Endoplazmatické retikulum
Endoplazmatické retikulum (ER) je soustava vzájemně propojených miniaturních membránových cisteren a kanálků, která se nachází v cytoplazmě drtivé většiny eukaryotních buněk. Napojuje se na buněčné jádro a obvykle i na Golgiho aparát.

Endoplazmatické retikulum zvětšuje vnitřní povrch buňky, což má velký význam pro metabolické procesy. Rozlišujeme
- drsné (nebo hrubé) ER, na jehož vnějším povrchu jsou přisedlé ribozomy, a
- hladké ER bez přisedlých ribozomů.
Drsná část endoplazmatického retikula se specializuje na syntézu některých bílkovin a procesy s tím související, jako je skládání těchto proteinů a jejich oligomerizace či navěšování jistých cukerných zbytků na tyto bílkoviny. V drsném ER také probíhá rozklad špatně sbalených či poškozených bílkovin – mechanismus za to zodpovědný se označuje jako ER-asociovaná degradace proteinů.[1]
V hladkém endoplazmatickém retikulu se odehrávají zcela odlišné procesy – odstraňování toxických (odpadních) látek, některé části metabolismu lipidů a metabolismu hemu. Dále se mohou z hladkého ER regulovaně uvolňovat vápenaté ionty.[1]
Historie výzkumu
První pozorování endoplazmatického retikula se uskutečnila již světelným mikroskopem, ale výzkumníci neměli představu o tom, co vlastně pozorují. Příkladem jsou Nisslova tělíska objevená Franzem Nisslem v nervových buňkách v roce 1884.[2] Endoplazmatické retikulum jako takové tak bylo ve srovnání s ostatními organelami objeveno až poměrně pozdě – v roce 1945 – a to díky použití elektronového mikroskopu ke studiu buněk.[3] Práce se jmenovala A Study of Tissue Culture Cells by Electron Microscopy[pozn. 1] a jejími autory byli Keith R. Porter, Albert Claude a Ernest F. Fullam.[4] Bylo popsáno ve fixovaných ptačích buňkách jako systém vzájemně propojených váčků a vláken o velikosti kolem 100–150 mikrometrů. V následujících letech se popisu této struktury věnovali i další studie a v roce 1953 Keith R. Porter poprvé navrhl termín „endoplazmatické retikulum“ („endoplazmatické“ proto, že se nachází ve vnitřní části cytoplazmy (v endoplazmě), „retikulum“ kvůli své komplikované větvené struktuře – latinsky „reticulum“ = síť).[3][5]
Evoluční původ
Endoplazmatické retikulum se vyskytuje v buňkách všech eukaryotických (jaderných) organismů, ačkoliv je někdy velmi redukované a sotva znatelné.[6][7] Společný předek všech eukaryot s nejvyšší pravděpodobností vlastnil funkční systém vnitřních membrán včetně ER. Tomu nasvědčuje i rozšíření proteinových rodin, důležitých pro funkci ER, napříč všemi eukaryotickými organismy. K těmto univerzálně se vyskytujícím bílkovinám patří coat proteiny, SNARE, GAPy a GEFy.[7] O průběhu vzniku endoplazmatického retikula bylo publikováno několik hypotéz, autorem jedné z nich je Thomas Cavalier-Smith. Ten vychází ze skutečnosti, že je ER v podstatě kontinuální s membránou jaderného obalu, a domnívá se, že ER vzniklo vlastně současně s jádrem. Představuje si to tak, že u předka eukaryot byla DNA navázána u cytoplazmatické membrány, následně však v místě, kde byla navázána DNA, došlo k odškrcení této cytoplasmatické membrány a vzniku vnitřního váčku s DNA uvnitř. Membránový váček se na vnější straně postupně nařasil a vznikl systém endoplazmatického retikula. Následně došlo ke specializaci systému jaderné membrány a membrán ER – tyto membrány totiž začaly využívat kotranslační transport pomocí částice SRP, což je něco, co cytoplazmatická membrána neumí. Byly vypracovány teorie, jak mohl tento kotranslační transport vzniknout, když proteiny k němu potřebné jsou zároveň vytvářeny právě jen díky kotranslačnímu transportu.[8]
Stavba
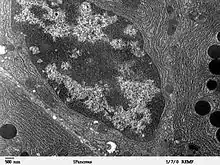
Transmisní elektronový mikroskop
ER je neuvěřitelně dynamický systém vzájemně propojených váčků, v rámci kterého dochází k čilému transportu lipidů a bílkovin. Každou minutu projde endoplazmatickým retikulem mezi 2 a 10 miliony bílkovin[1] a jen málokterá v něm zůstane. Přesto je v dlouhodobém měřítku udržována přibližně konstantní velikost ER a rozložení jeho váčků. Významnou roli v udržování rovnováhy v ER hrají molekulární motory asociované s mikrotubuly, které jsou schopné roztahovat váčky endoplazmatického retikula do požadovaného sektoru cytosolu.[1]
Rozsah
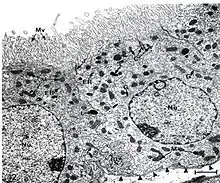
Transmisní elektronový mikroskop
Endoplazmatické retikulum představuje pravděpodobně rozsahem nejvýznamnější část systému vnitřních membrán v eukaryotické buňce (tzv. endomembránového systému).[1] Někdy může zaujímat až 10 % objemu buňky.[9] Je tvořeno trojrozměrným systémem vzájemně propojených trubic či plochých váčků, tzv. cisteren, který může sahat od jádra až k povrchu buňky.[1] Celkový povrch membrán ER např. u jaterních buněk tvoří více než 50 % celkového povrchu buněčných membrán[10] (u exokrinních buněk slinivky dokonce 60 %[10] a v Leydigových buňkách dokonce 71 %[11]). Celkový povrch endoplazmatického retikula v jaterních buňkách činí asi 56 000 μm2, v (objemově mnohem menších) exokrinních buňkách slinivky asi 7 800 μm2[10] a v Leydigových buňkách asi 32 000 μm2.[11] Existují však i opačné extrémy: ve zralých červených krvinkách se endoplazmatické retikulum nevyskytuje, je vypuzeno společně s jádrem v průběhu zrání červených krvinek.[12] Někdy se udávalo, že ER chybí také u prvoka E. histolytica,[13] případně, že funkci ER zastává systém izolovaných váčků. Pomocí pokročilých fluorescenčních technik (GFP fúzní protein a následné FLIP) však bylo u tohoto prvoka nakonec nalezeno i opravdové membránové retikulum.[14]
Významné rozdíly v rozsahu endoplazmatického retikula jsou patrné při srovnávání různých buněčných typů v rámci lidského těla. Je známo, že například pankreatické acinární buňky, fibroblasty a plazmatické buňky mají obrovské drsné endoplazmatické retikulum. To souvisí s jejich vysokou proteosyntetickou aktivitou, tzn. vyrábí spoustu proteinů, ať už to jsou trávicí enzymy v případě acinárních buněk, kolagen v případě fibroblastů či protilátky v případě plazmatických buněk. K velkému rozvoji hladkého endoplazmatického retikula dochází v buňkách, které vyrábí steroidy (např. v buňkách kůry nadledvin), dále také v hepatocytech, ale i ve svalových buňkách, kde se označuje jako sarkoplazmatické retikulum.[15]
Membrána
Cisterny endoplazmatického retikula jsou obaleny buněčnou membránou s charakteristickým složením. Například membrána ER v potkaních hepatocytech obsahuje z 53 % fosfatidylcholin, z 20,2 % fosfatidylethanolamin, z 3,7 % sfingomyelin a zbývajících 14,1 % zaujímají fosfatidylserin a fosfatidylinositol.[16] Podobná čísla (s menšími odchylkami) platí i např. pro kořenové buňky ječmene. U kvasinky Saccharomyces cerevisiae je složení rovněž podobné, jen obsahuje o něco více fosfatidylethanolaminu (33,4 %) na úkor fosfatidylcholinu.[16] V membráně ER se udávají i stopová množství signálních fosfolipidů (DAG, CDP-DAG, PA a lysofosfolipidů) a také dolicholu. Membrána ER je v porovnání s ostatními membránami extrémně chudá na steroly (jako je cholesterol) a složitější sfingolipidy. Absence cholesterolu je způsobena tím, že tento sterol je sice v endoplazmatickém retikulu syntetizován, ale následně je velmi rychle transportován do jiných organel.[17]
Lumen
Lumen (dutina) endoplazmatického retikula má charakteristické chemické složení, jež toto prostředí odlišuje od cytosolu. Mluvíme o proteomu a metabolomu endoplazmatického retikula. Panují zde zvláštní redoxní podmínky, za kterých mohou vznikat disulfidické můstky (oxidací SH skupin), na druhou stranu se v endoplazmatickém retikulu pyridinové nukleotidy (nutné pro řadu metabolických drah) udržují v redukovaném stavu.[18] Další anomálií je velmi vysoká koncentrace vápenatých iontů: ta je v některých buňkách až 10 000× vyšší, než v cytosolu (cca 1 mM ve srovnání s 0,1 μM v cytosolu).[19] Naopak pH v endoplazmatickém retikulu je v podstatě neutrální, srovnatelné s cytosolem.[20][pozn. 2]
Funkční části ER
Endoplazmatické retikulum se skládá z několika morfologicky odlišitelných částí, které však mezi sebou volně komunikují a nedá se např. zamezit difuzi proteinů přítomných uvnitř ER, aby rovnoměrně vyplnily celý prostor ER. Nejčastěji uváděnými částmi ER jsou drsné endoplazmatické retikulum (RER, z angl. rough) a hladké endoplazmatické retikulum (SER, z angl. smooth). Základní rozdíl mezi drsným a hladkým ER spočívá v přítomnosti ribozomů na membráně drsného endoplazmatického retikula, zatímco v hladkém tyto struktury chybí.
Některé studie mají tendenci uvádět jako třetí část ER ještě jaderný obal, který se od ER liší přítomností jaderných pórů a proteinů, které se vážou na chromatin a jadernou laminu.[1] Čtvrtou část ER mohou u rostlin představovat desmotubuly endoplazmatického retikula, procházející skrz plazmodezmata mezi buňkami.[21] William C. Pollard tyto části označuje jako subdomény a přidává k nim ještě „exportní zónu ER“ (z níž se váčky odštěpují do sekretorické dráhy) a „kontaktní zónu ER“ (kde dochází k výměně lipidů s jinými membránovými organelami).[1]
Funkce
Drsné ER
Význam drsného endoplazmatického retikula spočívá především ve výrobě a úpravě bílkovin, jejich transportu a rozkladu.
Syntéza bílkovin
V drsném ER dochází k výrobě (translaci) prakticky všech transmembránových bílkovin, tedy bílkovin, jež jsou trvale zabudovány do některé z buněčných membrán. Mohou to být jak proteiny určené pro cytoplazmatickou membránu, tak i např. pro membránu Golgiho aparátu, buněčného jádra, endozomů, lyzozomů, ale i pro membránu samotného endoplazmatického retikula. Mimo to jsou zde vyráběny bílkoviny, které se vyskytují uvnitř ER, Golgiho aparátu, endozomů, lysozomů a dalších složek sekreční dráhy. Konečně zde dochází k vytváření většiny proteinů, které jsou z buňky následně uvolňovány ven, do mimobuněčného prostoru.[1]
Translace na drsném endoplazmatickém retikulu je jedna ze dvou hlavních možností, jak může v buňce k syntéze bílkovin docházet (druhý způsob je translace na volných ribozomech v cytosolu). Translace proteinů určených pro ER začíná rovněž v cytosolu, ale následně jsou rozeznány jisté signální sekvence nacházející se na začátku vyráběného proteinu (k rozeznání dojde pomocí signál rozpoznávající částice, SRP), načež je protein naveden na SRP receptor na membráně endoplazmatického retikula a translace je znovu spuštěna. Díky SRP receptoru dojde k vazbě na speciální membránový kanál, skrz který může být polypeptid soukán do lumen endoplazmatického retikula. Existují i speciální mechanismy, jak například vyrobit bílkoviny, jež membránu překračují několikrát.[19]
Úpravy proteinů
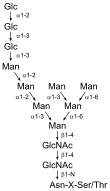
Uvnitř endoplazmatického retikula se nachází velké množství enzymů, které usnadňují skládání bílkovin, jež jsou translokovány do retikula. Jedním z těchto enzymů je proteindisulfidizomeráza (PDI), která usnadňuje vznik SS můstků. Bílkoviny vyráběné v ER se od těch cytosolických liší mimo jiné tím, že mohou mít SS můstky, protože v lumen ER jsou pro to dostatečně oxidativní podmínky.[pozn. 3] Další bílkovinou, která se účastní skládání bílkovin, je BiP protein. Tento chaperon rozeznává hydrofobní oblasti na povrchu špatně složených proteinů a pokouší se přimět tyto proteiny, aby se složily správně.[19]
V endoplazmatickém retikulu probíhá i část tzv. „posttranslačních modifikací“, konkrétně glykosylace, tedy výroba glykoproteinů. Většina proteinů přítomných v endoplazmatickém retikulu i v jiných částech sekreční dráhy jsou ve skutečnosti (N-)glykoproteiny. V endoplazmatickém retikulu se na proteiny přidává poměrně komplikovaný větvený oligosacharid (tzv. prekurzorový oligosacharid), který je pro tento účel předpřipraven v membráně (navázaný na dolichol). V následujícím kroku je část monosacharidů promptně odstraněna. Ačkoliv většina glykosylační rozmanitosti vzniká až v Golgiho aparátu, i glykosylace v ER má zřejmě důležitou funkci. Na glykosylované proteiny se totiž vážou chaperony kalnexin a kalretikulin, které drží špatně sbalené proteiny v ER do té doby, než se správně uspořádají.[19]
Dalším důležitým procesem, který probíhá v endoplazmatickém retikulu, je připevňování vybraných membránových proteinů na GPI kotvu, která v konečném důsledku umožňuje periferní připevnění bílkovin na vnější list cytoplazmatické membrány.[19]
Degradace proteinů
Pokud se nepodaří sbalit protein do prostorového uspořádání, které mu přísluší, nabízí se možnost tento protein degradovat (rozložit), aby v endoplazmatickém retikulu nepřekážel či neškodil. Dráha, která toto zjišťuje, se označuje „ERAD“ – z angl. ER-associated degradation, tedy degradace asociovaná s endoplazmatickým retikulem. Celý proces začíná rozpoznáním špatně složeného proteinu. Následně je takový protein transportován (retrotranslokován) do cytosolu, kde je ubikvitinován. Ubikvitinace v podstatě říká, že daný protein má být v proteazomu rozložen na základní stavební kameny.[1]
Pokud se v endoplazmatickém retikulu vyskytuje příliš mnoho špatně složených bílkovin, značí to, že je něco v nepořádku s enzymatickou mašinérií chaperonů a dalších proteinů podílejících se na skládání proteinů. Za těchto podmínek se spustí tzv. unfolded protein odpověď (UPR), která mění expresi několika stovek genů a způsobí, že je posléze ER výkonnější a schopnější, co se týče skládání proteinů.[1]
Hladké ER
V hladkém endoplazmatickém retikulu dochází k širokému spektru různých anabolických či katabolických reakcí. Soustředí se zde i enzymy schopné zneškodňovat toxické látky. Hladké ER je mimoto jakýmsi vnitrobuněčným skladem vápenatých iontů. Dochází zde také k tvorbě tuků a cukrů.[zdroj?]
Metabolické dráhy
V endoplazmatickém retikulu (zejména v tom hladkém, kde není kladen důraz na syntézu bílkovin) se odehrává řada poměrně významných metabolických pochodů. Přímo uvnitř (v lumen) hladkého ER dochází například k hydrolýze glukóza-6-fosfátu, oxidaci a redukci různých steroidních hormonů, syntéze vitamínu C[22] a k některým krokům rozkladu hemu,[23] probíhá zde i desaturace mastných kyselin, jeden z typů elongace mastných kyselin či třeba jejich ω-oxidace.[24] Na vnitřní straně membrány ER probíhá např. část syntézy cholesterolu nebo ceramidu.[1] Celá řada dalších pochodů probíhá na vnější straně membrány endoplazmatického retikula, namátkou syntéza fosfolipidů[25] či výroba acyl-CoA pomocí acyl-CoA syntetázy.[26]
Detoxifikace
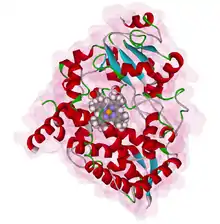
V membráně endoplazmatického retikula se vyskytuje skupina cytochromů P450,[1] proteinů schopných přidávat různé funkční skupiny na různé mastné kyseliny, steroidy a prostaglandiny, ale také na cizorodé látky včetně léků, organických rozpouštědel, pesticidů a karcinogenů. Tím dochází k jejich biotransformaci, načež mohou být např. léky následně vyloučeny z těla.[27] Jedním z cytochromů P450 přítomných v endoplazmatickém retikulu je cytochrom P450 3A4 (CYP3A4),[28] který metabolizuje celou polovinu všech známých léků (namátkou nifedipin, cyklosporin, erytromycin, gestoden či aflatoxiny).[27]
Skladování a uvolňování Ca2+
Jednou z důležitých rolí endoplazmatického retikula je skladovat vápenaté ionty (Ca2+) a v případě potřeby je řízeně uvolňovat do cytosolu. Hladina vápníku v cytosolu má totiž zásadní vliv na průběh celé řady procesů v buňce a je nutné ji udržovat pod striktní kontrolou. V endoplazmatickém retikulu dosahuje koncentrace vápenatých iontů asi 1–3 mM (cca stejně jako vně buňky), tedy mnohokrát více než v cytosolu. Většina vápenatých iontů v ER je navázána na Ca2+ vazebné proteiny, jako je např. kalretikulin, GRP94, BiP či kalsekvestrin. Malá část (asi 0,1–0,4 mM) je volně v lumen ER (nenavázána na proteiny).[29] Vápenaté ionty se do ER pumpují pomocí SERCA ATPázy a uvolňují se (po přijetí urč. signálu) skrz kanály v inositoltrifosfátových receptorech či v ryanodinových receptorech.[29] Hladké endoplazmatické retikulum ve svalových buňkách (tzv. sarkoplazmatické retikulum) se specializuje právě na uvolňování vápenatých iontů: bez otevírání vápníkových kanálů by nemohlo docházet ke svalovému stahu.[15]
ER jako součást sekrečního systému
Z ER vychází množství váčků do jiných organel, jiné naopak s ER splývají. ER je první zastávkou v tzv. sekrečním systému, který umožňuje transport látek směrem k plazmatické membráně a ven z buňky. Bílkoviny, které mají v ER zůstat, musí být opatřeny značkou KDEL. Pro většinu bílkovin přítomných v ER je endoplazmatické retikulum pouze první zastávkou na jejich pouti buňkou. Tyto bílkoviny se shromažďují v „ER exportních doménách“ a pomocí speciálních signálních motivů na svém povrchu se váží na celou řadu pomocných proteinů a nakonec pučí ven ve váčcích obalených pláštěm z COPII komplexů. Váčky směřují ke Golgiho aparátu, a to podél speciálních „kolejnic“ tvořených mikrotubuly. Byly však prokázány i váčky, které se od Golgiho aparátu vrací zpátky k endoplazmatickému retikulu (jsou tzv. „retrográdní“). Jsou důležité např. pro recyklaci „omylem“ odeslaných bílkovin z Golgiho aparátu zpět do ER. Tyto váčky pučí díky plášťovému proteinu COPI.[1]
Odkazy
Poznámky
- česky „Studium buněk z tkáňových kultur pomocí elektronové mikroskopie“
- V dalších částech sekrečního systému, jako je Golgiho aparát či lyzozomy, je pH kyselé.[20]
- Oxidativní podmínky v ER jsou způsobeny převážně tím, že poměr glutathion:glutathiondisulfid (GSH:GSSG) je v ER asi stokrát nižší než v cytosolu.[9]
Reference
- POLLARD, Thomas D; EARNSHAW, William C. Cell Biology. 2. vyd. [s.l.]: Saunders, 2007. 928 s. ISBN 1416022554.
- KIERNAN, John A. Staining Sections of the Central Nervous System [online]. [cit. 2011-11-05]. Dostupné v archivu pořízeném dne 2012-05-13.
- PALADE, G. E.; PORTER, K. R. Studies on the endoplasmic reticulum. I. Its identification in cells in situ. J Exp Med.. 1954, roč. 100, čís. 6, s. 641–56. Dostupné online. ISSN 0022-1007.
- PORTER, Keith R.; CLAUDE, Albert; FULLAM, Ernest F. A Study of Tissue Culture Cells by Electron Microscopy. The Journal of Experimental Medicine. 1945-03-01, roč. 81, čís. 3, s. 233–246. Dostupné online. ISSN 0022-1007.
- PORTER, Keith R. Observation on a Submicroscopic Basophilic Component of Cytoplasm. The Journal of Experimental Medicine. 1953-05-01, roč. 97, čís. 5, s. 727–750. ISSN 0022-1007.
- MIRONOV, A. A.; BANIN, V. V.; SESOROVA, I. S., et al. Evolution of the endoplasmic reticulum and the Golgi complex. Adv Exp Med Biol.. 2007, roč. 607, s. 61–72. Dostupné online. ISSN 0065-2598.
- DACKS, J. B.; FIELD, M. C. Evolution of the eukaryotic membrane-trafficking system: origin, tempo and mode. J Cell Sci.. 2007, roč. 120, čís. Pt 17, s. 2977–85. Dostupné online. ISSN 0021-9533.
- HAMPL, Vladimír. Vznik eukaryotické buňky - Eukaryogeneze (protistologie 2011) [online]. [cit. 2011-11-06]. Dostupné v archivu pořízeném dne 2012-03-14.
- CSALA, M.; BÁNHEGYI, G.; BENEDETTI, A. Endoplasmic reticulum: a metabolic compartment. FEBS Lett.. 2006, roč. 580, čís. 9, s. 2160–5. Dostupné online. ISSN 0014-5793.
- Alberts, Bruce , et al. The Molecular Biology of the Cell. [s.l.]: Garland Science, 2002. (4th. ed). ISBN 0-8153-3218-1.
- MORI, H.; HIROMOTO, N.; NAKAHARA, M., et al. Stereological analysis of Leydig cell ultrastructure in aged humans. J Clin Endocrinol Metab.. 1982, roč. 55, čís. 4, s. 634–41. Dostupné online. ISSN 0021-972X.
- HUME, R.; BURCHELL, A.; ALLAN, B. B., et al. The ontogeny of key endoplasmic reticulum proteins in human embryonic and fetal red blood cells. Blood.. 1996, roč. 87, čís. 2, s. 762–70. Dostupné online. ISSN 0006-4971.
- VOLF, Petr; HORÁK, Petr. Paraziti a jejich biologie. Praha: Triton, 2007. (Vyd. 1). ISBN 978-80-7387-008-9. S. 318.
- TEIXEIRA, Jose E.; HUSTON, Christopher D. Evidence of a Continuous Endoplasmic Reticulum in the Protozoan Parasite Entamoeba histolytica. Eukaryotic Cell. 2008-07, roč. 7, čís. 7, s. 1222–1226. ISSN 1535-9778. DOI 10.1128/EC.00007-08.
- JUNQUEIRA, Luiz Carlos; CARNEIRO, Jose. Basic Histology text and atlas. 11. vyd. [s.l.]: McGraw Hill, 2005. Dostupné online.
- LOMIZE, Andrei; LOMIZE, Mikhail; POGOZHEVA, Irina. Membrane Protein Lipid Composition Atlas: Lipid Composition in Endoplasmic reticulum membrane [online]. Dostupné online.
- VAN MEER, Gerrit; VOELKER, Dennis R.; FEIGENSON, Gerald W. Membrane lipids: where they are and how they behave. Nature reviews. Molecular cell biology. 2008-02, roč. 9, čís. 2, s. 112–124. ISSN 1471-0072. DOI 10.1038/nrm2330.
- BÁNHEGYI, Gábor. Endoplasmic Reticulum Redox [online]. SciTopics, Elsevier, 2010. Dostupné online.
- ALBERTS, B, et al. Molecular Biology of the Cell. 5. vyd. [s.l.]: Garland Science, 2007. 1392 s. ISBN 978-0-8153-4105-5.
- PAROUTIS, P.; TOURET, N.; GRINSTEIN, S. The pH of the secretory pathway: measurement, determinants, and regulation. Physiology Bethesda.. 2004, roč. 19, s. 207–15. Dostupné online. ISSN 1548-9213.
- ROBINSON, David G. The plant endoplasmic reticulum. [s.l.]: Springer, 2006. 337 s. Dostupné online.
- NATO Public Diplomacy Division. Endoplasmic reticulum: a metabolic compartment. Příprava vydání Angello Benedetti, Gábor Bánhegyi, Ann Burchell. [s.l.]: IOS Press, 2005. 151 s.
- KIM, H. P.; PAE, H. O.; BACK, S. H., et al. Heme oxygenase-1 comes back to endoplasmic reticulum. Biochem Biophys Res Commun.. 2011, roč. 404, čís. 1, s. 1–5. Dostupné online. ISSN 1090-2104.
- VOET, Donald; VOET, Judith. Biochemie. 1.. vyd. Praha: Victoria Publishing, 1995. ISBN 80-85605-44-9.
- LEHNINGER, Albert L., David L. Nelson, Michael M. Cox. Lehninger principles of biochemistry. 3rd. vyd. New York: Worth Publishers, 2000. Dostupné online. ISBN 1-57259-931-6.
- VOET, Donald; VOET, Judith. Biochemistry. 4. vyd. [s.l.]: John Wiley & Sons, 1990. Dostupné online. ISBN 978-0470-57095-1.
- BERNHARDT, Rita. Cytochrome P-450. In: Lennarz,W.J., Lane, M.D. Encyclopedia of Biological Chemistry , Four-Volume Set, 1-4. [s.l.]: [s.n.]
- Entrez - Gene - CYP3A4 cytochrome P450, family 3, subfamily A, polypeptide 4 [ Homo sapiens ] [online]. NCBI. Dostupné online.
- GROENENDYK, Jody; MICHALAK, Marek. Calcium Buffering Proteins: ER Luminal Proteins. In: Lennarz,W.J., Lane, M.D. Encyclopedia of Biological Chemistry , Four-Volume Set, 1-4. [s.l.]: [s.n.]
Externí odkazy
 Obrázky, zvuky či videa k tématu endoplazmatické retikulum na Wikimedia Commons
Obrázky, zvuky či videa k tématu endoplazmatické retikulum na Wikimedia Commons - (česky) WikiSkripta - Stres endoplazmatického retikula
- (česky) Návod na barvení endoplazmatického retikula pro přípravu mikroskopických preparátů
- (anglicky) Animace zachycující tzv. kotranslační import bílkovin dovnitř ER
- (anglicky) Graficky zpracovaný seznam membránových proteinů endoplazmatického retikula
- (německy) Sbírka elektronových mikrofotografií drsného endoplazmatického retikula
- (německy) Sbírka elektronových mikrofotografií hladkého endoplazmatického retikula