaebacterial cells.. Ann N Y Acad Sci. ;503. 1987, s. 17–54.. (anglicky)
- CAVALIER-SMITH, T. The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification.. Int J Syst Evol Microbiol. 52(Pt 1). 2002, s. 7–76. (anglicky)
- ZAREMBA-NIEDZWIEDZKA, Katarzyna; CACERES, Eva F.; SAW, Jimmy H.; BÄCKSTRÖM, Disa; JUZOKAITE, Lina; VANCAESTER, Emmelien; SEITZ, Kiley W., ANANTHARAMAN, Karthik; STARNAWSKI, Piotr; KJELDSEN, Kasper U.; STOTT, Matthew B.; NUNOURA, Takuro; BANFIELD, Jillian F.; SCHRAMM, Andreas; BAKER, Brett J.; SPANG, Anja; ETTEMA, Thijs J. G. Asgard archaea illuminate the origin of eukaryotic cellular complexity. S. 353–358. Nature [online]. Macmillan Publishers Limited, Springer Nature, 11. leden 2017 [cit. 2017-01-23]. Svazek 541, čís. 7637, s. 353–358. Dostupné online. PDF . ISSN 1476-4687. DOI 10.1038/nature21031. PMID 28077874. (anglicky)
- MIHULKA, Stanislav. Eukaryotní buňky pocházejí z Ásgardu. OSEL.cz [online]. 22. leden 2017 [cit. 2017-01-23]. Dostupné online.
- CUNHA, Violette Da; GAIA, Morgan; GADELLE, Daniele; NASIR, Arshan; FORTERRE, Patrick. Lokiarchaea are close relatives of Euryarchaeota, not bridging the gap between prokaryotes and eukaryotes. S. 1–38. PLoS Genetics [online]. 12. červen 2017. Svazek 13, čís. 6: e1006810, s. 1–38. Dostupné online. ISSN 1553-7404. DOI 10.1371/journal.pgen.1006810. PMID 28604769. (anglicky)
- SPANG, Anja; EME, Laura; SAW, Jimmy H.; CACERES, Eva F.; ZAREMBA-NIEDZWIEDZKA, Katarzyna; LOMBARD, Jonathan; GUY, Lionel, ETTEMA, Thijs J. G. Asgard archaea are the closest prokaryotic relatives of eukaryotes. S. 1–4. PLoS Genetics [online]. 29. březen 2018. Svazek 14, čís. 3: e1007080, s. 1–4. Dostupné online. ISSN 1553-7404. DOI 10.1371/journal.pgen.1007080. PMID 29596421. (anglicky)
- ADL, Sina, et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. S. 4–119. Journal of Eukaryotic Microbiology [online]. John Wiley & Sons, Inc., 26. září 2018. Svazek 66, čís. 1, s. 4–119. Dostupné online. Dostupné také na: . PDF . ISSN 1550-7408. DOI 10.1111/jeu.12691. PMID 30257078. (anglicky)
- PARFREY, L .W., Erika Barbero, Elyse Lasser, Micah Dunthorn, Debashish Bhattacharya, David J Patterson, Laura A Katz. Evaluating support for the current classification of eukaryotic diversity. PLoS Genetics. 2006, roč. 2, čís. 12, s. e220. Dostupné online. ISSN 1553-7404. DOI 06-PLGE-RA-0174R3. (anglicky)
- BAURAIN, Denis; BRINKMANN, Henner; PETERSEN, Jörn, Naiara Rodríguez-Ezpeleta, Alexandra Stechmann, Vincent Demoulin, Andrew J. Roger, Gertraud Burger, B. Franz Lang, Hervé Philippe. Phylogenomic Evidence for Separate Acquisition of Plastids in Cryptophytes, Haptophytes, and Stramenopiles. Molecular Biology and Evolution. 1. březen 2010, svazek 27, čís. 7, s. 1698–1709. Dostupné online [cit. 2010-06-14]. ISSN 0737-4038. DOI 10.1093/molbev/msq059. (anglicky)
- Burki F., Shalchian-Tabrizi K., Minge M., Skjæveland Å., Nikolaev S. I., et al. Phylogenomics Reshuffles the Eukaryotic Supergroups. PLoS ONE. 2007, roč. 2, čís. 8: e790, s. e790. DOI 10.1371/journal.pone.0000790. (anglicky)
- OKAMOTO, Noriko, Chitchai Chantangsi, Aleš Horák, Brian S. Leander, Patrick J. Keeling. Molecular Phylogeny and Description of the Novel Katablepharid Roombia truncata gen. et sp. nov., and Establishment of the Hacrobia Taxon nov. S. 1–11, e7080. PLoS ONE [online]. 17. září 2009 [cit. 2009-10-05]. Svazek 4, čís. 9, s. 1–11. Dostupné online. PDF . DOI 10.1371/journal.pone.0007080. (anglicky)
- ADL, Sina M., et al. The Revised Classification of Eukaryotes. S. 429–514. Journal of Eukaryotic Microbiology [online]. 28. září 2012. Svazek 59, čís. 5, s. 429–514. Dostupné online. PDF . ISSN 1550-7408. DOI 10.1111/j.1550-7408.2012.00644.x. PMID 23020233. (anglicky)
- HAMPL, Vladimír; HUG, Laura; LEIGH, Jessica W., Joel B. Dacks, B. Franz Lang, Alastair G. B. Simpson, Andrew J. Roger. Phylogenomic analyses support the monophyly of Excavata and resolve relationships among eukaryotic “supergroups”. Proceedings of the National Academy of Sciences of the USA. 10. březen 2009, svazek 106, čís. 10, s. 3859–3864. Dostupné online [cit. 2009-12-10]. DOI 10.1073/pnas.0807880106. (anglicky)
- ČEPIČKA, Ivan. Diverzita protist. S. 220–223. Živa [online]. Academia, Středisko společných činností AV ČR, 17. říjen 2019. Roč. 2019, čís. 5, s. 220–223. Dostupné online. ISSN 0044-4812.
- BURKI, Fabien; ROGER, Andrew J.; BROWN, Matthew W.; SIMPSON, Alastair G.B. The New Tree of Eukaryotes. S. 43–55. Trends in Ecology & Evolution [online]. Elsevier Ltd., 9. říjen 2019. Svazek 35, čís. 1, s. 43–55. Dostupné online. Dostupné také na: . ISSN 0169-5347. DOI 10.1016/j.tree.2019.08.008. (anglicky)
- KATINKA, M. D., S. Duprat, E. Cornillot, G. Méténier, F. Thomarat, G. Prensier, V. Barbe, E. Peyretaillade, P. Brottier, P. Wincker, F. Delbac, H. El. Alaoui, P. Peyret, W. Saurin, M. Gouy, J. Weissenbach, C. P. Vivares. Genome sequence and gene compaction of the eukaryote parasite Encephalitozoon cuniculi. Nature. 2001, roč. 414, čís. 6862, s. 450–3. Dostupné online. ISSN 0028-0836. DOI 11719806. (anglicky)
- MIRANDA-SAAVEDRA, Diego, Michael J. R. Stark, Jeremy C. Packer, Christian P. Vivares, Christian Doerig, Geoffrey J. Barton. The complement of protein kinases of the microsporidium Encephalitozoon cuniculi in relation to those of Saccharomyces cerevisiae and Schizosaccharomyces pombe. BMC Genomics. 2007, roč. 8, s. 309. Dostupné online. ISSN 1471-2164. DOI 1471-2164-8-309. (anglicky)
- PARFREY, Laura Wegener, Daniel J. G. Lahr, Laura A. Katz. The dynamic nature of eukaryotic genomes. Molecular Biology and Evolution. 2008, roč. 25, čís. 4, s. 787–94. Dostupné online. ISSN 1537-1719. DOI msn032. (anglicky)
- ROSYPAL, Stanislav. Nový přehled biologie. [s.l.]: Scientia, 2003. S. 797.
- STORCHOVÁ, Zuzana. Mikrosvět, stále se objevují noví zástupci mikrobiální říše. Vesmír. Listopad 1997, roč. 76.
- Roberts, Dave. Eukaryotes in extreme environments [online]. National History Museum. Dostupné online. (anglicky)
Literatura
- ELIÁŠ, Marek. Potíže s kořenem. Vesmír. 2017, roč. 90, čís. 5, s. 270–273. Dostupné online. ISSN 0042-4544.
- MACHÁLEK, Tomáš; MIKEŠOVÁ, Kateřina; TURJANICOVÁ, Libuše; HAMPL, Vladimír. Proměny vyšší systematiky eukaryot a její odraz ve středoškolské biologii. Živa. 2016, čís. 1, s. 27–30. Dostupné online. ISSN 0044-4812.
- ČEPIČKA, Ivan; ELIÁŠ, Marek; HAMPL, Vladimír. Řád z Chaosu. Vesmír. 2010, roč. 89, čís. 7, s. 464–469. Dostupné online. ISSN 0042-4544.
- ČEPIČKA, Ivan. Diverzita protist. Živa. 2019, čís. 5, s. 220–223. Dostupné online. ISSN 0044-4812.
- HAUSMANN, Klaus; HOLZMANN, Norbert. Protozoologie. Praha: Academia, 2003. ISBN 80-200-0978-7.
- VOLF, Petr; HORÁK, Petr. Paraziti a jejich biologie. Praha: Triton, 2007. (Vyd. 1). ISBN 978-80-7387-008-9. S. 318.
- ZRZAVÝ, Jan. Fylogeneze živočišné říše. Praha: Scientia, 2006. (Vyd. 1). ISBN 80-86960-08-0. S. 255.
- KALINA, Tomáš; VÁŇA, Jiří. Sinice, řasy, houby, mechorosty a podobné organismy v současné biologii. Praha: Karolinum, 2005. 606 s. ISBN 80-246-1036-1.
- PETR, Jaroslav. Zrození jádra. VTM Science. 2005, roč. 59, čís. 3.
- STORCHOVÁ, Zuzana. Mikrosvět, stále se objevují noví zástupci mikrobiální říše. Vesmír. Listopad 1997, roč. 76.
- EMELYANOV, Victor V. Mitochondrial connection to the origin of the eukaryotic cell. European Journal of Biochemistry. Roč. 270, čís. 8, s. 1599–1618. Dostupné online. (anglicky)[nedostupný zdroj]
Externí odkazy
 Obrázky, zvuky či videa k tématu Eukaryota na Wikimedia Commons
Obrázky, zvuky či videa k tématu Eukaryota na Wikimedia Commons  Taxon Eukaryota ve Wikidruzích
Taxon Eukaryota ve Wikidruzích- (česky) Přednášky z protistologie na Přírodovědné fakultě UK ke stažení a k poslechu
- (anglicky) Obecné informace o eukaryotických organismech a strom života
- (anglicky) Přehledně uspořádaná klasifikace eukaryot dle Adla et al. na Taxonomiconu
- (česky) Eukaryotická buňka – organely
- (česky) Starší článek v časopise Vesmír zachycující tehdejší pohled na evoluci eukaryotické buňky (dnes překonaný)