Evoluce člověka
Evoluce člověka (antropogeneze), tedy proces, který podle evoluční teorie vedl ke vzniku druhu Homo sapiens, se odehrál během dlouhého období kenozoika. Na jeho studiu se v současnosti podílí velké množství vědních oborů – vedle paleoantropologie je to například primatologie, geologie (stratigrafie), klimatologie nebo embryologie. V poznání mladších fází antropogeneze pomáhá také archeologie; velký posun v novější době umožnil rozvoj genetiky.
V širším slova smyslu není otázka antropogeneze rozšířena pouze na fylogenetický vývoj rodu Homo a jeho předchůdce, ale také na postupné osídlování Země moderními lidmi i na současné tendence v evoluci člověka, vliv zemědělství nebo industrializace.
Dějiny studia
Snahy poznat vlastní původ je možné předpokládat již v pravěku, neboť představy o vzniku člověka jsou součástí nejstarších mýtů. Počátky moderního vědeckého studia je však možné datovat až do 18. století.
Již Carl Linné postavil člověka pod druhovým označením Homo sapiens do blízkosti moderních lidoopů. V jeho monumentálním díle ovšem nešlo o blízkost danou vývojem, ale morfologickou podobností.

Zásadním impulsem k hledání lidských předků se staly práce o evoluci, které vydali Jean Baptiste Lamarck, Alfred Russel Wallace a Charles Darwin v 19. století. O společném předku lidoopů a lidí poprvé otevřeně napsal Thomas Huxley v roce 1863. Tyto myšlenky však zůstávaly ještě dlouho kontroverzní a narážely i v rámci vědecké komunity na hluboce zakořeněné náboženské představy. Ačkoliv již v této době byly známy nálezy vyhynulých zvířat i hominidů, byly všeobecně pokládány za kuriozity a postižené jedince, staré maximálně několik set let, případně za doklady života před biblickou potopou.
Přesto již v roce 1856, tedy tři roky před zveřejněním Darwinovy evoluční teorie, oznámil německý profesor Schaffshausen nález neandrtálského člověka a opatrně jej popsal jako „nižší typ, než kterákoliv ze současně žijících lidských ras“. Ostatní badatelé ovšem objev považovali za ostatky málo inteligentního či přímo tělesně i duševně postiženého člověka.[1]
S postupným přijetím evoluční teorie během druhé poloviny 19. století byly již nalezené ostatky hominidů bez pochybností řazené k předkům moderního člověka. Začalo hledání kolébky lidstva v Evropě, Africe i Asii, zpočátku však bylo silně ovlivněné podvrhem Piltdownské lebky. Mezi nejznámější paleoantropology první poloviny 20. století patří Davidson Black, Eugène Dubois, Raymond Dart nebo manželé Mary a Louis Leakeyovi.
K prvním významnějším objevům došlo v 90. letech 19. století na Jávě (Homo erectus), protože část odborníků na rozdíl od Darwina hledala kořeny lidstva v Asii. Teprve kolem poloviny 20. století se zájem odborníků upřel na Afriku. Přestože zde první doklady lidských předků zachytil již v roce 1924 Raymond A. Dart (Australopithecus africanus), většina odborníků tyto nálezy ignorovala.
V 60.–70. letech 20. století došlo k mnoha významným objevům ve východní Africe, kde byly zachyceny ostatky australopitéků i nejstarších lidí (zejm. Australopithecus afarensis, Homo habilis). Definitivně tak bylo potvrzeno, že kořeny rodu Homo jsou africké. Zásadní nálezy kosterních ostatků i nejstarších kamenných nástrojů byly učiněny v Olduvajské rokli. Mezi další významné lokality patří Hadar v Etiopii a Laetoli v Tanzanii, které potvrdily v té době ještě nejistou domněnku, že pliocenní homininé již chodili vzpřímeně po dvou končetinách jako moderní lidé. Množství nálezů bylo získáno také v okolí jezera Turkana – na dolním toku řeky Omo na jihu Etiopie a v oblasti Koobi Fora v Keni.
Rychle po sobě následující objevy nejstarších homininů ve východní a střední Africe (Sahelanthropus, Orrorin, Ardipithecus) na přelomu 20. a 21. století poodhalily dosud nejstarší a málo poznané základy lidského rodu. Nálezy druhů Sahelanthropus a Australopithecus bahrelghazali ve středoafrickém Čadu přitom ukázaly, že vývoj homininů neprobíhal pouze ve východní a jižní Africe. Také moderní objevy v jihoafrických krasových jeskyních (Australopithecus sediba, Homo naledi) naznačují, že lidská evoluce byla poměrně složitým procesem.
I přes neustálé nové a často překvapující nálezy stále není fylogeneze člověka v mnoha ohledech vyřešena. Nový impuls v paleoantropologii přinesla moderní genetika a metoda molekulárních hodin. I ta má však svoje omezení, a tak četné otázky dosud zůstávají nezodpovězeny.
Novější data stále častěji ukazují na skutečnost, že evoluce homininů nebyla lineární a nevedla jednoduše od opic a lidoopů k modernímu člověku. Zdá se, že vývojový strom se spíše podobal hustému keři s mnoha slepými větvemi, kde pouze druh Homo sapiens dosáhl současnosti. Nejpozději v pliocénu však lze sledovat větší množství druhů, které se vyskytovaly současně. Tito vyhynulí hominini přitom často přežívali po mnohem delší dobu než současný Homo sapiens, byli tedy z evolučního hlediska velmi úspěšní.[2]
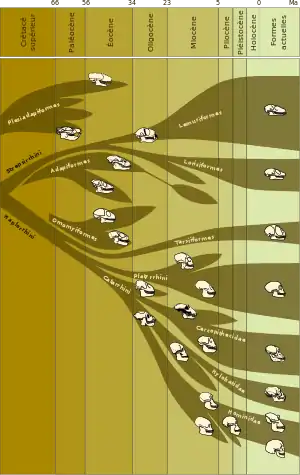
Předchůdci rodu Homo
První primáti


Již v triasu, zhruba před 220 miliony let, se ze savcovitých plazů (Synapsida) vyvinuli první savci. Byli to velmi drobní a nenápadní tvorové. Nejmenší z nich dosahovali váhy jen asi 2 gramy a ti největší, dlouzí okolo 14 cm, vážili asi 30 gramů. Museli přežít v prostředí, jemuž dominovali velcí plazi, a proto se specializovali na noční život a lov hmyzu. Byli teplokrevní, což jim během chladnějších nocí poskytovalo oproti plazům výhodu a volný prostor k obživě. Měli ale i další zajímavé vlastnosti – několik odlišných typů zubů, tvrdé patro, lebkou dobře chráněný a v poměru k velikosti těla výrazně větší mozek, srst, dobrý sluch a jiné. Aby zvýšili šance na přežití u velmi malých, křehkých mláďat a zamezili tak energeticky náročné potřebě plození velkého množství potomků, přešli od kladení vajec k rození živých mláďat, živených mateřským mlékem.
V paleocénu (před 65–55 miliony let) došlo k obrovskému rozvoji savců a mezi nimi se objevili i první primáti. Pohybovali se nejvíce v korunách stromů a jejich zadní nohy byly proto vybavené typickým silným palcem v opozici k ostatním prstům. Oproti jiným savcům se u nich vyvinul velmi dobrý zrak a oči umístěné vpředu, nikoliv po stranách hlavy. Byli tedy schopní stereoskopického vidění. Rostlinnou stravu dobře zpracovávali díky nově utvářeným stoličkám s velkou třecí plochou. Ti, kteří se i nadále živili hmyzem, měli stoličky ostřejší, s výraznějšími hrbolky. Primáti také rodili menší, ale vyvinutější mláďata, která již nebyla slepá, hluchá a bez srsti. Samice měly většinou pouze jedno mládě a jen jeden pár mléčných žláz.
Takzvaní vyšší primáti se zhruba před 35 miliony lety, tedy koncem eocénu, rozdělili na dvě hlavní větve. Jižní Ameriku osídlily takzvané opice Nového světa (Ploskonosí), zatímco v Africe se vyvíjely opice Starého světa (Úzkonosí). Po oddělení Afriky a Jižní Ameriky asi před 25 miliony let se pak obě větve utvářely zcela samostatně. Z úzkonosých opic Starého světa se zhruba ve stejné době oddělila nadčeleď Hominoidi (Hominoidea), do které dnes patří gibonovití a hominidé, včetně člověka.[3][4]
| Nadčeleď | Čeleď | Podčeleď | Tribus | Zástupci |
|---|---|---|---|---|
| Hominoidi (Hominoidea) | Gibonovití (Hylobatidae) | gibon (Hoolock, Hylobates, Nomascus), siamang (Symphalangus) | ||
| Hominidé (Hominidae) | Orangutani (Ponginae) | orangutan (Pongo) | ||
| Homininae | Gorillini | gorila (Gorilla) | ||
| Hominini | člověk (Homo), šimpanz (Pan) |


Úzkonosí primáti oligocénu
- Aegyptopithecus je asi nejznámější zástupce oligocénních primátů. Je jedním z předků všech dnešních úzkonosých opic včetně člověka. Žil zhruba před 35–28 miliony let v provincii Fajjúm v dnešním Egyptě. Dosahoval velikosti asi 60–70 cm a hmotnosti 4–6 kg.[5]
- Saadanius byl objeven teprve v roce 2009 a dosud je znám pouze z jediného zlomku lebky. Podle všech znaků je však mnohem blíže poslednímu společnému předku současných úzkonosých opic než Aegyptopithecus. Žil v oligocénu, zhruba před 29–28 miliony let v okolí dnešní Mekky v Saúdské Arábii. Mohl dosahovat značné velikosti, zhruba 15–20 kg. Žil patrně jako příbuzné rody v korunách stromů, kde se pohyboval pomocí všech čtyř končetin.[6]
- Kamoyapithecus je obtížně zařaditelný úzkonosý primát, žijící koncem oligocénu (před 24,2–27,5 miliony let) v severní části dnešní Keni. Je nejstarším známým primátem, u nějž by se dalo uvažovat o příslušnosti k nadčeledi Hominoidea. Nálezů je však zatím velmi málo, a proto je jeho zařazení nejasné. Odhadovaná váha Kamoyapitheca je 30–40 kg.[7]
Miocenní hominoidi
Teprve počátkem miocénu (23 až 5 milionů let před současností) se v paleontologickém záznamu objevují první jistí zástupci hominoidů. Toto geologické období lze ovšem označit jako zlatý věk hominoidů, protože během něj došlo k velkému rozvoji celé nadčeledi. Problematiku oddělení od úzkonosých opic (Cercopithecoidea) ztěžuje nedostatek fosilií z období před 23–32 miliony let, špatně dochované ostatky i vzájemná podoba jednotlivých rodů.[3]
Moderní hominoidi jsou uzpůsobeni k životu v korunách stromů tropického deštného lesa, kde již neběhají po všech čtyřech, ale skáčou a ručkují. Mají proto odlišně utvářené končetiny a celý hrudník. Zcela zřetelným znakem současných hominoidů je chybějící ocas, který již nemusí vyvažovat rovnováhu těla v běhu po čtyřech. Typické jsou pro ně i stoličky s pěti hrbolky (tak zvaný Y-5 neboli dryopitekový vzor) a zubní vzorec 2.1.2.3 (2 řezáky, 1 špičák, 2 třenové zuby a 3 stoličky). I mozek hominoidů je mnohem rozvinutější než u ostatních savců.[8][9]
Nejstarší hominoidi z časného miocénu však sdíleli jen některé z těchto znaků. Nebyli ještě uzpůsobeni šplhu a brachiaci a pohybovali se ponejvíce po všech čtyřech končetinách.[8] Zuby se už ale blížily podobě chrupu u dnešních lidoopů. Jejich zástupci, představovaní především úspěšnou čeledí Proconsulidae, jsou známí výhradně z Afriky (Keňa, Uganda).[10]
Ve středním miocénu se hominoidi rozšířili také do Evropy a Asie, kde došlo k jejich bouřlivému rozvoji. Naproti tomu doklady z Afriky jsou v této době vzácné.[11] Dříve hojně rozšíření proconsulidé ustupují měnícím se klimatickým podmínkám.[11]
Zhruba před 18–20 miliony let došlo podle analýzy DNA k oddělení gibonů,[12] ovšem jejich předci dosud nebyli identifikováni a všichni uvažovaní kandidáti (např. čeleď Pliopithecidae) byli vždy zamítnuti.[8] Postupně se oddělovaly další evoluční linie, vedoucí k dnešním orangutanům (asi před 13–16 miliony let) a gorilám (před 8–9 miliony let). Stejně jako v případě gibonovitých není zcela jasné, zda tato štěpení proběhla v Africe, Evropě nebo v Asii.
V mladším miocénu se hominoidi rozvíjejí opět spíše v Africe, zatímco v Evropě a ve většině Asie postupně mizí. Dosud není jisté, zda v Africe na počátku středního miocénu téměř nebo zcela vyhynuli a byli později převrstveni eurasijskými formami[3] nebo zda hlavní vývojová linie směrem k dnešním lidoopům pokračovala nerušeně v Africe, avšak v oblastech, které neměly vhodné podmínky pro fosilizaci.[13] Jednotlivé rody z Afriky, Evropy i Asie se spolu mohly různým způsobem prolínat a v současnosti nelze určit, která z linií vymřela bez potomků a z které vzešli předci moderních hominidů.
Do konce miocénu většina hominoidů vyhynula. Vymírání úspěšné nadčeledi je dáváno do souvislosti s dalším vysušováním a ochlazováním klimatu a ústupem subtropického klimatického pásu blíž k rovníku.[14] Geografické i klimatické změny však na druhou stranu mohly v Africe stimulovat rychlý rozvoj přizpůsobivějších forem, aby se v závěru miocénu objevili první bipední tvorové, směřující ke vzniku rodu Homo.[14]


| Rod | Stáří | Popis a druhy |
|---|---|---|
| Proconsul | 14–22 milionů let | Většinou odborníků je řazený již mezi hominoidy. Tento až nadprůměrně známý primát obýval po celý časný miocén oblast západní Keni. Vyčleňováno je několik druhů, jejichž velikost se značně liší (cca 10–90 kg). Proconsul již neměl ocas, ale stále ještě běhal po všech čtyřech končetinách. Je používán jako modelový příklad vzhledu společného předka moderních hominoidů.[13]
Proconsul africanus, Proconsul gitongai, Proconsul heseloni, Proconsul major, Proconsul meswae, Proconsul nyanzae |
| Afropithecus |
14–18 milionů let | Tvoří jakýsi přechod mezi časně miocenními zástupci čeledi Proconsulidae a hominidy mladšího miocénu. Ukazují na utváření nové, modernější skupiny hominoidů na přelomu staršího a středního miocénu, přizpůsobené novým podmínkám suššího podnebí. Její zástupci již nesli typické pokročilé znaky, zejména robustní čelisti a zuby se silnou sklovinou, uzpůsobené pro konzumaci tužší potravy a také náznaky adaptace pro pozemní lokomoci, vzpřímenější postoj či pohyb v závěsu. Přesto se stále jednalo o kvadrupední tvory, žijící převážně v korunách stromů. Někteří z nich mohli zahájit expanzi hominoidů mimo Afriku.[15]
Afropithecus turkanensis, Equatorius africanus, Kenyapithecus wickeri |
| Sivapithecus | 8,5–12,5 milionů let | Značně úspěšný rod hominidů obýval ve středním a mladším miocénu oblast Sivalických hor v severním Pákistánu a Indii. Dosahoval váhy asi 35 kg. Je nejstarším známým zástupcem vývojové linie orangutanů (Ponginae). Byl schopen obratného lezení ve větvích stromů, ale ještě nedokázal v závěsu na pažích ručkovat tak, jako moderní orangutani.[16]
Sivapithecus indicus, Sivapithecus parvada, Sivapithecus sivalensis |
| Dryopithecus | 9,5–12 milionů let | Úspěšný rod, obývající ve středním a mladším miocénu rozsáhlé oblasti Evropy byl v mnoha ohledech podobný moderním africkým hominidům. Relativně dobře prozkoumaný tvor ukazuje, jak asi mohl vypadat společný předek goril, šimpanzů a člověka. Různé druhy mohly vážit mezi 15–45 kg. Lehce stavěné čelisti a stoličky se slabou sklovinou byly uzpůsobené ke konzumaci měkkého ovoce. Dlouhé přední končetiny s pohyblivými klouby a silnými zakřivenými prsty dovolovaly obratné ručkování ve větvích stromů.[17]
Dryopithecus brancoi, Dryopithecus crusafonti, Dryopithecus fontani, Dryopithecus laietanus |
| Pierolapithecus | 11,9 milionů let | Jeden z nejstarších známých hominidů žil koncem středního miocénu na Pyrenejském poloostrově. Ve větvích stromů se již místo běhu po čtyřech končetinách pohyboval obratným lezením, avšak ještě nebyl schopen brachiace. Výjimečně dobře dochovaná kostra ukazuje, že dosahoval výšky asi 100–120 cm a váhy kolem 35 kg.[18]
Pieralopithecus catalaunicus |
| Ouranopithecus | 7–10 milionů let | Jeden z nejmladších zástupců evropských hominoidů, žijící v dnešním Řecku a Turecku, nese množství znaků, kterými se blíží moderním africkým homininům. Často bývá uváděn jako jejich společný předek. Dosahoval váhy asi 70 kg, měl poměrně plochý obličej a výraznější nadočnicové valy. Prvním lidem se blíží nezvykle malými špičáky. Robustní čelisti a široké stoličky se silnou vrstvou skloviny mu umožňovaly konzumovat kromě ovoce i tužší rostlinnou stravu jako hlízy, ořechy a semena. Neobýval již subtropický les, ale otevřenější krajinu, takže se zřejmě pohyboval spíše po zemi než ve větvích stromů.
Ouranopithecus macedoniensis, Ouranopithecus turkae |
| Nakalipithecus | 9,8–9,9 milionů let | Dosud málo známý druh ze svrchního miocénu Keni byl velmi podobný rodu Ouranopithecus. Může být posledním společným předkem goril, šimpanzů a lidí a mohl by naznačovat, že se tyto linie odštěpily v Africe.[19]
Nakalipithecus nakayamai |
Bipední hominidé

Vývojové větve šimpanzů (Panina) a lidí (Hominina) se rozdělily zhruba před 6–8 miliony let,[20] doklady počátků lidské linie jsou však zatím vzácné a v jejich poznání převládá nejistota. Výzkumy ale ukazují, že poslední společný předek obou taxonů se moderním šimpanzům příliš nepodobal a že šimpanzi prošli stejně výrazným vývojem jako lidé.[21] Nejsou tudíž jakousi živou fosilií a přesným obrazem našeho předka, ačkoliv poslední společný předek obou větví vypadal jistě více jako lidoop než jako moderní člověk – blížil se mu velikostí mozku a snad se mohl i podobně chovat.[22] Stopy staré 6 miliónů let byly nalezeny na Krétě.[23]
Nejstarší nálezy, které zřejmě již představují zástupce taxonu Hominina nebo tvory velmi blízké, pochází z přelomu miocénu a pliocénu střední a východní Afriky. Mezi první příslušníky samostatné lidské vývojové linie po oddělení od předků šimpanzů by mohly patřit rody Sahelanthropus, Orrorin a Ardipithecus (ačkoliv Sahelanthropus podle novějších výzkumů mezi hominininy ve skutečnosti nepatřil).[24][25] Všechny se vyznačují především zkracujícími se špičáky, zanikajícím ostřícím komplexem mezi špičákem a třenovým zubem a také prvními adaptacemi k dvojnohé chůzi. Představují přechod od čtyřnohých lidoopů, žijících ve větvích stromů tropického lesa, k dvojnohým tvorům přizpůsobeným otevřenějšímu, méně zalesněnému prostředí.[26]
Pozůstatky těchto archaických homininů byly objeveny teprve na přelomu 20. a 21. století, takže jsou zatím málo početné a nekompletní. Existuje velké množství otázek ohledně jejich taxonomické a fylogenetické pozice. Odborníci nejsou jednotní ani v tom, zda všechny popsané ostatky vůbec patřily homininům a mnozí z nich jeden nebo více druhů z okruhu lidských předků vylučují.[27]
Problémem je i fakt, že nejsou známí prakticky žádní předchůdci moderních šimpanzů či goril, s nimiž by bylo možné nálezy prvních homininů porovnat. První ostatky archaických šimpanzů, představované pouze třemi zuby, byly zachyceny v roce 2004 u jezera Baringo v Keni, jsou však jen asi 545 000 let staré.[28] Zajímavé je, že pocházejí z prostoru typického pro vývoj člověka, mimo areál výskytu současných šimpanzů.
Všechny popsané druhy časných homininů nesou zcela svébytnou mozaiku primitivních i odvozených znaků.[29] Nemohou být ale navzájem porovnány, protože z každého byly nalezeny odlišné partie kostry. Jedinou částí těla, zastoupenou poměrně hojně u všech druhů, jsou zuby. Ty jsou si vzájemně značně podobné. Byl proto vysloven i názor, že by se mohlo jednat o jediný rod s druhy, které na sebe časově navazují a tvoří anagenetickou linii.[30] S touto představou ovšem velká část odborníků nesouhlasí.[20][26]
O způsobu života nejstarších homininů lze říci jen velmi málo. Klíčová je schopnost chodit vzpřímeně po dvou končetinách, ačkoliv chůze těchto tvorů se od bipedie člověka ještě podstatným způsobem lišila a není dosud dobře poznaná.[21] Také je třeba mít na paměti, že ne všichni bipední hominidé museli být lidskými předky. S dvojnohou chůzí mohly „experimentovat“ i jiné druhy, které ovšem vymřely bez potomků (např. Oreopithecus).[31]
I přes adaptace k bipedii si první homininé stále uchovávali některé rysy, potřebné pro obratné lezení ve větvích stromů, kde i nadále trávili velké množství času.[32] Přírodovědné analýzy ukazují, že tito tvorové patrně obývali lesní porosty nebo mozaikovitou krajinu s menšími či rozsáhlejšími úseky lesa, v blízkosti vodních toků a jezer.[33] To odporuje staršímu předpokladu, že bipedie vznikla v otevřené bezlesé krajině, když se museli předci člověka vyrovnat s ústupem lesů a šířením savany vlivem měnícího se podnebí na počátku pliocénu.[34]
Dále jsou u všech časných homininů pozorovány změny na chrupu. Předpokládá se, že zmenšování špičáků, které šimpanzím a gorilím samcům slouží zejména k bojům o samice nebo teritorium, dokládá určité změny ve společenském uspořádání s méně dominantní rolí samců.[26] Objevují se však také názory, že redukce zubů může souviset se změnou stravy vlivem klimatických změn – při ústupu lesů s dostatkem měkkého tropického ovoce a tudíž při zvýšené potřebě žvýkání a drcení tužší stravy (hlízy, semena, aj.) velké přečnívající zuby překážely.[35]
| Rod | Stáří | Popis a druhy |
|---|---|---|
| Sahelanthropus | 7,2–6,8 milionů let | Žil na území dnešního Čadu ve střední Africe, daleko od proslulých nalezišť východní a jižní Afriky. Jeho postavení dosud není jisté, ale je nejstarším známým hominidem, u kterého se předpokládá, že patřil k samostatné lidské vývojové linii (subtribus Hominina). Byl ještě velmi blízko poslednímu společnému předku šimpanzů a lidí. Měřil 115–125 cm a vážil až 45 kg. Měl krátké špičáky a podle pozice velkého týlního otvoru mohl využívat bipední chůzi.[36]
Sahelanthropus tchadensis |
| Orrorin | 6,1–5,7 milionů let | Objeven byl v Keni ve východní Africe. Jeho příslušnost k homininům je odvozována zejména podle nálezu stehenních kostí, které se podobají stehenním kostem člověka. Ostatní části těla ale mají spíše lidoopí ráz. Byl vysoký asi 115–125 cm a těžký 30–50 kg. Zatímco na zemi mohl chodit po dvou končetinách, díky silným pažím a dlouhým zahnutým prstům byl zároveň schopný obratně šplhat ve větvích stromů.[37]
Orrorin tugenensis |
| Ardipithecus | 5,8–4,4 milionů let | Rod původem z dnešní Etiopie (východní Afrika) je dobře známý díky unikátnímu nálezu dobře dochované kostry samice „Ardi“. Jeho zástupci měřili asi 120 cm a vážili kolem 50 kg. Používali dvojnohou chůzi, která se ale lišila od chůze lidské. Zároveň si ještě uchovávali některé adaptace k životu v korunách stromů, jako je chápavý palec na noze, silné paže a dlouhé, zakřivené prsty rukou.[38][39]
Mohl být předkem rodu Australopithecus. Ardipithecus kadabba, Ardipithecus ramidus |

Australopitéci
Australopitékové tvoří značně variabilní skupinu evolučně velmi úspěšných homininů. Obývali po téměř 3 miliony let (v době před 4,2–1,3 miliony let) rozsáhlé oblasti východní, jižní a střední Afriky. Vyskytovali se převážně v tropickém podnebném pásu a jen výjimečně se šířili i do subtropů.[40] Rozeznává se nejméně 9 druhů, přičemž mezi nejznámější a také nejlépe poznané zástupce patří Australopithecus afarensis z východní Afriky a Australopithecus africanus z jižní Afriky.[29] Zástupci s extrémně masivní stavbou zubů a čelistí jsou některými badateli vyčleňováni do samostatného rodu Paranthropus. Současníkem a zřejmě blízkým příbuzným australopitéků je také málo poznaný druh hominina – Kenyanthropus platyops.
Australopitékové s největší pravděpodobností patří k vývojové linii moderního člověka. Dosud však nebylo vyřešeno, na který ze známých druhů (pokud vůbec na nějaký) první lidé navazují, s jistotou lze vyloučit jen paranthropy.[41]
Zatímco hlava se ještě podobala spíše lidoopům, tělo již neslo početné lidské znaky.[42] V porovnání s moderními lidmi byli australopitékové poměrně malí – s výškou 100–160 cm a hmotností 30–50 kg se podobali spíše dnešním šimpanzům.[13] Značné rozdíly ve velikosti nalézaných ostatků ukazují na celkem výrazný pohlavní dimorfismus, kdy samci mohli být proti samicím více než o polovinu větší (asi jako u dnešních goril nebo orangutanů).[43] Typický je vcelku malý mozek, jehož objem se pohybuje při horní hranici rozpětí současných lidoopů (v průměru asi 450 cm³). Proto se nepředpokládá, že by duševní schopnosti těchto časných homininů výrazně přesahovaly možnosti šimpanzů, goril nebo orangutanů.[29] K dalším charakteristickým znakům australopitéků patří i masivní stavba čelistí, zubů a navazujících částí obličeje. Utváření lebky a nápadné úpony odpovídají velmi silným žvýkacím svalům.[13]
Páteř, pánev i stavba nohou odpovídají běžně používané vzpřímené dvounohé chůzi.[44] Bipedie mohla být v porovnání s moderními lidmi nedokonalá, energeticky náročnější,[45] nevhodná pro přesuny na velké vzdálenosti a nebo ve velké rychlosti.[46] Přesto pro australopitéky mohla být plně dostačující.[47] Patrně ji totiž kombinovali s poměrně častým šplháním v korunách stromů, kde mohli hledat potravu i úkryt před dravci.[48] Svědčí pro to některé přetrvávající znaky, zděděné po předcích – dlouhé svalnaté předloktí, pohyblivá ramena i zápěstí, mírně prohnuté prsty schopné pevného úchopu nebo částečně odtažitelný palec na noze.[49]
Je obtížné spojit rod Australopithecus s konkrétním biotopem, protože s přibývajícími nálezy je neustále potvrzována značná ekologická tolerance rodu a schopnost najít obživu v různých typech prostředí od sušší, otevřené savany přes krasové oblasti až po prosvětlené či hustší vlhké lesy, spojované nejčastěji se záplavovým územím řek a jezer.[50] Nové výzkumy tedy příliš nepotvrzují dřívější předpoklady o výhradním sepětí australopitéků se suchými travnatými pláněmi bez stromů. Naopak se zdá, že přednost dávali spíše zalesněnému prostředí s dostatkem vláhy, odkud však postupně pronikali i do otevřené krajiny.[48]

Také strava byla zřejmě silně variabilní, obměňovaná podle aktuálně dostupných zdrojů.[51] Preferováno mohlo být ovoce a další měkké části rostlin, nahrazované pouze v dobách nedostatku jinými zdroji.[52] K rozvoji robustního žvýkacího aparátu tak mohla spíše než denní rutina přispět jen krátkodobá, ale kriticky významná konzumace méně oblíbených zdrojů v nepříznivých obdobích roku.[53] Tuhá a abrazivní potrava, vyžadující masivní žvýkací aparát, mohla patřit k důležitým záložním zdrojům. Nemusela být nutričně nejvýhodnější ani snadno dostupná, ale byl jí dostatek i v době, kdy běžné složky potravy selhávaly. Za velmi pravděpodobný záložní zdroj jsou často považovány různé podzemní zásobní orgány travin nebo šáchorovitých (hlízy, cibule, oddenky).[53] I přes náznaky zájmu o živočišné zdroje zůstávali australopitékové převážně (zřejmě ne výhradně) býložraví. Vzhledem k potravním zvyklostem moderních lidoopů i lidí lze ovšem předpokládat příležitostnou konzumaci hmyzu (zejména termitů), drobných obratlovců nebo vajec. V rozporu s dřívějšími představami se nepředpokládá aktivní lov větších savců.[54]
Na základě znalostí nástrojové činnosti moderních lidoopů, ale i jiných druhů zvířat (vydry, makakové) se všeobecně předpokládá, že také australopitékové mohli využívat jednoduché nástroje – větve, klacíky nebo neupravené valouny. Takové chování lze však jen obtížně doložit, protože případné nástroje prakticky nelze rozeznat.[49]
| Rod | Popis a druhy |
|---|---|
| Australopithecus | Široce rozvětvený rod, známý z východní, jižní i střední Afriky. Jeho zástupci byli asi 100–150 cm vysocí (při váze do 50 kg) a pohybovali se vzpřímeně po dvou končetinách. Žili v ostrůvkovitých lesních porostech a lužních lesích při březích řek a jezer. Patrně ještě hojně využívali stromové patro, ale postupně pronikali i do bezlesých travin. Živili se převážně rostlinnou stravou, ale vyloučit nelze ani konzumaci hmyzu, drobných obratlovců nebo vajec. K získávání potravy mohli užívat jednoduché nástroje – valouny, větve, klacíky.
Australopithecus anamensis, Australopithecus afarensis, Australopithecus africanus, Australopithecus garhi, Australopithecus bahrelghazali, Australopithecus sediba |
| Paranthropus | Skupina australopitéků s extrémně masivní stavbou čelistí a zubů, tzv. robustní australopitékové. Žili v jižní i východní Africe, kde obývali přetrvávající ostrůvky hustší vegetace při březích řek a jezer. Více než starší australopitékové pronikali do otevřené krajiny. Živili se převážně rostlinnou potravou. Představovali slepou vývojovou větev, navazující na starší druhy australopitéků.
Paranthropus aethiopicus, Paranthropus robustus, Paranthropus boisei |
Rod Homo

Znaky
Příslušníci rodu Homo se od australopitéka odlišují těmito znaky:
- cílevědomou výrobou a užíváním nástrojů
- větším mozkem – rozdíl minimálně o 500 cm3[zdroj?] – velikost mozku lidských předků se zvětšovala postupně,[55] což vedlo ke zvýšené potřebě příjmu energie. Jedna ze studií tvrdí, že k tomu byla třeba konzumace tuků spíše než masa samotného.[56] Nebo také delší doba kojení mohla mít vliv.[57] Nicméně je teorie o konzumaci masa dosavadně obecně uznávaný názor.[zdroj?] Člověk byl spíše masožravec.[58] Posledních několik tisíc let se ale mozek evolučně zmenšuje a to patrně z důvodu socializace.[59]
- mnohem menšími zuby a spodní čelistí – při změně stavby mohla hrát velmi důležitou roli příprava potravy nástroji a tepelnou úpravou[60]
- stavbou některých kostí – mají tenčí lebeční kosti, velkou hlavici a kratší krček stehenní kosti a nezešikmenou pánev
Nejstarší nález
- Nejstarší nález rodu Homo pochází z Etiopie (Afarsko) a je přibližně starý 2,8 miliónu let.[61] Byl nalezen na území typickém výskytem australopithéka afarského.
- Nejstarší nález druhu Homo sapiens, tzv. Omo 1, byl nalezen v Etiopii při řece Omo a je starý přibližně 190 tisíc let.
Teorie vzniku
Existují dvě klasické teorie vzniku rodu Homo, ve kterých je klimatická změna impulsem evoluce, ale tato teorie je také zpochybňována:[62]
- Teorie převratných evolučních pulsů (Elizabeth Vrbová, polovina 80. let): V důsledku klimatických změn, které způsobily prudké zmenšení plochy lesů a jejich zachování se jen ve formě izolovaných „lesíků“ roztroušených po savaně se před 2,6 milióny let stali „izolovanými“ i obyvatelé jednotlivých lesíků, a proto časem vzniklo nejen mnoho nových druhů Homo, ale i jiných druhů živočichů. Tuto hypotézu uvítali hlavně zastánci teorie přerušované rovnováhy (punctuated equilibrium), která vysvětluje evoluci střídáním období vývojové stagnace a prudkých evolučních výbuchů.
- Teorie kumulativních ekologických změn (Anna K. Behrensmeyer, 1997): V důsledku klimatických změn, které způsobily před 2,5–1,8 miliony let nástup chladnějšího a suššího podnebí a klimatické výkyvy, došlo ke změně fauny ve prospěch savanových druhů (nahrazení až 60 % suchozemských savců), ke kterým patřili i první představitelé rodu Homo. Tato teorie je založená na výzkumu Turkanské kotliny v Keni.
Variabilita klimatu patrně evoluční změny bezpůsobila.[63]
Vodní opice
Proti těmto dvěma "klasickým" teoriím stojí teorie "vodních opů", jež je oficiální akademickou obcí již desítky let popírána, ovšem konečně začíná být přijímána.[64] Podle ní začal člověk chodit po dvou ne při přechodu do savan, aby snad viděl přes vysokou trávu, ale přechodem ze suchého lesa přímo do vody. I ostatní dnešní opi mohou chodit po zadních, ale činí tak výhradně, zato vždy při překonávání vody. Tím u člověka došlo zároveň ke dvěma změnám:
- Ztratil srst, která by mu při častých přechodech mezi vodou a lesem překážela. Vodní savci mohou a i nemusí mít srst, ovšem v opačném směru implikace funguje: Tvor bez srsti je jistě vodní.[64] Na druhou stranu existuje i mnoho jiných teorií o příčinách zřídnutí srsti člověka: Např. kvůli kožním parazitům nebo jako důsledek sexuálního výběru.[65]
- Získal schopnost vědomě zadržovat a řídit svůj dech, čímž byl umožněn vznik řeči. Pro ostatní živočichy, ryze suchozemské, je dech naopak pouze mimovolní reflex.[64]
Dokonce se u člověka již projevily i další adaptace na vodní prostředí, jako by jeho evoluce prošla vodním obdobím podobně jako např. u slona.[64]
Další teorie
Dále je například uvažován vliv využití ohně.[66] Jistou roli mohla hrát potřeba více pečovat o potomky.[67] Kosmické záření z nedaleké supernovy mohlo ovlivnit prostředí a tak i vývoj člověka.[68]
Členění

Členění je spíše výčtem nálezů než nějakým objektivním způsobem kvantifikovaná hranice. Proto by bylo třeba zavést definice než se spoléhat na historické členění nálezů.[69]
Časné formy rodu Homo lze jen obtížně odlišit od mladších zástupců australopitéků, oba rody navíc žily déle než 1 milion let (v době před 2,8–1,3 miliony let) souběžně vedle sebe.[13]
- blíže nespecifikovatelný nález z Hadaru
- Homo habilis (Člověk zručný, po publikaci nálezu z Dmanisi je od roku 2013 považovaný spíše také za Homo erectus)
- Homo rudolfensis (Člověk východoafrický; původně považovaný za Homo habilis)
- Homo ergaster (Člověk pracující/člověk dělný; původně považovaný za Homo habilis nebo Homo erectus, od roku 2013 opět považovaný za Homo erectus)
- Homo georgicus (Člověk gruzínský; nálezy z let 1999 a 2001 v gruzínském Dmanisi, považovaný za Homo erectus)
- Homo erectus (Člověk vzpřímený)
- H. e. erectus (Člověk vzpřímený trinilský; původně Pithecantropus erectus)
- H. e. modjokertensis (Člověk vzpřímený modžokertský)
- H. e. lantianensis (Člověk vzpřímený lantianský; nálezy z okolí města Lan-tchien, provincie Šen-si, Čína; období před 900–790 tisíci lety)
- H. e. pekinensis (Člověk vzpřímený pekingský; původně Sinanthropus pekinensis)
- Homo antecessor (Člověk předchůdce)
- Homo cepranensis (před 800–900 tis. lety)
- Homo floresiensis (Člověk floreský; nález na ostrově Flores v Indonésii z roku 2003)
- Homo luzonensis[70]
- Homo heidelbergensis (Člověk heidelberský; archaický Homo sapiens v užším smyslu, původně /do poloviny 90. let 20. století/ Homo erectus heidelbergensis, od roku 2013 opět považovaný za Homo erectus)
- H. steinheimensis (Člověk steinheimský; dříve Homo sapiens steinheimensis)
- H. soloensis (dříve Homo javanthropus, Javanthropus soloensis, Homo sapiens soloensis; nález z Jávy v Indonésii)
- H. rhodesiensis (dříve Cyphanthropus rhodesiensis, Homo sapiens rhodesiensis)
- H. cepranensis (sporný nález z roku 1994 z Ceprany v Itálii, někdy považovaný za Homo erectus nebo Homo neanderthalensis)
- Homo helmei (část)
- Homo neanderthalensis (Člověk neandrtálský, neandrtálec, paleoantrop, pračlověk; předtím považovaný za poddruh Homo sapiens – Homo sapiens neanderthalensis)
- Homo naledi (nález z roku 2013)
- Homo sapiens (Člověk rozumný, neoantrop)
- H. s. idaltu (nález z roku 1997)
- H. helmei (část)
- H. s. palestinus
- H. s. sapiens fossilis (Člověk kromaňonský, kromaňonec, člověk předvěký, Homo sapiens fossilis, Neoanthropus fossilis, Homo sapiens diluvialis)
- H. s. sapiens recens (Člověk moudrý; moderní člověk)
- Za samostatný druh zatím nejsou uznáni neandrtálcům blízcí denisované, současníci Homo neanderthalensis i Homo sapiens (genetické analýzy naznačují, že se dokonce křížili s člověkem moudrým, konkrétně s Melanésany).[71][72]
- Homo sapiens sapiens, který se dříve užíval k odlišení od Homo sapiens neanderthalensis, se nyní používá pro poslední vývojovou fázi člověka po kromaňoncovi, která započala před 10 000 lety.
- Homo erectus v širším, tedy starším, smyslu (archantrop, opočlovek) zahrnuje druhy Homo erectus, Homo ergaster, Homo antecessor, Homo georgicus a Homo heidelbergensis (mimo rhodesiensis a cepranensis).
- Menší skupina badatelů se domnívá, že i rod šimpanz (Pan) a rod gorila (Gorilla) by se měly řadit do rodu Homo pro jejich podobnost s člověkem.
- Existuje návrh z roku 1999 (Wood), podle kterého jsou Homo habilis a Homo rudolfensis ve skutečnosti z rodu Australopithecus.
- Někteří badatelé (Mayr 1950, Wolpoff 1994, Jelinek) považují Homo erectus za poddruh Homo sapiens. Coppens například považuje všechny příslušníky rodu Homo za Homo sapiens.
- Homo heidelbergensis a Homo neanderthalensis se často zařazovali jako poddruhy Homo sapiens, ale analýzy mitochondrického DNA z fosílií Homo neanderthalensis naznačují, že rozdíl je dost velký na to, aby tyto případy byly klasifikovány jako samostatné druhy.
- Někteří autoři namísto Homo heidelbergensis rozlišují:
- Homo heidelbergensis v užším smyslu (evropští neandrtálci)
- Homo rhodesiensis (afričtí předchůdci Homo sapiens)
- Někteří badatelé nadále považují Homo neanderthalensis a Homo heidelbergensis za poddruhy Homo sapiens. Příklad – dělení pro Afriku dle Bräuera:
- Archaický Homo sapiens (nebo vyspělý Homo erectus)
- Starší archaický Homo sapiens (Homo heidelbergensis – z Afriky, bez Homo helmei = archaický Homo sapiens v užším smyslu = asi před 500 000–300 000 lety).
- Novější archaický Homo sapiens (Homo helmei = asi před 300 000–100 000 lety).
- Starší moderní Homo sapiens (Homo sapiens – z Afriky, bez Homo helmei = asi před 195 000–75 000 lety; Omo 1)
- (Homo sapiens idaltu patří v tomto schématu buď do bodu II, nebo do bodu III)
- Moderní Homo sapiens
- Archaický Homo sapiens (nebo vyspělý Homo erectus)
Homo habilis
Žil asi před 2,4–1,5 milióny let, byl asi 130 cm vysoký, vážil 30–42 kg. Vyvinul se v jižní a východní Africe v pozdním pliocénu a raném pleistocénu. Lebka byla zaoblenější než u australopithéka, čelo ploché, měl mohutné nadočnicové valy (hlavně u mužů); bradový výběžek nebyl vytvořen. Tradičně se předpokládá, že se mezi 2,5–2 milióny let vyčlenil z australopithéka. Podle posledních nálezů však žil zhruba ve stejné době.[zdroj?] A byl učiněn i nález, kde se na jednom místě nachází Australophitecus, Homo habilis a Homo erectus (Olduvajská jeskyně ve východní Africe). Má menší stoličky a větší mozek než australopithék. Průměrná kapacita mozkovny činila 641 cm3. Měl vzpřímenou postavu a byl dobře přizpůsoben k pohybu po dvou končetinách. Vyráběl nástroje z kamene a možná ze zvířecích kostí.
Homo ergaster
Žil od 1,9 do 1,4 miliónů let před dneškem, je označován také jako Homo erectus ergaster. Vytvořil pěstní klín (-> později nahrazen bambusem). Má již tělo bez ochlupení, objevují se potní žlázy kvůli chlazení organismu. Objevovaly se u něj počátky řeči. Zabýval se lovem menších zvířat a sběrem. Jeho mozek vážil už okolo 1 kg. Stopařství. Šířil se z Afriky dále do Evropy a Asie.
Homo erectus
Žil před 1 900 000–100 000 lety. Rozšířil se do Asie před více než 1 800 000 lety.[73] V raném pleistocénu (před 1,5–1 miliónem let) vyspěl Homo habilis a začal vyrábět komplikovanější kamenné nástroje; tyto a jiné odlišnosti jsou pro antropology dostatečné, aby druh klasifikovali jako nový – Homo erectus. Jeho slavným příkladem je tzv. pekingský člověk (Homo erectus pekinensis); jiné pozůstatky byly nalezeny v Asii (význačná je Indonésie), Africe a Evropě. Mnozí paleoantropologové nyní používají termín Homo ergaster pro neasijské členy této skupiny a rezervují název Homo erectus pouze pro fosílie nalezené v Asii. Toto tvrzení podkládají drobnými rozdíly ve stavbě kostry a zubů, které Homo erectus trochu odlišují od Homo ergaster. Hlava člověka vzpřímeného byla předkloněná, obličej byl mohutný, hrubě modelovaný s malým širokým nosem a se silnými vystouplými čelistmi. Čelo bylo nízké, nazad ubíhající, nadočnicové oblouky byly nadměrně vyvinuty. Čelisti byly silné, spodní čelist byla bez brady. Měl mírně klenutou mozkovnu, kapacita lebeční se pohybovala v rozmezí 750–1 250 cm3. Podle tvaru nalezené stehenní kosti byla jeho postava vzpřímená a dosahovala výšky 165–170 cm. Nicméně nálezy nástrojů v Číně naznačují, že člověk opustil Afriku před více než 2 milióny let.[74]
Člověk pekingský (Homo erectus pekinensis)
Obýval Severočínskou nížinu, naleziště pozůstatků se nachází 44 km jihozápadně od Pekingu – u vesnice Čou-kchou-tien ve vápencových kopcích s mnoha jeskyněmi. Jedna zvlášť velká jeskyně dosahovala původně rozlohy srovnatelné s fotbalovým hřištěm (150 m × 45 m a v jednom místě 36,6 m na výšku). Na severovýchodě měla jeskyně malý vchod. Tuto jeskyni poprvé osídlil lidský rod zhruba před 400 000 lety a osídlení trvalo bez přerušení 200 000 let, až do doby, než byla jeskyně téměř úplně zaplněna odpadky. Toto sídliště pekingského člověka je zapsáno na seznamu památek UNESCO.
V roce 1921 byl jeden zub z této lokality identifikován jako zub primitivního předchůdce člověka. První lebka byla nalezena v roce 1929. Pečlivé vykopávky z let 1921–1937, obnovené roku 1959, dosud odkryly přes sto tisíc kamenných nástrojů, přes stovku zubů, čtrnáct lebek a mnoho dalších kostí celkem asi ze čtyřiceti jedinců druhu Homo erectus, velmi podobného jiným raným formám lidského rodu objeveným na Jávě (1891), v Evropě, na Předním východě a v Africe.
Byli to lidé malého vzrůstu. Pekingský muž měřil na výšku necelých 1,6 metru, pekingská žena kolem 1,4 metru. Měli silné lebeční kosti a ustupující brady, avšak kapacita jejich mozku byla 850–1 300 cm3 (proti 775–900 cm3 člověka z Jávy a 1 350 cm3 raného Homo sapiens). Byli to lovci, rybáři a sběrači a používali oheň; osvětlovali jím jeskyni a upravovali na něm maso – ze sedmdesáti procent se jednalo o vysokou zvěř, v obydlí pekingského člověka však byly nalezeny také kosti levharta, medvěda, šavlozubého tygra, hyeny, slona, nosorožce, velblouda, vodního buvola, divokého prasete a koně. V jeskyni nebyly nalezeny žádné hroby ani kompletní kostry, avšak některé lebky byly proražené, z čehož se usuzuje, že pekingský člověk byl částečný kanibal, nebo alespoň lovec lebek, který si pochutnával na mozku svých obětí. To vše dohromady znamená, jak říká K. C. Chang (1986), že nález pekingského člověka je „obrovským úlovkem paleoantropologie“.
Homo heidelbergensis
Heidelberský člověk žil před 800–300 tisíci lety, je označován také jako Homo sapiens heidelbergensis a Homo sapiens paleohungaricus. Měl tzv. maurskou čelist. Kapacita mozkovny dosahovala 1 200 cm3. Jako první osídlil Evropu, nálezy pocházejí z Německa a Maďarska (Vértesszőlős).
Homo neanderthalensis
Poslední společný předek neandertálců a moderních lidí žil před 700 000 lety a k definitivnímu rozštěpení obou druhů došlo zhruba před 370 000 lety. Protoneandrtálci (před 150 000–120 000 lety) a klasičtí neandrtálci (před 100 000–40 000 lety) měli robustní kostru a byli 160 cm vysocí. První zbytky byly objeveny v roce 1856 ve Feldhofské jeskyni v Neanderově údolí řeky Düssel, východně od Düsseldorfu v Německu; nejvýznamnějšími nalezišti v České republice a na Slovensku jsou Gánovce u Popradu, Šipka u Štramberka a Moravský kras. První nálezy byly původně popsány jako Homo neanderthalensis King, 1864.
Jde o vyhynulou či asimilovanou vývojovou větev. Vědci dodnes nedokázali přesněji určit důvod vymizení této větve. Člověk neandrtálský žil v období před 250–30 tisíci lety, je označován také jako Homo sapiens neanderthalensis. Vedou se spory, zda tvoří „neandrtálec“ samostatný druh, nebo jestli se jedná o poddruh Homo sapiens. Genetické analýzy prokázaly, že docházelo ke křížení mezi zástupci moderního člověka a neandertálci. V genomu jedinců neafrických populací člověka jsou obsažena cca 2 procenta neandertálských genů, v celkovém genofondu lidstva bylo zjištěno zatím asi 20 procent neandertálského genomu přičemž další výzkum naznačuje, že by ho mohlo být až 40%. To však nemusí znamenat žádné rozsáhlejší křížení, dle vyjádření vědců stačí k takovému zastoupení genů pouhých 300 jednotlivých aktů vedoucích k životaschopnému potomstvu.[75] Nicméně, dle nových výzkumů je jisté, že neandrtálci a člověk moderního typu spolu koexistovali více než 5000 let a pravděpodobně u těchto dvou druhů docházelo ke kontaktům jak kulturním a společenským tak i fyzickým (tedy mísení druhů).[76]
Denisované
Z molekulárních analýz vyplývá, že se před asi 500 000 lety od vývojové linie vedoucí k modernímu člověku nejprve oddělila větev, zahrnující denisovany a neandertálce, ze které se později denisované oddělili, ale přesto docházelo ke křížení denisovanů a neandrtálců ještě v době před 80 000 lety.[77] Ke křížení s moderními lidmi docházelo ještě před 15 000 lety.[78]
Neznámý druh
Podle analýz DNA člověka existoval minimálně další předek,[79] se kterým se moderní člověk křížil, ale pro který nebyly nalezeny pozůstatky. Oddělil se od linie člověka před více než půl milionem let.[80] Pocházel z Asie a Oceánie.[81] Takových linií identifikovaných pouze genetickou analýzou současné populace mohlo být více.[82]
Homo floresiensis
Obýval ostrov Flores v dnešní Indonésii před 95 000–12 000 lety a byl v průměru asi 110 cm vysoký. Jeho evoluce je nejistá, předpokládá se původ z lokální populace Homo erectus nebo ještě dřívějších hominidů (Homo habilis či georgicus)
Homo sapiens
Jeden typ tohoto člověka (se širokým obličejem) se nazývá kromaňonec podle naleziště Cro-Magnon ve Francii. Na území České republiky se nachází řada nalezišť: Předmostí v Přerově, Dolní Věstonice, Pavlov, Brno, Koněprusy, Mladeč aj.
Druh Homo sapiens žije od období před 800 000 lety dodnes. Mezi obdobím před 400 000 lety a druhou meziledovou dobou, asi před 250 000 lety, se zvětšovala jeho lebka a energetická náročnost mozku.[83] Vyvinulo se lepší zpracovávání kamenných nástrojů, což dokládá změnu z Homo erectus na Homo sapiens. Důkazy dokládají migraci Homo erectus z Afriky a následnou transformaci Homo erectus na Homo sapiens v Africe (jinde důkazy chybějí). Následující migrace (expanze, kterou záhájilo klimatické zlepšení následující Eemský interglaciál) v rámci Afriky a ven z ní nahradila dříve rozšířený druh Homo erectus. Ovšem Homo sapiens byl v Evropě již před více než 200 tisíci lety. O tom svědčí nález v jeskyni Apidima (na Peloponésu).[84] V současnosti nejsou žádné další důkazy o různém vývoji Homo sapiens na různých místech světa, a tak je tato oblast častým debatním tématem v oblasti paleoantropologie. V Asii dochází ke křížení s původními formami člověka.[85] Do Evropy se Homo sapiens rozšířil v několika vlnách, a to nejen z jihovýchodu, ale také z jihozápadu (přes dnešní Španělsko).[86] Charakteristickým znakem člověka je domestikace, kterou pravděpodobně člověk působil sám na sebe,[87] a jejíž rysy ale nejsou patrné na neandrtálcích a denisovanech.[88] Všichni současní lidé mají jen 1,5 % společné unikátní DNA a v současném člověku z celého světa lze najít 7 % DNA, která nepatřila neandrtálcům či denisovanům.[89] Adaptaci na změny prostředí jako jednoho z hybatelů evoluce člověka nyní řídí spíše kulturní vývoj, než genetické změny.[90]
Do Jižní Ameriky se člověk moderního typu dostal patrně již před asi 30 000 lety, jak ukazují nové objevy z Argentiny.[91]
Odkazy
Reference
- DART, R. A.; CRAIG, D. Dobrodružství s chybějícím článkem. 1. vyd. Praha: Mladá fronta, 1963. 300 s. 23-027-63.
- WOOD, Bernard; RICHMOND, Brian G. Human evolution: taxonomy and paleobiology. S. 19–60. Journal of Anatomy [online]. 2000-07. Roč. 197, čís. 1, s. 19–60. Dostupné online. DOI 10.1046/j.1469-7580.2000.19710019.x. PMID 10999270. (anglicky)
- BEGUN, D. R. Fossil record of Miocene hominoids. In: HENKE, W. C.; TATTERSALL, I. Handbook of palaeoanthropology 2. Berlín: Springer, 2007. S. 921–977.
- HARRISON, T. Catarrhine Origins. In: BEGUN, D. R. A Companion to Paleoanthropology. Oxford: Willey - Blackwell, 2013. S. 376–396.
- RASMUSSEN, D. T. Early Catarrhines of the African Eocene and Oligocene. In: HARTWIG, W. C. The Primate Fossil Record. Cambridge: Cambridge University Press, 2002. S. 203–220.
- ZALMOUT, I. S., et al. New Oligocene primate from Saudi Arabia and the divergence of apes and Old World monkeys. Nature. Červenec 2010, roč. 466, čís. 7304, s. 360–364. Dostupné online.(anglicky)
- LEAKEY, M.G.; UNGAR, P.S.; WALKER, A. A new genus of large primate from the Late Oligocene of Lothidok, Turkana District, Kenya. S. 519–531. Journal of Human Evolution [online]. 1995-06. Roč. 28, čís. 6, s. 519–531. Dostupné online. DOI 10.1006/jhev.1995.1040. (anglicky)
- CARTMILL, M.; SMITH, F. H. The Human Lineage. Oxford: Willey-Blackwell, 2011.
- MAZÁK, V. Sága rodu Homo. Praha: Práce, 1986. 424 s.
- HARRISON, T. Dendropithecoidea, Proconsuloidea and Hominoidea. In: WERDELIN, L.; SANDERS, W. J. Cenozoic Mammals of Africa. Berkeley: University of California Press, 2010. S. 429–469.
- BEGUN, D. R. Planet of the Apes. Scientific American. 2003, roč. 289, s. 74–83. Dostupné online.
- DAWKINS, R. Příběh předka: pouť k úsvitu života. Praha: Academia, 2008. 831 s.
- CONROY, G. C. Reconstructing human origins. New York: W. W. Norton & company, 1997. 547 s. Dostupné online.
- KOUFOS, G. D. Potential hominoid ancestors for Hominidae. In: HENKE, W. C.; TATTERSALL, I. Handbook of palaeoanthropology 3. Berlín: Springer, 2007. S. 1347–1377.
- BROOKS, A. S., et al. Encyclopedia of human evolution and prehistory. New York: Taylor & Francis, 2000. 800 s.
- KELLEY, J. The hominoid radiation in Asia. In: HARTWIG, W. C. The primate fossil record. Cambridge: Cambridge University Press, 2002. Kapitola 21, s. 369–384.
- BEGUN, David R. Dryopithecins, Darwin, de Bonis, and the European origin of the African apes and human clade. S. 789–816. Geodiversitas [online]. 2009-12. Roč. 31, čís. 4, s. 789–816. Dostupné online. DOI 10.5252/g2009n4a789. (anglicky)
- MOYÀ-SOLÀ, Salvador; KÖHLER, Meike; ALBA, David M.; CASANOVAS-VILAR, Isaac; GALINDO, Jordi. Pierolapithecus catalaunicus , a New Middle Miocene Great Ape from Spain. S. 1339–1344. Science [online]. 2004-11-19. Roč. 306, čís. 5700, s. 1339–1344. Dostupné online. DOI 10.1126/science.1103094. (anglicky)
- KUNIMATSU, Yutaka; NAKATSUKASA, Masato; SAWADA, Yoshihiro; SAKAI, Tetsuya; HYODO, Masayuki; HYODO, Hironobu; ITAYA, Tetsumaru. A new Late Miocene great ape from Kenya and its implications for the origins of African great apes and humans. S. 19220–19225. Proceedings of the National Academy of Sciences [online]. 2007-12-04. Roč. 104, čís. 49, s. 19220–19225. Dostupné online. DOI 10.1073/pnas.0706190104. PMID 18024593. (anglicky)
- BEGUN, David R. The Earliest Hominins--Is Less More?. S. 1478–1480. Science [online]. 2004-03-05. Roč. 303, čís. 5663, s. 1478–1480. Dostupné online. DOI 10.1126/science.1095516. PMID 15001766. (anglicky)
- WHITE, T. D. Early hominid femora: The inside story. Comptes Rendus Palevol. 2006, roč. 5, s. 99–108.
- BEGUN, David R. Miocene Hominids and the Origins of the African Apes and Humans. S. 67–84. Annual Review of Anthropology [online]. 2010-10-21. Roč. 39, čís. 1, s. 67–84. Dostupné online. DOI 10.1146/annurev.anthro.012809.105047. (anglicky)
- Oldest footprints of pre-humans identified in Crete. phys.org [online]. 2021-10-11 [cit. 2022-05-02]. Dostupné online. (anglicky)
- MACCHIARELLI, Roberto; BERGERET-MEDINA, Aude; MARCHI, Damiano; WOOD, Bernard. Nature and relationships of Sahelanthropus tchadensis. S. 102898. Journal of Human Evolution [online]. 2020-12. Roč. 149, s. 102898. Dostupné online. DOI 10.1016/j.jhevol.2020.102898. (anglicky)
- YIRKA, Bob. Study of partial left femur suggests Sahelanthropus tchadensis was not a hominin after all. phys.org [online]. 2020-11-24 [cit. 2022-05-02]. Dostupné online. (anglicky)
- SIMPSON, S. W. Before Australopithecus: The Earliest hominins. In: BEGUN, D. R. A Companion to Paleoanthropology. Oxford: Willey-Blackwell, 2012. S. 417–433.
- GIBBONS, A. První lidé: závody v hledání nejstaršího předka. Praha: Academia, 2011. 307 s.
- MCBREARTY, Sally; JABLONSKI, Nina G. First fossil chimpanzee. S. 105–108. Nature [online]. 2005-09. Roč. 437, čís. 7055, s. 105–108. Dostupné online. DOI 10.1038/nature04008. (anglicky)
- ACKERMANN, Rebecca Rogers; SMITH, Richard J. The Macroevolution of our Ancient Lineage: What We Know (or Think We Know) about Early Hominin Diversity. S. 72–85. Evolutionary Biology [online]. 2007-08-03. Roč. 34, čís. 1-2, s. 72–85. Dostupné online. DOI 10.1007/s11692-007-9002-7. (anglicky)
- HAILE-SELASSIE, Yohannes; SUWA, Gen; WHITE, Tim D. Late Miocene Teeth from Middle Awash, Ethiopia, and Early Hominid Dental Evolution. S. 1503–1505. Science [online]. 2004-03-05. Roč. 303, čís. 5663, s. 1503–1505. Dostupné online. DOI 10.1126/science.1092978. PMID 15001775. (anglicky)
- ANDREWS, P.; HARRISON, T. The last common ancestor of apes and humans. In: LIEBERMAN, D. E.; SMITH, R. J.; KELLEY, J. Interpreting the Past: Essays on Human, Primate, and Mammal Evolution. Lieden, Boston: Brill Academic Publishing, 2005. S. 103–121.
- COPPENS, Y. The Bunch of Prehumans and the Emergence of the Genus Homo. The Proceedings of the Plenary Session on Scientific Insights into the Evolution of the Universe and of Life. 2009, roč. 2008, s. 367–372.
- ROCHE, Damien; SÉGALEN, Loïc; SENUT, Brigitte; PICKFORD, Martin. Stable isotope analyses of tooth enamel carbonate of large herbivores from the Tugen Hills deposits: Palaeoenvironmental context of the earliest Kenyan hominids. S. 39–51. Earth and Planetary Science Letters [online]. 2013-11. Roč. 381, s. 39–51. Dostupné online. DOI 10.1016/j.epsl.2013.08.021. (anglicky)
- BALTER, Michael; GIBBONS, Ann. Another Emissary From the Dawn of Humanity. S. 187–189. Science [online]. 2001-07-13. Roč. 293, čís. 5528, s. 187–189. Dostupné online. DOI 10.1126/science.293.5528.187b. (anglicky)
- HARCOURT-SMITH, William H. E. The First Hominins and the Origins of Bipedalism. S. 333–340. Evolution: Education and Outreach [online]. 2010-09. Roč. 3, čís. 3, s. 333–340. Dostupné online. DOI 10.1007/s12052-010-0257-6. (anglicky)
- BRUNET, Michel; GUY, Franck; PILBEAM, David; MACKAYE, Hassane Taisso; LIKIUS, Andossa; AHOUNTA, Djimdoumalbaye; BEAUVILAIN, Alain. A new hominid from the Upper Miocene of Chad, Central Africa. S. 145–151. Nature [online]. 2002-07-11. Roč. 418, čís. 6894, s. 145–151. Dostupné online. DOI 10.1038/nature00879. PMID 12110880. (anglicky)
- SENUT, Brigitte; PICKFORD, Martin; GOMMERY, Dominique; MEIN, Pierre; CHEBOI, Kiptalam; COPPENS, Yves. First hominid from the Miocene (Lukeino Formation, Kenya). S. 137–144. Comptes Rendus de l'Académie des Sciences - Series IIA - Earth and Planetary Science [online]. 2001-01. Roč. 332, čís. 2, s. 137–144. Dostupné online. DOI 10.1016/S1251-8050(01)01529-4. (anglicky)
- HAILE-SELASSIE, Y.; WOLDEGABRIEL, G. Ardipithecus kadabba: late Miocene evidence from the Middle Awash, Ethiopia. Berkeley: University of California Press, 2009. 641 s.
- GIBBONS, Ann. Ardipithecus ramidus. S. 1598–1599. Science [online]. 2009-12-18. Roč. 326, čís. 5960, s. 1598–1599. Dostupné online. DOI 10.1126/science.326.5960.1598-a. PMID 20019252. (anglicky)
- REED, K. E.; FISH, J. L. Tropical and temperate seasonal influences on human evolution. In: BROCKMAN, D. K.; VAN SCHAIK, C. P. Seasonality in Primates: Studies of Living and Extinct Human and Non-Human Primates. Cambridge: Cambridge University Press, 2005. S. 491–520.
- STRAIT, David S. The Evolutionary History of the Australopiths. S. 341–352. Evolution: Education and Outreach [online]. 2010-09. Roč. 3, čís. 3, s. 341–352. Dostupné online. DOI 10.1007/s12052-010-0249-6. (anglicky)
- LEWIN, R. Human evolution: an illustrated introduction. 5. vyd. Suffolk: Wiley-Blackwell, 2004. 284 s. Dostupné online. ISBN 978-1405103787.
- AIELLO, Leslie C. Variable but singular. S. 399–400. Nature [online]. 1994-03. Roč. 368, čís. 6470, s. 399–400. Dostupné online. DOI 10.1038/368399a0. (anglicky)
- CARTMILL, M.; SMITH, M. H. The Human Lineage. New Jersey: Willey-Blackwell, 2009. 624 s. Dostupné online.
- RICHMOND, Brian G.; AIELLO, Leslie C.; WOOD, Bernard A. Early hominin limb proportions. S. 529–548. Journal of Human Evolution [online]. 2002-10-01. Roč. 43, čís. 4, s. 529–548. Dostupné online. DOI 10.1006/jhev.2002.0594. PMID 12393007. (anglicky)
- SUSSMAN, R. W.; HART, D. Modeling the Past: The Primatological Approach. In: HENKE, W. C.; TATTERSALL, I. Handbook of palaeoanthropology 2. Berlín: Springer, 2007. S. 702–721.
- RAK, Yoel. Lucy's pelvic anatomy: its role in bipedal gait. S. 283–290. Journal of Human Evolution [online]. 1991-04. Roč. 20, čís. 4, s. 283–290. Dostupné online. DOI 10.1016/0047-2484(91)90011-J. (anglicky)
- COLLARD, M.; WOOD, B. Grades among the African Early Hominids. In: BROMAGE, T. G.; SCHRENK, F. African Biogeography. Climate Change & Human Evolution. Oxford: Oxford University Press, 1999. S. 316–327.
- ŠMAHEL, Zbyněk. Příběh lidského rodu. Brno: Moravské zemské muzeum, 2005. 85 s. ISBN 80-7028-262-2.
- BEHRENSMEYER, Anna K.; REED, Kaye E. The paleobiology of Australopithecus : contributions from the fourth Stony Brook Human Evolution Symposium and Workshop, Diversity in Australopithecus: Tracking the First Bipeds, September 25-28, 2007. In: REED, Kaye E.; FLEAGLE, John G.; LEAKEY, Richard. The Paleobiology of Australopithecus. Dordrecht: Springer, 2013. Dostupné online. ISBN 978-94-007-5919-0. DOI 10.1007/978-94-007-5919-0_4. S. 41–60. (anglicky)
- CODRON, Daryl; LEE-THORP, Julia A.; SPONHEIMER, Matt; DE RUITER, Darryl; CODRON, Jacqui. What Insights Can Baboon Feeding Ecology Provide for Early Hominin Niche Differentiation?. S. 757–772. International Journal of Primatology [online]. 2008-06. Roč. 29, čís. 3, s. 757–772. Dostupné online. DOI 10.1007/s10764-008-9261-x. (anglicky)
- SPONHEIMER, Matt; DUFOUR, Darna L. The evolution of hominin diets : integrating approaches to the study of palaeolithic subsistence. In: HUBLIN, J.J.; RICHARDS, M.P. The Evolution of Hominin Diets. Dordrecht: Springer, 2009. Dostupné online. ISBN 978-1-4020-9698-3. DOI 10.1007/978-1-4020-9699-0_18. S. 229–240. (anglicky)
- BERTHAUME, Michael; GROSSE, Ian R.; PATEL, Nirdesh D.; STRAIT, David S.; WOOD, Sarah; RICHMOND, Brian G. The Effect of Early Hominin Occlusal Morphology on the Fracturing of Hard Food Items. S. 594–606. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology [online]. 2010-04. Roč. 293, čís. 4, s. 594–606. Dostupné online. DOI 10.1002/ar.21130. PMID 20235316. (anglicky)
- SPONHEIMER, Matt; LEE-THORP, Julia A. Differential resource utilization by extant great apes and australopithecines: towards solving the C4 conundrum. S. 27–34. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology [online]. 2003-09. Roč. 136, čís. 1, s. 27–34. Dostupné online. DOI 10.1016/S1095-6433(03)00065-5. PMID 14527627. (anglicky)
- Brain size of human ancestors evolved gradually over 3 million years. phys.org [online]. 2018-02-21 [cit. 2022-05-02]. Dostupné online. (anglicky)
- A taste for fat may have made us human, says study. phys.org [online]. 2019-02-05 [cit. 2022-05-02]. Dostupné online. (anglicky)
- First human ancestors breastfed for longer than contemporary relatives. phys.org [online]. 2019-08-29 [cit. 2022-05-02]. Dostupné online. (anglicky)
- HOUSER, Pavel. Člověk byl přes 2 miliony let vrcholovým predátorem. sciencemag.cz [online]. 2021-04-07 [cit. 2022-05-02]. Dostupné online.
- When and why did human brains decrease in size 3,000 years ago? Ants may hold clues. phys.org [online]. 2021-08-22 [cit. 2022-05-02]. Dostupné online. (anglicky)
- Processing food before eating likely played key role in human evolution, study finds. phys.org [online]. 2016-03-09 [cit. 2022-05-02]. Dostupné online. (anglicky)
- DIMAGGIO, Erin N.; CAMPISANO, Christopher J.; ROWAN, John; DUPONT-NIVET, Guillaume; DEINO, Alan L.; BIBI, Faysal; LEWIS, Margaret E. Late Pliocene fossiliferous sedimentary record and the environmental context of early Homo from Afar, Ethiopia. S. 1355–1359. Science [online]. 2015-03-20. Roč. 347, čís. 6228, s. 1355–1359. Dostupné online. DOI 10.1126/science.aaa1415. (anglicky)
- Researcher's paper challenges the claim that the genus Homo originated in response to environmental changes. phys.org [online]. 2017-08-04 [cit. 2022-05-02]. Dostupné online. (anglicky)
- Climatic variability might not drive evolutionary change as much as previously thought, study finds. phys.org [online]. 2022-04-11 [cit. 2022-05-02]. Dostupné online. (anglicky)
- (anglicky) Elaine Morgan: I believe we evolved from aquatic apes (Elaine Morganová říká, že jsme se vyvinuli z vodních opic) Archivováno 15. 6. 2010 na Wayback Machine, ted.com, Ted talks, TEDGlobal, v červenci 2009, video s českými titulky.
- Petrželka, Alexandr: Proč nejsme chlupatí jako opice, novinky.cz, 1.11.2012, původně pro Právo.
- New hypothesis explains how human ancestors used fire to their advantage. phys.org [online]. 2016-04-12 [cit. 2022-05-02]. Dostupné online. (anglicky)
- Did human-like intelligence evolve to care for helpless babies?. medicalxpress.com [online]. 2016-05-23 [cit. 2022-05-02]. Dostupné online. (anglicky)
- Researchers wonder if ancient supernovae prompted human ancestors to walk upright. phys.org [online]. 2019-05-28 [cit. 2022-05-02]. Dostupné online. (anglicky)
- MIKSCH, Joe. Researcher argues that there's more to the genus Homo than we may think. phys.org [online]. 2015-08-28 [cit. 2022-05-02]. Dostupné online. (anglicky)
- New species of early human found in the Philippines. phys.org [online]. 2019-04-10 [cit. 2022-05-02]. Dostupné online. (anglicky)
- REICH, David; GREEN, Richard E.; KIRCHER, Martin; KRAUSE, Johannes; PATTERSON, Nick; DURAND, Eric Y.; VIOLA, Bence. Genetic history of an archaic hominin group from Denisova Cave in Siberia. S. 1053–1060. Nature [online]. 2010-12. Roč. 468, čís. 7327, s. 1053–1060. Dostupné online. ISSN 1476-4687. DOI 10.1038/nature09710. (anglicky)
- ZIMMER, Carl. Siberian Fossils Were Neanderthals’ Eastern Cousins, DNA Reveals. The New York Times [online]. 2010-12-22 [cit. 2022-05-03]. Popularizační článek k předchozí referenci. Dostupné online. ISSN 0362-4331. (anglicky)
- ANTÓN, Susan C.; POTTS, Richard; AIELLO, Leslie C. Evolution of early Homo: An integrated biological perspective. S. 1236828. Science [online]. 2014-07-04. Roč. 345, čís. 6192, s. 1236828. Dostupné online. DOI 10.1126/science.1236828. (anglicky)
- Discovery of ancient tools in China suggests humans left Africa earlier than previously thought. phys.org [online]. 2018-07-11 [cit. 2022-05-02]. Dostupné online. (anglicky)
- At least 20% of Neanderthal DNA Is in Humans. Live Science. Dostupné online [cit. 2018-06-06].
- Neanderthals Coexisted with Humans for More Than 5,000 Years | Anthropology | Sci-News.com. Breaking Science News | Sci-News.com. Dostupné online [cit. 2018-06-07]. (anglicky)
- Z molekulárních analýz vyplývá, že se před asi 500 000 lety od vývojové linie vedoucí k modernímu člověku nejprve oddělila větev, zahrnující denisovany a neandertálce, ze které se později denisované oddělili, ale přesto docházelo ke křížení denisovanů a neandrtálců ještě v době před 80 000 lety.
- YIRKA, Bob. Evidence found of Denisovans interbreeding with humans in Southeast Asia more recently than thought. phys.org [online]. 2019-04-02 [cit. 2022-05-02]. Dostupné online. (anglicky)
- MIHULKA, Stanislav. Umělá inteligence vystopovala přízračného předka v našem genomu. osel.cz [online]. 2019-02-11 [cit. 2022-05-02]. Dostupné online.
- CHOI, Charles Q. Human Ancestor Mated with 'Ghost Lineage' And the Proof Is in Your Spit. livescience.com [online]. 2017-07-25 [cit. 2022-05-02]. Dostupné online. (anglicky)
- MONDAL, Mayukh; BERTRANPETIT, Jaume; LAO, Oscar. Approximate Bayesian computation with deep learning supports a third archaic introgression in Asia and Oceania. S. 246. Nature Communications [online]. 2019-12. Roč. 10, čís. 1, s. 246. Dostupné online. DOI 10.1038/s41467-018-08089-7. (anglicky)
- BRAHIC, Catherine. Traces of mystery ancient humans found lurking in our genomes. newscientist.com [online]. 2018-10-10 [cit. 2022-05-02]. Dostupné online. (anglicky)
- Smarter brains are blood-thirsty brains. phys.org [online]. 2016-08-30 [cit. 2022-05-02]. Dostupné online. (anglicky)
- DELSON, Eric. An early dispersal of modern humans from Africa to Greece. S. 487–488. Nature [online]. 2019-07. Roč. 571, čís. 7766, s. 487–488. Dostupné online. DOI 10.1038/d41586-019-02075-9. (anglicky)
- Ancient molar points to interbreeding between archaic humans and Homo sapiens in Asia. phys.org [online]. 2019-07-08 [cit. 2022-05-02]. Dostupné online. (anglicky)
- Genetic analysis of Ice Age Europeans. phys.org [online]. 2016-05-02 [cit. 2022-05-02]. Dostupné online. (anglicky)
- KOTTKE, Jason. The Self-Domestication of Humans. kottke.org [online]. 2019-01-11 [cit. 2022-05-02]. Dostupné online. (anglicky)
- THEOFANOPOULOU, Constantina; GASTALDON, Simone; O’ROURKE, Thomas; SAMUELS, Bridget D.; MESSNER, Angela; MARTINS, Pedro Tiago; DELOGU, Francesco. Self-domestication in Homo sapiens: Insights from comparative genomics. S. e0185306. PLOS ONE [online]. 2017-10-18. Roč. 12, čís. 10, s. e0185306. Dostupné online. DOI 10.1371/journal.pone.0185306. PMID 29045412. (anglicky)
- LARSON, Christina. Just 7% of our DNA is unique to modern humans, study shows. phys.org [online]. 2021-07-16 [cit. 2022-05-02]. Dostupné online. (anglicky)
- Researchers: Culture drives human evolution more than genetics. phys.org [online]. 2021-06-03 [cit. 2022-05-02]. Dostupné online. (anglicky)
- AZCUY, Carlos L.; CARRIZO, Hugo A.; TONNI, Eduardo P.; PANARELLO, Héctor; AMENÁBAR, Cecilia R. Late Pleistocene fossil hominids tracks on the beaches of Claromecó, Argentina. S. 43–57. Acta Geológica Lilloana [online]. 2021-08-26. S. 43–57. Dostupné online. DOI 10.30550/j.agl/2021.33.2/2021-08-03. (anglicky)
Literatura
- DAWKINS, Richard. Příběh předka: pouť k úsvitu života. Praha: Academia, 2008. 831 s. Galileo, sv. 18. ISBN 978-80-200-1611-9.
- DUNBAR, R. I. M. Příběh rodu Homo: nové dějiny evoluce člověka. Praha: Academia, 2009. 222 s. Galileo, sv. 33. ISBN 978-80-200-1715-4.
- FLEGR, Jaroslav. Zamrzlá evoluce, aneb, Je to jinak, pane Darwin. Praha: Academia, 2006. 326 s. Galileo, sv. 4. ISBN 80-200-1453-5.
- GIBBONS, Ann. První lidé: závody v hledání nejstaršího předka. Praha: Academia, 2011. 307 s. ISBN 978-80-200-1978-3. Galileo, sv. 48.
- RICHERSON, Peter J. a BOYD, Robert. V genech není všechno, aneb, Jak kultura změnila evoluci člověka. Praha: Academia, 2012. 407 s. Galileo, sv. 51. ISBN 978-80-200-2066-6.
- ROBERTS, Alice M. Evoluce: příběh člověka. Praha: Knižní klub, 2012. 256 s. ISBN 978-80-242-3597-4. Universum.
- ROČEK, Zbyněk. Historie obratlovců: evoluce, fylogeneze, systém. Praha: Academia, 2002. 512 s. ISBN 80-200-0858-6.
- SVOBODA, Jiří A. Předkové: evoluce člověka. 2., uprav. vyd. Úvodní texty Miloš Macholán. Praha: Academia, 2017. 479 s. ISBN 978-80-200-2750-4.
- SVOBODA, Jiří A. a MALINA, Jaroslav, ed. Panoráma antropologie biologické – sociální – kulturní: modulové učební texty pro studenty antropologie a „příbuzných“ oborů. 32, Čas lovců: aktualizované dějiny paleolitu. Brno: Nadace Universitas, 2009. 298 s. Scientia. ISBN 978-80-7204-628-7.
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu vývoj člověka na Wikimedia Commons
Obrázky, zvuky či videa k tématu vývoj člověka na Wikimedia Commons - Taxonomie člověka na serveru BioLib
- Projekt Human origins (National Geographic) (anglicky)
- Lidská evoluce na stránkách Smithsonian institution (anglicky)
- Indonéská senzace – hobit! 31. 12. 2004 Svět vědy
- Mapping Transdisciplinarity in Human Sciences
- Fundamental Theory of Human Sciences