Mitochondrie
Mitochondrie je membránově obalená organela, kterou lze nalézt ve většině[pozn. 1] eukaryotických (např. lidských) buněk. Dosahuje obvykle rozměrů v řádu i několika mikrometrů a v buňce se jich může vyskytovat několik stovek, ale i sto tisíc. Obvykle se považují za struktury klobásovitého tvaru, nicméně v řadě případů vytváří spíše bohatě se větvící síť vláken po celé buňce. Funkce mitochondrií se do jisté míry dá přirovnat k buněčné elektrárně, jelikož v nich díky procesům buněčného dýchání vzniká energeticky bohatý adenosintrifosfát (ATP) používaný jako „palivo“ pro průběh jiných reakcí v celé buňce. Mitochondrie je uzavřena dvěma membránami. Vnější membrána je značně pórovitá a skutečnou bariéru pro malé molekuly představuje spíše vnitřní membrána. Na vnitřní membráně a uvnitř ní se také odehrávají ty nejdůležitější metabolické pochody. Konkrétně v mitochondriích probíhá Krebsův cyklus, dýchací řetězec, ale také beta-oxidace mastných kyselin. Navíc se mitochondrie podílejí na dalších procesech, jako je buněčná diferenciace, buněčná smrt i kontrola buněčného cyklu a růstu.[2] Na druhou stranu mohou mít poruchy jejich funkce za následek různá mitochondriální onemocnění, která obvykle souvisí s neschopností správně provádět metabolické mitochondriální pochody.
.jpg.webp)
(transmisní elektronový mikroskop)
Některé vlastnosti mitochondrií je činí unikátními z evolučního hlediska. V mnohých ohledech totiž připomínají bakterie; dnes je známo, že se ve skutečnosti opravdu před asi dvěma miliardami let vyvinuly z bakteriálních předků. Přesný scénář však není s určitostí znám. Tehdy mimo jiné byla teplota moří přibližně 50 °C.[3] To také odpovídá zjištění, že mitochondrie i nyní uvnitř buněk fungují při 50 °C, na což jsou evolučně optimalizovány i dané enzymy.[4] Uvnitř mitochondrie se nachází stále zachovalá mitochondriální DNA, která se v mnohém podobá té bakteriální. Spolu s plastidy, jež mají podobný původ, se řadí mezi semiautonomní buněčné organely. V některých případech dochází ke značné redukci mitochondrií, tak u některých jednobuněčných organismů vznikly hydrogenozomy a mitozomy.
Historie výzkumu
S rozvojem mikroskopie byly v buňkách identifikovány různé struktury, o nichž se až později zjistilo, že představují jediný typ organely, tedy mitochondrii. Pro tyto různě vypadající struktury bylo používáno nespočetné množství různých termínů, jako například chondriozom, chondrioplast, fuchsinofilní či parabazální tělísko a podobně. Obvykle byla tato slova odvozena z řeckého výrazu pro zrno (χόνδρος, „chondros“), případně z jeho anglického („grain“) a německého („Korn“) ekvivalentu. Se zlepšením pozorovacích metod se na konci 19. století ukázalo, že tyto struktury mohou nabývat i vláknitého tvaru (vlákno je řecky μίτος, tedy „mitos“), a výrazy chondros a mitos daly společně vzniknout složenině, slovu mitochondrie.[5]
V letech 1912–1913 B. F. Kingsbury a O. Warburg prokázali, že mitochondrie jsou sídlem energetického metabolismu eukaryotické buňky. Detaily složitých procesů odehrávajících se v nitru mitochondrie však byly odhalovány postupně (Hans Adolf Krebs, 1937 – Krebsův cyklus) a k poznání přispěla zejména postupná izolace jednotlivých enzymů účastnících se těchto cyklů. Postupně byly odhalovány podrobnější informace o stavbě mitochondrií, například skutečnost, že jsou obaleny dvěma membránami, nebo v roce 1963 i existence mitochondriální DNA.[5]
Evoluční původ
V dnešní době je obecně akceptováno, že mitochondrie představují potomky endosymbiotické bakterie, které se v procesu vzniku eukaryotické buňky určitým způsobem transformovala v semiautonomní organelu.[5][6] Předpokládá se, že touto bakterií, která vstoupila do primitivní eukaryotické buňky, byl zástupce alfaproteobakterií z příbuzenského okruhu rodu Rickettsia.[7] Tyto názory však byly odbornou veřejností poměrně dlouho odmítány jako „nehodící se do slušné přírodovědné společnosti“.[5] První představy o takovém původu se však paradoxně začaly objevovat již na konci 19. století, kdy ještě ani nebyl ustanoven termín mitochondrie (např. Richard Altmann, 1890). Známými zastánci této tzv. endosymbiotické teorie byl na začátku 20. století Konstantin Merežkovskij, případně v druhé polovině 20. století Lynn Margulisová.[5] Silnou podporu pak teorie nalezla v molekulárních analýzách genomů, prokazujících evoluční příbuznost alfaproteobakterií a mitochondrií, nebo v objevu analogie mitochondriálních krist s intracytoplazmatickými membránámi alfaproteobakterií.[8]
Přesný scénář „endosymbiotické události“, tedy procesu, při němž se bakterie fakticky změnila na mitochondrii, je stále poměrně zahalen tajemstvím. Podle tzv. molekulárních hodin (metoda založená na pozorování mutací v DNA) se zdá, že k tomu došlo přibližně před 2 miliardami let, tedy zřejmě nedlouho po vzniku samotné eukaryotické buňky.[9] Dnes se také považuje za prokázané, že všechny známé eukaryotické organizmy buď mitochondrii mají, nebo alespoň měly za předka eukaryota, který mitochondrii měl. Proto se dnes ustupuje od termínu Archezoa, který označoval skupiny eukaryot, jež vznikly ještě před „endosymbiotickou událostí“: žádné takové nebyly objeveny.[7] Zřejmé je také to, že mitochondrie vznikla v historii jednou jedinkrát, a všechny současné mitochondrie vznikly na základě této unikátní události (mluví se o tzv. monofyletickém původu).[5]
Konkrétní scénář poskytují například známé vodíkové hypotézy, které jsou v podstatě všechny založené na předpokladu, že hostitelskou buňkou byla anaerobní, vodík metabolizující, autotrofní archebakterie, která jistým způsobem pohltila symbiotickou bakterii schopnou v rámci své respirace produkovat vodík. Potomci této bakterie následně prošli evolucí a změnili se na mitochondrie.[10][11] Ať už to bylo tak, či onak, po endosymbiotické události muselo zcela jistě dojít k částečné redukci genomu symbiotické bakterie a pravděpodobně také k tzv. horizontálnímu transferu, tedy přechodu části genů z mitochondrie do jádra. V současnosti je 600–1000 mitochondriálních proteinů kódováno jadernou DNA a v mitochondriích je uloženo nanejvýš několik desítek genů[12] (viz kap. genetická informace).
Redukce mitochondrií
Ačkoliv všechny současné známé eukaryotické organizmy pochází ze společného předka, který mitochondrii měl, u mnohých eukaryot byly místo mitochondrií popsány jen jakési redukované organely neschopné oxidativní fosforylace, jako jsou hydrogenozomy a mitozomy. Tyto organely jsou často schopné syntézy ATP bez přítomnosti kyslíku, a hodí se tedy do anaerobního prostředí. Hydrogenozomy a mitozomy byly popsány zejména (ne však výhradně) v říších Excavata a Amoebozoa, známé jsou například Entamoeba histolytica, Giardia intestinalis, různé mikrosporidie (Microsporidia) a chytridiomycety (Chytridiomycota), případně různí anaerobní nálevníci (Ciliophora).
Redukce mitochondrie spojená s úplnou ztrátou schopnosti buněčného dýchání byla v r. 2020 nalezena i u mnohobuněčného organismu. Henneguya salminicola, výtrusenka parazitijící na lososech, vývojem přišla nejen o mitochondriální DNA nutnou pro dýchání, ale i části jaderné DNA nezbytné pro reprodukci mitochondriální DNA. Energii získává od svého hostitele pomocí specializovaných bílkovin.[13][14]
V roce 2016 byla dokonce poprvé popsána prokazatelná absence mitochondrie i jakékoli jiné organely tohoto typu, ba i jakéhokoli jaderného genu či proteinu mitochondriálního původu u eukaryotického organismu, a to u oxymonád rodu Monocercomonoides, střevního parazita činčil. Vzhledem k tomu, že se jedná o značně odvozeného (nemá např. ani peroxizomy, ani Golgiho aparát) a svou stavbou přitom velmi komplexního prvoka, lze předpokládat, že o pozůstatky mitochondrií přišel druhotně a své buněčné procesy modifikované parazitickým životem má nastaveny tak, že mitochondriální geny/proteiny pravděpodobně ani nepotřebuje. Autoři studie, mj. i biologové Univerzity Karlovy a Ostravské univerzity, předpokládají, že podobnou absenci budou vykazovat i další oxymonády. Byla tak vyvrácena představa, že eukaryotní buňka bez mitochondrií nebo jejich pozůstatků ani nemůže fungovat.[15][16][17]
Stavba
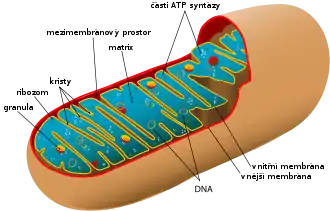
Mitochondrie mohou mít velice rozmanité prostorové uspořádání a velikost, sahající od vláknité struktury až po kompaktní zrna. Například v hepatocytech a fibroblastech mají klobásovitý tvar a rozměry 3–4 μm na délku a 1 μm na šířku.[5] Tím také připomínají bakterie (např. E. coli dosahuje délky 2–3 μm a šířky 0,6 μm.[18]). Jindy vzniká v buňkách spíše síťovité retikulum mitochondriálních membrán tvořené tenkými větvícími se trubičkami.[19] Co se týče počtu, v hepatocytech se obvykle nachází asi 800 mitochondrií, lidský oocyt jich však obsahuje až 100 000, měňavka Chaos chaos dokonce 500 000, ale např. lidská spermie jen několik[5] a Trypanosoma[20] nebo buňka lidské sítnice[19] jen jednu. Mitochondrie se množí procesem podobným binárnímu dělení bakterií, může však docházet i opačnému procesu, tedy fúzi dvou mitochondrií dohromady.[21]
Jednotlivé mitochondrie mohou být vzájemně propojené nanotunely specifické stavby, odlišné od stavby nanotrubiček cytoskeletu. Mají dvoumembránovou stěnu a pravděpodobně slouží k transportu proteinů.[22][23]
Co se týče biochemické stavby, v mitochondriích bylo identifikováno několik stovek proteinů (např. v mitochondriích v srdci se toto číslo odhaduje na 615[24]). Až polovina mitochondriálních proteinů je zřejmě specifická pro danou tkáň.[19]
Membrány
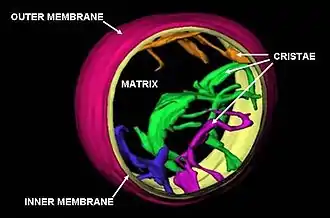
Mitochondrie je, podobně jako například primární plastidy, oddělena od okolí dvěma fosfolipidovými membránami, mezi nimiž je tzv. mezimembránový prostor. Takový systém je srovnatelný se stavbou membrány gramnegativních bakterií, z nichž se mitochondrie vyvinuly.[25] Mezimembránový prostor má složení velice podobné složení okolního cytosolu, tzn. obsahuje mnohem méně proteinů, než vnitřní mitochondriální prostor (tzv. matrix). Z bílkovin je v mezimembránovém prostoru zastoupen hlavně cytochrom c a různé kinázy.
Vnější membrána mitochondrií představuje první bariéru mezi vnitřním prostředím organely a okolním cytosolem. Je v úzkém kontaktu s endoplazmatickým retikulem, a do jisté míry se mu podobá i svou stavbou. Obsahuje však ve své ploše mnoho pórů (mitochondriální poriny VDAC), a tak je tato membrána propustná pro většinu látek s molekulovou hmotností nepřesahující přibližně 5000 Da[26] (někdy se uvádí i 10 000 Da[27]). Proto zamezuje spíše jen vstupu bílkovin a jiných makromolekul. To je významné především ze dvou hledisek: kdyby byly póry větší, nejen, že by unikly z mezimembránového prostoru důležité proteiny dýchacího řetězce, ale uvolněním cytochromu c by se navíc nastartovala v buňce programovaná buněčná smrt. Přesto se proteiny transportovat musí, a tak vnější membrána obsahuje tzv. Tom komplex, který je přenáší z cytoplazmy do intermembránového prostoru. Dále se na vnější membráně nacházejí enzymy, které jsou součástí metabolismu mastných kyselin a fosfolipidů.[26]
Vnitřní membrána má velkou plochu, protože je zvlněna v tzv. kristy. U jaterních buněk, které mají kristy ještě relativně malé, je plocha vnitřní membrány mitochondrie 15× větší než plocha plazmatické membrány buňky.[27] Obecně závisí velikost krist na metabolické aktivitě konkrétní buňky.[27] Vnitřní membrána je zásadní z hlediska funkčnosti mitochondrie. Je schopná propouštět molekuly jen velice selektivně, ionty přes ni téměř nedifundují, a to možná díky speciální stavbě fosfolipidové dvouvrstvy. Obsahuje totiž neobvyklý fosfolipid kardiolipin, z jehož fosfátové hlavy vychází ne dvě, ale hned čtyři mastné kyseliny.[28] Na této membráně je totiž umístěno množství enzymů tzv. dýchacího řetězce, včetně ATP syntázy a enzymu ANT, který vynáší konečný produkt dýchání – adenosintrifosfát – ven do buňky. Vnitřní membrána (podobně jako ta vnější) dále obsahuje komplex, který umožňuje přenos bílkovin přes ni – ten se označuje jako Tim komplex.[26]
Mitochondriální kristy mohou mít i předendosymbiotický původ, protože jsou analogické intracytoplazmatickým membránám alfaproteobakterií. Prvotní mitochondrie tak možná zdědily od svých bakteriálních předků existující ultrastruktury adaptované na účinný přenos energie.[8]
Matrix
Uvnitř mitochondrie se nachází matrix, hustá hmota s obsahem vody méně než 50%,[27] velice bohatá na bílkoviny. Patří k nim zejména enzymy Krebsova cyklu. Dále matrix obsahuje různé nukleotidové koenzymy, anorganické ionty (vápník), z větších struktur mitochondriální DNA, příslušnou RNA a dokonce mitochondriální ribozomy (ve shodě s endosymbiotickou teorií jsou velice podobné ribozomům bakteriálním[25]).
Transport bílkovin do mitochondrie
Mitochondrie jsou schopné vyrábět si jen velmi omezené množství bílkovin, a většina mitochondriálních bílkovin je proto vytvářena v cytosolu (z jaderných genů). Tyto proteiny jsou vzápětí přenášeny skrz jednu či obě mitochondriální membrány, aby skončily na místě svého finálního určení. Transport do mitochondrie je složitý molekulární proces závislý na celé řadě transportních proteinů. Mitochondriální protein je rozeznán podle speciální aminokyselinové signální sekvence bohaté na bazické, hydroxylované a hydrofobní aminokyseliny. Tato sekvence (obvykle na N-konci) je ve většině případů po dokončení transportu z bílkoviny odstraněna.[12]
V cytosolu se mitochondriální bílkoviny obvykle navážou na chaperony typu Hsp70. Díky nim zůstávají v nesbaleném stavu – v tomto stavu je transport bílkovin skrz kanály jednodušší. První překážkou je vnější membrána, v níž jsou umístěny Tom kanály Tom40. Některé mitochondriální bílkoviny se začlení do vnější membrány (jako namátkou právě Tom40), jiné zůstávají v mezimembránovém prostoru a další pokračují svou pouť skrz vnitřní membránu. Proteiny vstupující do matrix využívají komplex Tim23 (složený z Tim23 a Tim17), zatímco proteiny, jež se mají začlenit do vnitřní membrány a zůstat tam jako transmembránové, využívají komplex Tim22.[12]
Funkce
Buněčné dýchání a syntéza ATP
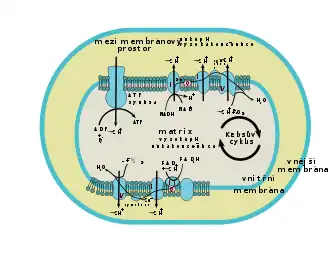
V mitochondriích probíhá převážná část buněčného dýchání (vyjma zejména glykolýzy, jež probíhá v cytosolu). Tento sled reakcí představuje rozklad různých organických látek, čímž se získává energie potřebná pro syntézu adenosintrifosfátu (ATP) z ADP. Výchozí látkou jsou zejména pyruvát a mastné kyseliny, obě látky v mitochondriální matrix prochází reakcemi, při nichž vzniká acetylkoenzym A (pyruvát prochází dekarboxylací, mastné kyseliny beta-oxidací). Acetylkoenzym A následně vstupuje do Krebsova cyklu, což je série reakcí, které umožňují redukci koenzymů NAD+ na NADH a FAD na FADH2. Elektrony z těchto koenzymů jsou předávány do dýchacího řetězce, jenž je umístěn na vnitřní membráně mitochondrie. Dýchací řetězec přenáší do mezimembránového prostoru vodíkové kationty (H+, čímž vzniká v prostoru mezi membránami kyselé pH. Toto pH má tendenci se vyrovnávat, a tak prochází otvorem v enzymu ATP syntáze zpět dovnitř buňky. Průchodem H+ však tento enzym vytváří ATP, jež je kýženým produktem celého sledu reakcí.[28]
Další funkce
Přestože je mitochondrie známá převážně jako energetická jednotka buňky, byly k ní přiřazeny i další důležité funkce. S buněčným dýcháním souvisí skutečnost, že jsou mitochondrie v hnědé tukové tkáni schopné produkovat teplo. Mechanismus je elegantní: místo toho, aby vodíkové kationty procházely ATP syntázou, prochází protonovým kanálem termogeninem (UCP1), jenž je právě zodpovědný za vznik tepelné energie.[29][30]
Navíc může matrix mitochondrií fungovat jako zásobárna vápníku (podobně jako endoplazmatické retikulum), čímž udržují vnitřní homeostázu buňky. Dovnitř se dostávají ionty vápníku speciálním přenašečem, který je poháněn membránovým potenciálem,[31][32] při masivním uvolňování vápníku ven se může tímto způsobem navodit v buňce určitá odpověď (produkce hormonů, neurotransmiterů, atp.). Navíc je v mitochondrii přítomna řada proteinů spouštějících apoptózu, programovanou buněčnou smrt buňky,[33] regulace buněčné proliferace a metabolismu,[2] probíhají však v nich i některé části syntézy hemu[34] a steroidů.[35]
Je známo, že v mitochondriích jsou (jako vedlejší produkt buněčného dýchání) produkovány kyslíkové radikály, jež mohou způsobit četné mutace v mitochondriální DNA.[36] Čím více je tato DNA poškozována, tím defektnější molekuly vznikají, a tím spíš se pak mohou degenerativní změny prohlubovat. Množství změn je v mitochondriích pozorováno během lidského stárnutí.[37] Některé změny mohou být dávány do souvislosti s Parkinsonovou chorobou.[38]
Genetická informace
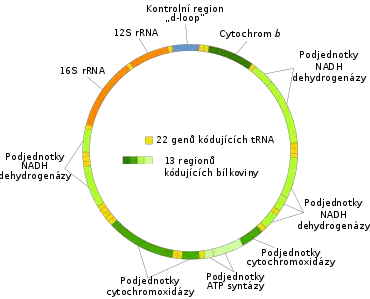
Mitochondriální DNA (též mtDNA) je označení pro DNA, která se nachází v mitochondriích a tvoří tak součást mimojaderné genetické informace. Jedná se o pozůstatek genomu bakterií, z nichž mitochondrie vznikla, a dodnes s nimi má mnoho společných rysů.[7] Mitochondriální DNA je totiž zpravidla kruhová (cirkulární) a svým charakterem se podobá prokaryontnímu nukleoidu a nikoliv eukaryotickým chromozomům. Je v porovnání s jaderným genomem velice malá. Například lidská mitochondriální DNA má velikost 16 569 párů bází,[39] celkem obsahuje 37 genů, z toho 24 představují geny pro různou nekódující RNA (2 geny pro 16S a 23S rRNA a 22 genů pro tRNA). Zbývajících 13 genů kóduje vlastní mitochondriální polypeptidy podílející se na enzymatické výbavě mitochondrií.[40] Na druhou stranu může být mtDNA v mitochondrii přítomná v několika kopiích a s přihlédnutím ke skutečnosti, že mitochondrií může být v buňce i několik set tisíc, může paradoxně představovat genom v mitochondrii většinovou složku celkového buněčného genomu.[28]
Mitochondriální DNA se dědí obvykle po matce, tedy maternálně, jelikož z vajíčka (a nikoliv ze spermie) pochází obvykle veškerá mitochondriální genetická informace zárodku.[28] To je také jeden z důvodů, proč je mtDNA velice cenným nástrojem genetiků a molekulárních biologů – geny matky a otce se nerekombinují. Navíc mtDNA mutuje rychleji než jaderný genom, čímž umožňuje zkoumat změny v mnohem kratším časovém měřítku. Tyto a další vlastnosti mtDNA umožňují zkoumat například migrace lidských populací (viz termín mitochondriální Eva).[5]
Kontakty mitochondrie-ER
Mitochondrie vytváří časté kontakty s endoplasmatickým retikulem, tzv. kontaktní místa mitochondrie – endoplazmatické retikulum (MERCS; z angl. Mitochondria and Endoplasmic Reticulum Contact Sites). Alternativně jsou tyto kontakty nazývány jako membrány asociované s mitochondriemi (MAM; angl. Mitochondria-Associated Membranes). Tato místa jsou oblasti mezi mitochondriemi a endoplazmatickým retikulem, kde membrány obou organel leží paralelně blízko sebe (10–90 nm).[41] Vzájemná blízkost tak umožňuje komunikaci a výměnu látek mezi oběma organelami. Kontaktní místa ovlivňují některé buněčné události a procesy, jako např. homeostázu vápníku[42], metabolismus a import lipidů z ER do mitochondrií[43], mitochondriální dělení[44], autofagii[45] či apoptózu.[46] Uvažuje se také o jejich roli při rozvoji neurodegenerativních onemocnění, např. při rozvoji Alzheimerovy choroby či Parkinsonovy nemoci.
Poruchy a onemocnění
Mitochondriální onemocnění je označení pro takové onemocnění, které je způsobené poruchou funkce mitochondrií, a to v širším významu jak vrozené, tak i získané. Za vrozené se považují onemocnění způsobené mutacemi v genech v jaderné DNA či v mitochondriální DNA). Naopak získané poruchy mitochondrií mohou být výsledkem infekce, užívání léků či vlivem negativních podmínek prostředí.[47] Obvykle se jedná o onemocnění způsobená poruchami v mitochondriálním metabolismu, zejména poruchy oxidativní fosforylace.[48]
Odkazy
Poznámky
- Mitochondrie nebyly nalezeny u některých prvoků (viz kapitola „redukce mitochondrií“), ani v lidských červených krvinkách[1]
Reference
V tomto článku byl použit překlad textu z článku mitochondrion na anglické Wikipedii.
- PALEČEK, Jiří. Biologie buňky. II, Základy mikroskopické histologie. Praha: Karolinum, 2000 (dotisk). ISBN 80-7184-279-6.
- McBride HM, Neuspiel M, Wasiak S. Mitochondria: more than just a powerhouse. Curr Biol.. Červenec 2006, roč. 16, čís. 14, s. R551–60. Dostupné online. DOI 10.1016/j.cub.2006.06.054. PMID 16860735.[nedostupný zdroj]
- https://www.researchgate.net/publication/6730234_A_palaeotemperature_curve_for_the_Precambrian_oceans_based_on_silicon_isotopes_in_cherts - A palaeotemperature curve for the Precambrian oceans based on silicon isotopes in cherts
- https://phys.org/news/2018-01-mitochondria-degrees.html - Do our mitochondria run at 50 degrees C?
- SCHEFFLER, Immo E. Mitochondria. [s.l.]: John Wiley & Sons, Inc., 1999. ISBN 0-471-19422-0.
- GRAY, M. W. The endosymbiont hypothesis revisited. International Review of Cytology. 1992, roč. 141, s. 233–357. Dostupné online. ISSN 0074-7696.
- EMELYANOV, Victor V. Mitochondrial connection to the origin of the eukaryotic cell. European Journal of Biochemistry. Roč. 270, čís. 8, s. 1599–1618. Dostupné online. (anglicky)[nedostupný zdroj]
- MUÑOZ-GÓMEZ, Sergio A.; WIDEMAN, Jeremy G.; ROGER, Andrew J.; SLAMOVITS, Claudio H. The Origin of Mitochondrial Cristae from Alphaproteobacteria. S. 943–956. Molecular Biology and Evolution [online]. 12. leden 2017. Svazek 34, čís. 4, s. 943–956. Dostupné online. ISSN 1537-1719. DOI 10.1093/molbev/msw298. PMID 28087774. (anglicky)
- HAMPL, Vladimír. Eukaryogeneze (PROTISTOLOGIE 2010) [online]. 2010. Dostupné online.[nedostupný zdroj]
- Martin W, Müller M. The hydrogen hypothesis for the first eukaryote. Nature. Mar 1998, čís. 392, s. 37–41. DOI 10.1038/32033. PMID 9510239. (anglicky)
- MOREIRA, Lopez-Garcia. Symbiosis between methanogenic archaea and delta-proteobacteria as the origin of eukaryotes: the syntrophic hypothesis. Journal of Molecular Evolution. 1998-11, roč. 47, čís. 5, s. 517–30. Dostupné online. ISSN 0022-2844. DOI 9797402.
- POLLARD, Thomas D; EARNSHAW, William C. Cell Biology. 2. vyd. [s.l.]: Saunders, 2007. ISBN 1416022554. S. 928.
- DVOŘÁK, Ondřej. Parazit bez dechu. Akademon [online]. 2020-04-17. Dostupné online.
- YAHALOMI, Dayana; ATKINSON, Stephen D.; NEUHOF, Moran; CHANG, E. Sally; PHILIPPE, Hervé; CARTWRIGHT, Paulyn; BARTHOLOMEW, Jerri L. A cnidarian parasite of salmon (Myxozoa: Henneguya) lacks a mitochondrial genome. S. 5358–5363. Proceedings of the National Academy of Sciences USA (PNAS) [online]. National Academy of Sciences, 24. únor 2020 [cit. 2020-04-21]. Svazek 117, čís. 10, s. 5358–5363. Dostupné v archivu pořízeném z originálu dne 2020-04-22. Dostupné také na: . Dále dostupné na: . ISSN 1091-6490. DOI 10.1073/pnas.1909907117. PMID 32094163. (anglicky)
- MIHULKA, Stanislav. Prvoci z činčil jsou kompletně bez mitochondrií. OSEL.cz [online]. 24. květen 2016. Dostupné online.
- KARNKOWSKA, Anna; VACEK, Vojtěch; ZUBÁČOVÁ, Zuzana; TREITLI, Sebastian C.; PETRŽELKOVÁ, Romana; EME, Laura; NOVÁK, Lukáš, ŽÁRSKÝ, Vojtěch; BARLOW, Lael D.; HERMAN, Emily K.; SOUKAL, Petr; HROUDOVÁ, Miluše; DOLEŽAL, Pavel; STAIRS, Courtney W.; ROGER, Andrew J.; ELIÁŠ, Marek; DACKS, Joel B.; VLČEK, Čestmír; HAMPL, Vladimír. A Eukaryote without a Mitochondrial Organelle. S. 1274–1284. Current Biology [online]. Elsevier B.V., 12. květen 2016. Svazek 26, čís. 10, s. 1274–1284. Dostupné online. Dostupné také na: . ISSN 0960-9822. DOI 10.1016/j.cub.2016.03.053. PMID 27185558. (anglicky)
- Cell Press. Surprise! This eukaryote completely lacks mitochondria. Phys.org [online]. 12. květen 2016. Popularizační článek k předchozí referenci. Dostupné online. (anglicky)
- KYSILKA, Jiří; KRMENČÍK, Pavel. Toxicon - Escherichia coli [online]. Dostupné online.
- JENSEN, R. E. Control of mitochondrial shape. Curr Opin Cell Biol.. 2005, roč. 17, čís. 4, s. 384–8. Dostupné online. ISSN 0955-0674.
- KUBIŠTA, Václav. Buněčné základy životních dějů. Praha: Scientia, 1998. ISBN 80-7183-109-3. S. 210.
- ALBERTS, Bruce, et al. Essential Cell Biology. 2. vyd. New York: Garland Science, 2004. Dostupné online.
- VINCENT, Amy E.; TURNBULL, Doug M.; EISNER, Veronica; HAJNÓCZKY, György; PICARD, Martin. Mitochondrial Nanotunnels. Trends in Cell Biology [online]. Elsevier Inc., 18. září 2017. Online před tiskem. Dostupné online. ISSN 0962-8924. DOI 10.1016/j.tcb.2017.08.009. (anglicky)
- HEWITT, John. Complete structure of mitochondrial respiratory supercomplex decoded. Phys.Org [online]. 22. září 2017. Dostupné online. (anglicky)
- Taylor SW, Fahy E, Zhang B, Glenn GM, Warnock DE, Wiley S, Murphy AN, Gaucher SP, Capaldi RA, Gibson BW, Ghosh SS. Characterization of the human heart mitochondrial proteome. Nat Biotechnol.. Březen 2003, roč. 21, čís. 3, s. 281–6. Dostupné online. DOI 10.1038/nbt793. PMID 12592411.
- ROBINSON, Richard. Reference Biology. New York: Thomson Gale, 2002. ISBN 0-02-865551-6.
- Mitochondrial Membranes, Structural Organization;. In: Lennarz,W.J., Lane, M.D. Encyclopedia of Biological Chemistry , Four-Volume Set, 1-4. [s.l.]: [s.n.]
- VOET, Donald; VOET, Judith. Biochemie. 1.. vyd. Praha: Victoria Publishing, 1995. ISBN 80-85605-44-9.
- Alberts, Bruce , et al. The Molecular Biology of the Cell. [s.l.]: Garland Science, 2002. (4th. ed). Dostupné online. ISBN 0-8153-3218-1. Kapitola Energy Conversion: Mitochondria and Chloroplasts.
- Mozo J, Emre Y, Bouillaud F, Ricquier D, Criscuolo F. Thermoregulation: What Role for UCPs in Mammals and Birds?. Bioscience Reports.. Listopad 2005, roč. 25, s. 227–249. DOI 10.1007/s10540-005-2887-4.
- Nicholls DG, Lindberg O. Brown-adipose-tissue mitochondria. The influence of albumin and nucleotides on passive ion permeabilities. Eur. J. Biochem.. 1973, roč. 37, s. R551. DOI 10.1111/j.1432-1033.1973.tb03014.x. PMID 4777251.
- Miller RJ. Mitochondria – the kraken wakes!. Trends in Neurosci.. 1998, roč. 21, čís. 3, s. 95–97 doi=10.1016/S0166–2236(97)01206–X.
- Siegel GJ, Agranoff BW, Fisher SK, Albers RW, Uhler MD. Basic Neurochemistry. 6. vyd. [s.l.]: Lippincott Williams & Wilkins, 1999. ISBN 0-397-51820-X.
- Green DR. Apoptotic pathways: the roads to ruin. Cell.. Září 1998, roč. 94, čís. 6, s. 695–8. DOI 10.1016/S0092-8674(00)81728-6. PMID 9753316.
- OH-HAMA, T. Evolutionary consideration on 5-aminolevulinate synthase in nature. Orig Life Evol Biosph.. Srpen 1997, roč. 27, čís. 4, s. 405–12. DOI 10.1023/A:1006583601341. PMID 9249985.
- ROSSIER, M. F. T channels and steroid biosynthesis: in search of a link with mitochondria. Cell Calcium.. 2006, roč. 40, čís. 2, s. 155–64. DOI 10.1016/j.ceca.2006.04.020. PMID 16759697.
- Richter C, Park J, Ames BN. Normal Oxidative Damage to Mitochondrial and Nuclear DNA is Extensive. PNAS. 1988 September, roč. 85, čís. 17, s. 6465–6467. DOI 10.1073/pnas.85.17.6465. PMID 3413108.
- Mitochondria and Aging. [online]. Dostupné online.
- Bender A, Krishnan KJ, Morris CM, Taylor GA, Reeve AK, Perry RH, Jaros E, Hersheson JS, Betts J, Klopstock T, Taylor RW, Turnbull DM. High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat Gen.. 2006, roč. 38, s. 515–517. DOI 10.1038/ng1769. PMID 16604074.
- ALBERTS, Bruce , et al. The Molecular Biology of the Cell. [s.l.]: Garland Science, 2002. (4th. ed). Dostupné online. ISBN 0-8153-3218-1.
- The Report of the Committee on the Human Mitochondrial Genome, http://www.mitomap.org/report.html Archivováno 22. 12. 2008 na Wayback Machine
- GIACOMELLO, M; PELLEGRINI, L. The coming of age of the mitochondria–ER contact: a matter of thickness. Cell Death & Differentiation. 2016-06-24, roč. 23, čís. 9, s. 1417–1427. Dostupné online [cit. 2022-01-21]. ISSN 1350-9047. DOI 10.1038/cdd.2016.52.
- CSORDÁS, G.; HAJNÓCZKY, G. Sorting of calcium signals at the junctions of endoplasmic reticulum and mitochondria. Cell Calcium. 2001-04, roč. 29, čís. 4, s. 249–262. Dostupné online [cit. 2022-01-21]. ISSN 0143-4160. DOI 10.1054/ceca.2000.0191.
- RUSIÑOL, A E; CUI, Z; CHEN, M H. A unique mitochondria-associated membrane fraction from rat liver has a high capacity for lipid synthesis and contains pre-Golgi secretory proteins including nascent lipoproteins.. Journal of Biological Chemistry. 1994-11, roč. 269, čís. 44, s. 27494–27502. Dostupné online [cit. 2022-01-21]. ISSN 0021-9258. DOI 10.1016/s0021-9258(18)47012-3.
- ROBIN., Friedman, Jonathan. The mechanism and function of ER dynamics.. [s.l.]: [s.n.] Dostupné online. ISBN 978-1-267-33532-6, ISBN 1-267-33532-7. OCLC 874093806
- ZHONG, Qing. Faculty Opinions recommendation of Autophagosomes form at ER-mitochondria contact sites.. Faculty Opinions – Post-Publication Peer Review of the Biomedical Literature [online]. 2013-08-19 [cit. 2022-01-21]. Dostupné online.
- IWASAWA, Ryota; MAHUL-MELLIER, Anne-Laure; DATLER, Christoph. Fis1 and Bap31 bridge the mitochondria-ER interface to establish a platform for apoptosis induction. The EMBO Journal. 2010-12-24, roč. 30, čís. 3, s. 556–568. Dostupné online [cit. 2022-01-21]. ISSN 0261-4189. DOI 10.1038/emboj.2010.346.
- Mitochondrial Diseases [online]. National Library of Medicine - Medical Subject Headings. Dostupné online.
- ZEVIANI, Massimo; DI DONATO, Stefano. Mitochondrial disorders. Brain. 2004-10-01, roč. 127, čís. 10, s. 2153–2172. Dostupné online. DOI 10.1093/brain/awh259.
Externí odkazy
 Obrázky, zvuky či videa k tématu mitochondrie na Wikimedia Commons
Obrázky, zvuky či videa k tématu mitochondrie na Wikimedia Commons - Galerie mitochondrie na Wikimedia Commons
- Působivá animace o mitochondrii; z dílny BioVisions (Harvard University) Archivováno 18. 10. 2010 na Wayback Machine