Australopithecus africanus
Australopithecus africanus (česky též Australopiték africký)[1] je druh vyhynulého hominida, žijící na přelomu pliocénu a pleistocénu, před 3 - 2 miliony let v jižní Africe, na území dnešní Jihoafrické republiky. Ačkoliv není geologicky nejstarším australopitékem, byl rozeznán jako první a je tudíž typovým druhem celého rodu. Zároveň se jedná o jednoho z nejznámějších a nejlépe poznaných zástupců svého rodu. Je také jedním z kandidátů na předka rodu Homo.[1]
Stratigrafický výskyt: pliocén - pleistocén | |
|---|---|
 | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Třída | savci (Mammalia) |
| Řád | primáti (Primates) |
| Čeleď | Hominidé (Hominidae) |
| Podčeleď | Homininae |
| Tribus | Hominini |
| Rod | Australopithecus Dart, 1925 |
| Binomické jméno | |
| Australopithecus africanus Dart, 1925 | |
| Některá data mohou pocházet z datové položky. | |
Objevy
První ostatky popsal Raymond A. Dart. V nálezech zvířecích kostí z vápencových lomů poblíž města Taung (původně Taungs) v tehdejším Bečuánsku[2] objevil 28. listopadu 1924 taungské dítě - obličejovou část lebky, do níž přesně zapadal detailně dochovaný přirozený výlitek mozkovny.[3]
Dart si povšiml, že lebka nese znaky lidoopů i člověka a pochopil, že zachytil přelomový nález - pojící článek mezi šimpanzi a lidmi, potvrzující domněnku Charlese Darwina o původu lidstva v Africe. Dart své závěry publikoval 7. února 1925.[4] Pro lebku navrhl odborný název Australopithecus africanus (v překladu „jižní opice z Afriky“), a to podle málo očekávaného místa nálezu v Africe, na jižní polokouli.[3]
Zpráva však strhla velkou vlnu kritiky. Odborná veřejnost nález nepřijala a taungské dítě pokládala za pozůstatek vyhynulé opice nebo nanejvýš lidoopa, nikoliv lidského předka. Na počátku 20. století totiž byly doklady pliocénních a pleistocénních hominidů prakticky neznámé. Mnoho badatelů hledalo kolébku lidstva v Asii, kde byly právě odhaleny ostatky starších forem rodu Homo. Na základě podvrženého Piltdownského člověka - upravené lebky středověkého člověka spojené se spodní čelistí orangutana - se navíc pátralo po předcích s velkým mozkem a opičími zuby. Taungské dítě ale bylo přesně opačné - s téměř lidskými zuby a malým mozkem.[5] Pocházelo ze „zaostalé“ Afriky a lebka mláděte i přes dobrou zachovalost nemohla vypovídat o vzhledu dospělých jedinců. Většina „lidských“ znaků tudíž byla přičítána právě nedospělosti jedince.[3]

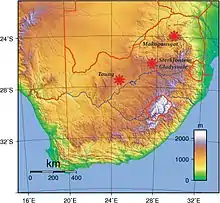
Takřka jediným, kdo názory R. A. Darta v té době přijal a podporoval, byl původem skotský lékař Robert Broom, který se po získání práce v Transvaalském muzeu v Pretorii roku 1934 sám pustil do pátrání po dalších ostatcích.[6] Dne 17. srpna 1936 získal z vápencového lomu u Sterkfontein výlitek lebky dospělého australopitéka a postupně se mu podařilo odhalit i další ostatky.[7] Nálezy byly ovšem popsány jako samostatný druh Australopithecus transvaalensis[7] a v roce 1938 dokonce kvůli drobným odlišnostem v utváření zubů a čelisti přesunuty do samostatného rodu Plesianthropus (P. transvaalensis).[8]
Během druhé světové války byly odkryvy v jižní Africe přerušeny, zejména kvůli zastavení těžby vápence.[9] Pokračovaly však práce na vydání monografie o jihoafrických australopitécích, která nakonec vyšla v roce 1946.[10] Význam australopitéků v lidské evoluci již nebylo možné přehlédnout. Vlna ostrého odporu opadla do konce třicátých let 20. století a koncem let čtyřicátých již badatelé rod Australopithecus přijali jako výchozí bod lidské vývojové linie.[11]
V květnu roku 1947 byly práce ve Sterkfontein obnoveny a téměř okamžitě byly zachyceny další početné ostatky.[12] 18. dubna 1947 zde Robert Broom zachytil i velmi dobře dochovanou lebku STS 5, která podle tehdejšího rodového názvu získala přezdívku Mrs. Ples.[3] Téhož roku popsal R. A. Dart nálezy z nové lokality Makapansgat a vytvořil pro ně samostatný druh Australopithecus prometheus.[13]
V roce 1954 navrhl John T. Robinson shrnout všechny známé druhy - Australopithecus africanus, Australopithecus prometheus a Plesianthropus transvaalensis - do jediného druhu,[14] což bylo ostatními badateli téměř bez výhrad přijato. Podle pravidel biologické nomenklatury dostalo přednost nejstarší pojmenování Australopithecus africanus.[6]
Přestože v 50. až 70. letech 20. století byly nálezy z jižní Afriky částečně zastíněny východoafrickými objevy nových druhů australopitéků i prvních lidí (Olduvai, Hadar, Laetoli), výzkumy na jihu kontinentu pokračovaly dál a přinášely stovky nálezů homininů, především ve Sterkfontein. Kromě částí lebek a čelistí byly získány i početné zlomky kostí končetin a trupu.[15]

V roce 1991 byly objeveny dva zlomky zubů druhu A. africanus také v Gladysvale. Poprvé od roku 1948 tak bylo zjištěno nové naleziště tohoto druhu australopitéků.[16]
V 90. letech 20. století se práce ve Sterkfontein přesunuly i do níže položených dutin. V roce 1995 zde byly odhaleny zlomky kostí australopitéků v Jacovec cavern[17] a roku 1997 i v Silberberg Grotto.[18] Kvůli mírným odlišnostem od ostatních nálezů však zatím není jasné, jestli patří k zástupcům Australopithecus africanus nebo zcela novému druhu australopitéků.[17]
Datace
Jihoafrické krasové jeskyně jsou oproti východoafrickým lokalitám jen obtížně datovatelné. V jejich výplni chybí vrstvy sopečného popela, na něž lze dobře aplikovat radiometrické datovací metody. Uplatňují se tak především přístupy relativní chronologie. Tradiční je využití biostratigrafie, tedy datace na základě přítomnosti určitých živočišných druhů. Jejich výskyt je přitom srovnáván s dobře datovatelnými lokalitami východní Afriky.[19] Teprve v posledních letech technický rozvoj umožnil uplatnění absolutních datovacích metod i v jižní Africe - využíván je paleomagnetismus,[20] optická luminiscence[21] nebo metoda uran-olovo.[22]
Ani výsledky těchto metod ovšem nejsou jednoznačně přijímány, různé přístupy často poskytují výrazně odlišná data. Problémem je mimo jiné velmi složitá a nesouvislá stratigrafie jeskynních výplní, především ve Sterkfontein.[20] Rovněž datace nálezů z Taung je problematická, neboť došlo k úplnému zničení původní lokality postupující těžbou vápence.[23] Slibnějším nalezištěm je Gladysvale, kde se částečně zachovala nezvykle dobře čitelná stratigrafie.[21]
Nejčastěji se tedy výskyt druhu Australopithecus africanus uvádí v době před 3 – 2 miliony let,[23] přičemž nejstarší nálezy ze Sterkfontein mohou dosahovat stáří až 3,67 milionu let.[24][25]
Popis

Australopithecus africanus patří mezi nejlépe poznané australopitéky, přestože je mnoho kostí dochováno jen částečně, ve zlomcích. Drtivá většina nálezů pochází ze Sterkfontein. Stejně jako u ostatních australopitéků, se i u druhu A. africanus předpokládá nápadný pohlavní dimorfismus ve velikosti těla. Samci tak mohli v průměru dosahovat výšky asi 138 cm při váze 41 kg, zatímco samice byly vysoké jen asi 115 cm, s váhou zhruba 30 kg.[26]
Díky nálezům přirozených výlitků mozkoven je intezivně studovaná nejen velikost, ale také stavba mozku. Jednotlivé rysy jsou však dosud předmětem živých debat, neboť jejich charakter a výskyt je na dochovaných ostatcích nejednoznačný nebo proměnlivý.[27] Většina odborníků se shoduje, že lze pozorovat náznaky redukce týlních partií mozku (především primární zrakové korové oblasti) a naopak rozšiřování temenního a čelního laloku i přední části spánkového laloku. Měsíčitá rýha (sulcus lunatus) se proto posouvá směrem vzad.[27] Celkově jsou však tvar i velikost mozku stále ještě bližší moderním lidoopům než lidem.[28]
Hojně diskutovaná je rovněž stavba páteře. Na základě dvou nálezů částečných koster ze Sterkfontein se často předpokládá, že A. africanus (a snad i ostatní australopitékové) měl delší bederní páteř, tvořenou 6 obratli. Taková páteř mohla být dědictvím po starších hominidech, přičemž u lidí se tento počet později snížil na 5 a u moderních lidoopů dokonce na 3-4 obratle.[29] Delší, tedy i pohyblivější bederní páteř mohla australopitékům usnadňovat vzpřímený postoj i chůzi, snižovat tlak na meziobratlové ploténky a lépe bránit poškození. K redukci počtu obratlů by pak došlo při zlepšení bipedie u mladších zástupců rodu Homo (ještě Homo erectus měl ovšem podle nálezu u jezera Turkana šest bederních obratů).[30] Odborníci však v této otázce nejsou jednotní a někteří z nich předpokládají, že první bederní obratel může být spíše špatně určený poslední obratel hrudní, takže australopitéci by stejně jako moderní lidé měli jen pět bederních obratlů.[31] Přítomnost šesti bederních obratlů tak dosud nebyla jednoznačně prokázána ani vyvrácena. Doklady obratlů a žeber jsou navíc vzácné, a tak nelze vyloučit, že se v případě nálezů ze Sterkfontein jedná jen o výjimku, anomálii.[32]

Lebka

Znaky mozkovny:[3]
- protáhlý, úzký tvar podobně jako u šimpanzů
- malý objem - velikost mozku se pohybuje při horní hranici rozpětí moderních lidoopů, v intervalu asi 428 – 515 cm³, s průměrem kolem 460 cm³
- chybějící nápadné úpony silných žvýkacích a šíjových svalů (šípový hřeben crista sagittalis, šíjový hřeben crista nuchae)
- poměrně klenuté temeno
- středně pneumatizovaná spánková kost (os temporale)
- méně výrazné zúžení lebky za očnicemi (menší prostor pro spánkový sval)
- hluboká, předozadně prodloužená jamka pro připojení dolní čelisti (fossa mandibularis)
- malý bradavkový výběžek (processus mastoideus)
- primitivně dlouhý vnější zvukovod, odlišný od krátkého zvukovodu moderních lidí
- vyklenutá spodina lebeční
- vpřed posunutý velký týlní otvor (foramen magnum) v porovnání s lidoopy
- primitivně malé výstupky pro skloubení s krční páteří (condylus occipitalis)
- týlní kosti s otisky žilních splavů sulcus sinus sigmoidei a sulcus sinus transversi, odlišně od robustních australopitéků a druhu A. afarensis, ale stejně jako u rodu Homo
Znaky obličeje:
- široká, miskovitá tvář s vystupujícími lícními kostmi a plochým nosem
- vzad ubíhající čelo
- nevýrazné nadočnicové oblouky
- malá vzdálenost mezi očnicemi
- nosní otvor se zaoblenými okraji
- „přední pilíře“ - zpevňující útvary po stranách nosního otvoru, které pomáhají odolávat tlaku a namáhání lebky při skusu
- silně vyklenuté jařmové oblouky (pro dobře vyvinuté žvýkací svaly musculus masseter)
- výrazně vystupující čelisti, bližší spíše lidoopům než člověku
- chybějící bradový výběžek dolní čelisti

Znaky na zubech:
- celkově masivní žvýkací aparát
- klenutější patro než u afarských australopitéků
- parabolický tvar zubního oblouku, podobně jako u lidí
- poměrně velké řezáky
- malé, řezákovité špičáky, jen vzácně vystupující z linie ostatních zubů
- chybějící ostřící komplex[33] na hranách špičáků a třenových zubů, typický pro lidoopy
- chybějící diastema - mezera mezi druhými řezáky a špičáky horní čelisti a mezi špičáky a třenovými zuby dolní čelisti (typická pro lidoopy)
- velké zadní zuby (třenové zuby, stoličky) s výraznými hrbolky, podobně jako u rodu Homo a odlišně od paranthropů
- třenové zuby molarizované (utvářené podobně jako stoličky - symetrické, obvykle se dvěma hrbolky)
- horní třenové zuby se dvěma nebo třemi kořeny
- silná zubní sklovina

Trup
- bederní obratle tvořící bederní lordózu
- obratle s nízkým a úzkým tělem
- pánev nesoucí všechny významné znaky lidské bipední chůze, celkově bližší lidem než lidoopům
- svébytný, platypeloidní tvar pánve (z boku na bok velmi široká, ale v předozadním směru plochá)
- kyčelní kost široká, výrazně kratší než u lidoopů (ale v porovnání s lidmi stále prodloužená)
- lopaty kyčelní kosti stočené vpřed, ale ne tak výrazně jako u lidí
- dobře vyvinutý velký sedací zářez (incisura ischiadica) i přední dolní trn kyčelní (spina iliaca anterior inferior)
- velikost a rozložení svalů pánevní oblasti podobné moderním lidem
- primitivně mělká jamka kyčelního kloubu (acetabulum)

Horní končetina
- paže v poměru ke krátkým dolním končetinám relativně dlouhé (delší než u starších australopitéků)
- celková stavba a proporce ruky téměř zcela lidské
- dobře osvalený, pohyblivý palec
- mírně zakřivené články prstů jako u lidoopů
- koncové články prstů ruky opatřené drsnatinou pro masitá bříška (tuberositas phalangis distalis) jako u lidí - odlišně od starších australopitéků i moderních lidoopů

Dolní končetina
- krátká stehenní kost (jako u lidoopů), s malou hlavicí
- dlouhý, předozadně zploštělý krček stehenní kosti (s oválným průřezem)
- zvýšený bikondylární úhel, valgózní koleno – distální část stehenní kosti je výrazně ukloněná do středu těla (vzhledem k extrémně široké pánvi dokonce víc než u lidí)
- svisle postavená, nevbočená holenní kost
- chodidlo celkově plošší a pohyblivější než u lidí, ale tužší než u lidoopů, s příčnou a snad i nízkou podélnou klenbou
- palec přitažený k ostatním prstům, ale částečně ještě schopný pohybu do strany
Způsob života
.jpg.webp)
Životní prostředí
První studie pokládaly okolí jihoafrických jeskyní za suché travnaté planiny bez souvislejších stromových porostů. Lokalita Taung na okraji Kalahari byla dokonce vnímána již jako poušť nebo polopoušť. Mladší studie ale prokazují, že oblast Kolébky lidstva tvořila v pliocénu spíše pestrá mozaika ekosystémů, zahrnující otevřené travnaté pláně, ale i křoviny a prosvětlené lesy.[34] Ačkoliv savana mohla převažovat, vyskytovaly se v ní hojné ostrůvky stromových porostů. Místy lze na základě nálezů zbytků lián předpokládat i hustší subtropický les s křovinatým podrostem.[34]
Četné adaptace pro šplhání ve větvích ukazují, že A. africanus se v porovnání s východoafrickým druhem A. afarensis přizpůsobil spíše lesnatému prostředí. Jihoafričtí australopitékové tak měli patrně v oblibě úseky vlhčí pobřežní vegetace - lužní lesíky, mokřady a křoviny při březích řek a jezer, s dostatkem vody, ale současně v blízkosti otevřené krajiny.[35]
Strava

Raymond A. Dart představil zástupce A. africanus jako aktivní lovce, využívající k zabití a zpracování kořisti osteodontokeratickou kulturu - nástroje ze zvířecích kostí, zubů a rohů.[15] Tafonomické studie C. K. Braina však v druhé polovině 20. století prokázaly, že A. africanus byl spíše sám kořistí velkých šelem,[36] přičemž podle moderních studií k obživě vyhledával především rostlinné zdroje.[37]
Masivní čelisti, velké stoličky se silnou sklovinou i další úpravy obličeje vedly k předpokladu, že jihoafričtí australopitékové se soustředili na konzumaci tvrdé potravy (ořechy, semena), případně tuhé, vláknité a silně abrazivní potravy, která vyžadovala důkladné žvýkání (kořínky, hlízy).[38] Mikroskopické stopy poškození zubní skloviny, kde jemné rýhy převažují nad odštěpky a důlky, ovšem ukazují spíše na konzumaci měkkých plodů a listů než tvrdých ořechů a semen.[39]

Rovněž analýza izotopů uhlíku ze zubní skloviny prokázala, že A. africanus v hojné míře (asi z 50-75%) využíval zdroje lesních porostů a křovin, tedy plody a listy (v Africe C3-rostliny), zatímco zbylou část jídelníčku tvořily zdroje z otevřené krajiny - traviny či šáchory (v Africe C4-rostliny).[39] Zajímavé je, že tento podíl se v průběhu výskytu druhu A. africanus výrazně neměnil, a to i přes evidentní změny přírodního prostředí, představované vysušováním a ústupem porostů. To nasvědčuje představě, že A. africanus nebyl výrazně specializovaný, ale podobně jako ostatní australopitékové představoval oportunistického všežravce s širokou ekologickou tolerancí, který byl schopný využívat různorodé potravní zdroje v odlišných typech prostředí. Obýval patrně lesnaté úseky krajiny a byl dobře vybaven pro získávání potravy v korunách stromů, ale pravidelně nacházel obživu také na travnatých pláních.[40] Složení jídelníčku patrně kolísalo v závislosti na sezónní dostupnosti zdrojů, přičemž zdroje z otevřených travnatých prostor byly důležitou, ale silně variabilní součástí přijímané potravy.[5] Některé byly konzumovány po celý rok, zatímco ostatní - především tvrdé či tuhé kusy, náročné na zpracování - mohly být získávány jen sezónně, jako záložní zdroje v dobách nedostatku jiné potravy. Ty pak vzhledem k jejich krátkodobému využívání nemusely zanechat výrazné stopy na zubní sklovině.[41] Zuby také svědčí o způsobu kojení podobném lidskému.[42] Poukazují také na silnou a dlouhodobou vazbu mezi matkami a potomky.[43]
I přes převažující konzumaci rostlinné potravy nelze zcela vyloučit ani využití živočišných zdrojů. Spíše než o mase velkých savců se však uvažuje o hmyzu (především termitech), drobných obratlovcích, vejcích a podobně.[40] Konzumace svaloviny není pravděpodobná i vzhledem k nevhodnému utváření zubů.[44]
Výrazné masožravosti odporuje také analýza izotopů stroncia. Zubní sklovina nalezených jedinců obsahuje značné množství tohoto prvku, zatímco svalovina jej má jen málo. Hodnoty australopitéků odpovídají spíše rozsáhlejší konzumaci podzemních zásobních orgánů travin a šáchorovitých, které jsou bohaté na stroncium a současně chudé na baryum, jehož je i ve sklovině A. africanus malé množství. Podobné hodnoty jako A. africanus mají i moderní konzumenti oddenků trav - prase savanové nebo rypoš hotentotský.[45] Množství rýh a škrábanců na nalezených zubech australopitéků lze navíc kromě pojídání listů vysvětlit i možnou přítomností jemných abrazivních složek na povrchu přijímaného jídla, například zbytky zeminy na podzemních hlízách a oddencích.[44] Dalším možným vysvětlením naměřených hodnot stroncia a barya může být zvýšený příjem hmyzu. Hodnoty těchto prvků jsou však u moderních hmyzožravců dosud jen málo prozkoumány, chybí tak srovnávací materiál.[46]
Přesto alespoň příležitostnou konzumaci živočišných zdrojů mohou naznačovat obratle jedince ze Sterkfontein s patologickými stopami, odpovídajícími nákaze brucelózou. Tato nemoc je spojována s konzumací živočišných produktů - bakterie Brucella abortus byla izolována z ostatků moderních zeber a některých druhů antilop.[47]
Pohyb
Nalezené kosti trupu a končetin prakticky vylučují pochybnosti o vzpřímené dvojnohé chůzi u A. africanus, ačkoliv tento pohyb se podobně jako u ostatních australopitéků ještě mírně lišil od chůze moderních lidí. Přispívala k tomu méně stabilní pánev, jen nízce klenuté chodidlo, pohyblivější koleno, kotník i palec nohy.[29] Chůze tak mohla být mírně kolébavá (podobně jako u malých dětí), ve srovnání s lidmi méně výkonná a energeticky náročnější, nevhodná pro přesuny na velké vzdálenosti a nebo ve velké rychlosti.[48] Pro jihoafrické australopitéky byl ovšem takový pohyb zcela dostačující.[49] Výkonnost bipedie totiž snižovaly především četné primitivní znaky, vhodné pro pohyb ve stromech, například pohyblivější klouby nohou i rukou, zakřivené články prstů nebo prodloužené a svalnaté předloktí.[50] Lze proto předpokládat, že A. africanus na zemi využíval bipední chůzi, ale současně se častěji než lidé pohyboval v korunách stromů, kde patrně hledal potravu i úkryt před predátory. Další kroky k vylepšení bipedie by mu ztížily pohyb ve větvích, což nebylo žádoucí.[51]
Stromové patro mohlo být pro jihoafrické australopitéky dokonce významnější než pro starší východoafrické druhy.[52] Naznačuje to mimo jiné i poměr délky horních a dolních končetin, který se u A. africanus blížil spíše lidoopům než lidem. Jeho paže byly v porovnání s nohama poměrně dlouhé, zatímco východoafrický A. afarensis, který se utvářením lebky a zubů podobá člověku mnohem méně než A. africanus, již vykazuje spíše lidské proporce, tedy kratší paže i mírně prodloužené nohy.[52]
Podobně jako u druhu A. afarensis se i u A. africanus předpokládá vliv bipední chůze na rozmnožování. Platypeloidní tvar pánve je u moderních lidí velmi nevhodný pro porod, který v tom případě bývá značně komplikovaný až nemožný.[53] Pro samice australopitéků mohl být relativně bezproblémový díky malému objemu mozku, a tudíž i malé hlavičce novorozenců. Přesto již plod zřejmě nemohl procházet porodními cestami přímo jako u šimpanzů, ale byl donucen k rotaci. Někteří autoři zastávají názor, že rotace byla jen částečná, tedy že hlavička novorozence procházela pánví příčně, čelně orientována k jednomu z kyčelních kloubů (protože se v předozadně zploštělé pánvi nemohla otáčet jako u lidí).[29] Jiné studie naopak předpokládají, že pánev byla přece jen méně zploštělá než u A. afarensis, což stačilo k tomu, aby se u A. africanus již vyvinul „lidský“ způsob porodu s plnou rotací plodu.[54]
Fylogeneze
Australopithecus africanus byl jako první známý australopiték dlouho považován za přímého předka rodu Homo, případně za společného předka rodů Homo a Paranthropus. Po objevech ve východní Africe jej však v 70. a 80. letech 20. století z této pozice vytlačil Australopithecus afarensis.[55] Ačkoliv A. africanus se v mnoha znacích podobá člověku víc než A. afarensis (větší zakřivení spodiny lebeční, klenutější mozkovna, utváření zubů, méně vystupující čelisti, nevýrazné nadočnicové valy), zdá se být již příliš odvozený na to, aby se mohl stát předkem nejstarších známých lidí. Nese navíc mnohé znaky směřující spíše k robustním australopitékům (tzv. přední pilíře, velké zadní zuby, robustní spodní čelist, molarizované třenové zuby).[23] Známí zástupci rodu Paranthropus však pochází spíše z východní Afriky.[56]
Nejčastěji je tak v současnosti zastáván názor, že A. africanus je slepou vývojovou linií.[32][57] Její ukončení může představovat ještě Australopithecus sediba, který však již podle mínění většiny badatelů vyhynul bez potomků.[58] Vzhledem ke značné variabilitě nálezů ze Sterkfontein někteří autoři předpokládají, že zde mohou být zastoupeny dva druhy homininů.[29] To však dosud nebylo přesvědčivě prokázáno, takže většina autorů zatím shrnuje veškeré objevy do jediného druhu A. africanus.[29]
Vztahy A. afarensis a A. africanus zůstávají nejasné. Není zřejmé, zda může být A. afarensis předkem A. africanus nebo zda je jen vzdáleným příbuzným. Druhé možnosti by nasvědčovaly i nejstarší nálezy ze Sterkfontein, které jsou patrně současné s výskytem A. afarensis ve východní Africe.[23] Stejně tak i primitivní poměr délky končetin u A. africanus (dlouhé paže, krátké nohy) zpochybňuje, že tento druh vychází z A. afarensis, který se délkou končetin více blížil lidem i přes archaické utváření zubů a lebky.[52] Primitivní poměr délky končetin však vykazuje i Homo habilis a vzhledem ke vzácným a neúplným nálezům časných homininů zatím není zřejmý význam poměru délky končetin v evoluci člověka. Nelze vyloučit, že u A. africanus se horní končetiny druhotně opět prodloužily kvůli většímu zapojení paží do pohybu v korunách stromů.[52]
Významné nálezy

Taung
Taungské dítě je prvním nalezeným australopitékem a typovým jedincem celého rodu. Zachytil je Raymond Dart již koncem roku 1924. Nalezena byla tvář s horní čelistí, dolní čelist i přirozený výlitek mozkovny. Některé znaky - např. lidské utváření zubů, málo vyčnívající čelisti, chybějící nadočnicové valy - ukazovaly na spojitost s moderními lidmi spíše než s lidoopy. Přes počáteční striktní odmítnutí vědeckou komunitou pozdější hojné nálezy v jižní Africe prokázaly, že australopitékové patří do lidské vývojové linie. Taungské dítě zemřelo v necelých 4 letech věku a podle nových studií se stalo kořistí dravého ptáka.[59]

MLD 1
Zlomek lebky (mozkovnu) MLD 1 zachytil James Kitching v roce 1947. Jedná se o první nález australopitéka z lokality Makapansgat.[60]

TM 1511
Zlomky lebky TM 1511 jsou prvním nálezem z lokality Sterkfontein, díky němuž bylo odhaleno velmi bohaté naleziště druhu A. africanus. Zachytil je Robert Broom v roce 1936. Současně se jedná o první známé ostatky dospělého australopitéka, které prokázaly platnost druhu, navrženého v roce 1925.[7]

STS 5 (Mrs. Ples)
Lebka STS 5 byla objevena již 18. dubna 1947, ale stále představuje jednu z nejkompletnějších, nejméně poškozených lebek druhu A. africanus.[61] Nález způsobil ve své době malou senzaci a její objevitel, Robert Broom ji podle tehdejšího rodového názvu Plesianthropus překřtil na Mrs. Ples.[15] Na místě nálezu byla ukotvena pamětní deska.[62] Lebka patří k nejmladším zástupcům svého druhu - předpokládá se, že může pocházet z doby asi před 2 miliony let.[20]
Robert Broom lebku původně určil jako samičí,[63] novější průzkumy však naznačily, že by mohlo jít i o samce.[3] Protože se ale jedná o ostatky jedince na hranici dospělosti, není určení pohlaví zcela jednoznačné. Většina odborníků lebku stále považuje za samičí.[61][62]
STS 14
Částečně dochovaná kostra samice byla odhalena 1. srpna 1947. Sestává z téměř kompletní pánve, části stehenní kosti, většiny obratlů a několika zlomků žeber. Je významným zdrojem poznání způsobu pohybu australopitéků, v době objevu potvrdila předpoklad vzpřímené dvounohé chůze.[3] Protože byla zachycena poblíž lebky STS 5 a v době smrti měla přibližně stejný věk, někteří badatelé předpokládají, že může společně s lebkou patřit témuž jedinci.[62]
STW 151
Pod katalogovým číslem STW 151 jsou uloženy zlomky čelistí a navazujících částí lebky asi pětiletého dítěte. Velmi dobře je zachován mléčný a částečně prořezaný trvalý chrup. Pozoruhodné je, že se zachovaly dokonce i drobné kůstky středního ucha.[3]
STW 252
Zlomkovitě dochovaná lebka STW 252 nese v porovnání s jinými nálezy některé pozoruhodně archaické znaky včetně špičáků zřetelně přečnívajících z linie ostatních zubů.[64]
STW 431
Částečně dochovaná kostra, patřící patrně samci byla nalezena v roce 1987. Tvoří ji zlomky lopatky, klíční kosti, pažní, vřetenní i loketní kosti, třetího žebra, hrudních a bederních obratlů, křížové a pánevní kosti. Nedaleko od kostry byly zachyceny i jednotlivé zuby, které mohou patřit témuž jedinci podobně jako zlomek mozkovny, záprstní i nártní kůstky.[65] Zajímavé je, že bederní obratle nesou patologické stopy, odpovídající nákaze brucelózou, což by naznačovalo konzumaci živočišné stravy.[47]
STW 505 (Mr. Ples)
Lebka STW 505 představuje další velmi dobře dochovanou lebku, patřící patrně samci. Vzhledem k výbornému stavu dochování se jí přezdívá Mr. Ples.[66]
STW 573 (Little Foot)
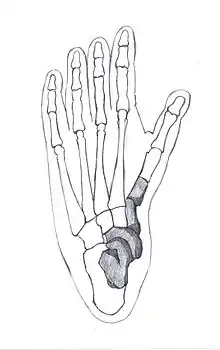
Kostra STW 573 byla nalezena v hlubší části jeskynního systému Sterkfontein, v místě zvaném Silberberg Grotto.[67] Představuje nevídaně kompletní ostatky a jednu z nejlépe dochovaných koster homininů. K dispozici jsou i velmi malé kůstky dlaní a chodidel. Významné je navíc i to, že se kostra zachovala s téměř neporušenou lebkou.[18]
První kosti byly objeveny náhodou 6. září 1994, když Ron J. Clarke zkoumal materiál získaný ve Sterkfontein ve 30. letech 20. století. V krabici s ostatky paviánů a šelem odhalil nejprve čtyři původně nepovšimnuté kosti levého chodidla.[68] O tři roky později - 19. května 1997 - znovu náhodou při práci s ostatky zvířat ze sbírek univerzity Witwatersrand zachytil část holenní kosti, pasující k předchozímu nálezu. Začal tudíž procházet i další nálezy zvířecích kostí ze stejné části vytěžené brekcie a podařilo se mu získat i dříve přehlédnuté kosti pravého chodidla. Měl tedy k dispozici celkem 12 kostí, které na sebe navazovaly a patřily témuž jedinci. Proto nabyl přesvědčení, že velká část kostry může být ještě zachována přímo ve Sterkfontein.[18] Proto se svými spolupracovníky začal pátrat v jeskyni. I přes rozsáhlý prostor, který museli prozkoumat, tmu a velmi malou šanci na úspěch Nkwane Molefe a Stephen Motsumi skutečně po pouhých dvou dnech práce dne 3. července 1997 objevili místo, odkud byly zmíněné bloky brekcie počátkem 20. století vytěženy. Ze stěny ještě vyčnívala část holenní kosti, navazující na již známé zlomky.[18] Následný výzkum odhalil téměř kompletní kostru, zvanou Little Foot podle toho, že nejprve byly zachyceny kosti chodidel.[69] Mladší geologické jevy (eroze a vertikální posun brekcie) sice způsobily zlomení stehenních kostí a propad střední části kostry do malé dutiny, jinak jsou ale ostatky neporušené a velmi dobře dochované - dochovala se lebka, obě paže i ruce, pravá lopatka a klíční kost, několik žeber a obratlů, pánev i obě nohy s chodidly.[70] Trvalo ovšem několik let, než byla kostra odhalená v celém rozsahu. Teprve poté mohla být v roce 2011 přenesena do laboratoře k dalším výzkumům.[71] Ještě před vyjmutím kostí bylo přímo v jeskyni provedeno jejich detailní nasnímaní laserovým scannerem za účelem vytvoření trojrozměrné dokumentace.[72]
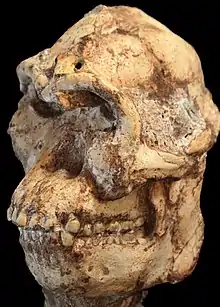
Není jasné, jestli kostra patří zástupci A. africanus nebo samostatnému druhu.[29] Nese některé svébytné znaky (protistojný, silně mobilní palec u nohy, primitivní stavba loketního kloubu, masivní jařmové oblouky). Může se jednat o předchůce druhu A. africanus nebo rodu Paranthropus.[70] Taxonomie je však zatím nejasná, podle jiných autorů kostra svým utvářením plně odpovídá druhu A. africanus.[26] Ron Clarke navrhl roku 2012 pro Little Foot používat označení Australopithecus prometheus, původně vytvořené Raymondem Dartem pro nálezy z Makapansgat.[25]
Stáří nálezu je předmětem živých debat. Nejprve bylo podle přítomností kostí vyhynulé hyeny Chasmaporthetes určeno na 3-3,5 milionu let.[24] Paleomagnetické datování tento předpoklad potvrdilo datací do doby před 3,3 miliony let.[73] Datace atomů berylia možné stáří původně posunula až na 4,17 milionu let,[17] ale podrobnější aplikace téže metody dataci opravila na 3,67 milionu let.[25] Jiné studie však předpokládají výrazně nižší stáří. Datace sintru pomocí metody uran-olovo a uran-thorium klade nález hominina do doby před 2,2 milionu let[22] Také nové využití paleomagnetismu ukázalo na stáří nejvýše 2,6 milionu let.[20] Podrobnější průzkum stratigrafie však ukázal, že sintr je výrazně mladší než brekcie a v ní uložená kostra.[74]
Dochovaná kloubní spojení ukazují, že tělo bylo překryto sedimenty ještě před úplným zetlením měkkých tkání. Kostra také nenese žádné stopy po poškození masožravci - Little Foot se nestala jejich obětí.[75] Pozice kostí (zkřížené nohy, levá ruka natažená přes hlavu) odpovídá tomu, že se tělo alespoň částečně kutálelo po hromadě sedimentů.[76] Lze proto předpokládat, že australopitékovi se stal osudným pád do jeskyně úzkým komínovým průduchem.[70]
Zajímavé je, že v blízkosti stehenní kosti australopitéka byla zachycena vřetenní kost opice. Je jedinou zvířecí kostí v těsné blízkosti Little Foot a je umístěná tak, jako by ji hominin v době smrti držel v ruce.[75]
STW 578
Lebka STW 578 byla nalezena 9. srpna 1995 v Jacovec cavern, která se nachází asi 30 m pod současným povrchem a je jedním z nejníže položených míst ve Sterkfontein. Usazeniny, vyplňující tuto prostoru, nemají žádný stratigrafický vztah k dalším částem jeskyně.[77] Lebka může patřit k nejstarším nálezům australopitéků v Africe. Datace pomocí atomů berylia udává stáří 4,0-4,2 milionu let.[17] Jednalo by se tak o jedince, současného s východoafrickým druhem Australopithecus anamensis, což by podstatným způsobem měnilo náhled na evoluci časných homininů. Datování sedimentů z Jacovec cavern však není jednoznačné, přítomnost kostí rodu Equus naznačuje, že usazeniny nemohou být starší než 2,36 milionu let (nebo byly starší sedimenty později výrazně narušeny).[78]
Mimo lebky STW 578 bylo v Jacovec cavern v letech 1995–2001 získáno dalších 11 zlomků kostí rodu Australopithecus, které se morfologicky mírně liší od nálezů ostatků A. africanus a není jasné, zda jde jen o důsledek drobných změn druhu v čase nebo zda kosti patří zcela novému druhu.[17]
Odkazy
Reference
- VANČATA, V. Paleoantropologie - přehled fylogeneze člověka a jeho předků. Brno: Akademické nakladatelství CERM, 2003. 212 s. ISBN 80-7204-272-6. S. 18.
- V současnosti je území součástí Jihoafrické republiky
- CARTMILL, M.; SMITH, M. H. The Human Lineage. New Jersey: Willey-Blackwell, 2009. 624 s. Dostupné online.
- DART, R. A. Australopithecus africanus: The man-ape of South Africa. Nature. 1925, roč. 115, s. 195–199.
- STRAIT, D. S. The evolutionary history of the australopiths. Evolution Education Outreach. 2010, roč. 3, s. 341–352.
- JELÍNEK, J. Velký obrazový atlas pravěkého člověka. 1. vyd. Praha: Artia, 1972. 560 s.
- BROOM, R. New Fossil Anthropoid Skull from South Africa. Nature. 1936, roč. 138, s. 486–488.
- BROOM, R. Further evidence on the structure of the South African Pleistocene anthropoids. Nature. 1938, roč. 142, s. 897–899.
- Ostatky hominidů v jižní Africe byly v první polovině 20. století nalézány především v důsledku intenzivní těžby vápence, který se získával z výplně starých krasových jeskyní.
- BROOM, R.; SCHEPERS, G. W. H. The South-African ape-men: the Australopithecineae. 1. vyd. Pretoria: Transvaal Museum, 1946.
- KEITH, A. Australopithecines or Dartians. Nature. 1947, roč. 159, s. 377.
- BROOM, R.; ROBINSON, J. T. Further remains of the Sterkfontein ape-man, Plesianthropus. Nature. 1947, roč. 160, s. 430–431.
- DART, R. A. The Makapansgat proto-human Australopithecus prometheus. American Journal of Physical Anthropology. 1948, roč. 6, s. 259–284.
- ROBINSON, J. T. The genera and species of the Australopithecinae. American Journal of Physical Anthropology. 1954, roč. 12, s. 181–200.
- DART, R. A.; CRAIG, D. Dobrodružství s "chybějícím článkem". 1. vyd. Praha: Mladá fronta, 1963. 304 s.
- BERGER, L. R.; TOBIAS, P. V. New discoveries at the early hominid site of Gladysvale. South African Journal of Science. 1994, roč. 90, s. 223–226. Dostupné online.
- PARTRIDGE, T. C., GRANGER, D. E., CAFFEE, M. W., CLARKE, R. J. Lower Pliocene hominid remains from Sterkfontein. Science. 2003, roč. 300, s. 607–612.
- CLARKE, R. J. First ever discovery of a well-preserved skull and associated skeleton of Australopithecus. South African Journal of Science. 1998, roč. 94, s. 460–463. Dostupné online.
- BOAZ, N. T. Paleoecology of early Hominidae in Africa. Kroeber Anthropological Society Papers. 1977, roč. 50, s. 37–62.
- HERRIES, A. I. R.; SHAW, J. Palaeomagnetic analysis of the Sterkfontein palaeocave deposits: Implications for the age of the hominin fossils and stone tool industries. Journal of Human Evolution. 2011, roč. 60, s. 523–539.
- BLACKWELL, B. A. B., SPALDING, C. N., BLICKSTEIN, J. I. B., LATHAM, A. G., QUINNEY, P., SKINNER, A. R., KUYKENDALL, K. L., REED, K. E. ESR dating the hominid-bearing breccias at the Makapansgat Limeworks Cave, South Africa. Journal of Human Evolution. 2001, roč. 40, s. A3-A4.
- PICKERING, T. R.; KRAMERS, J. D. Re-appraisal of the stratigraphy and determination of new U-Pb dates for the Sterkfontein hominin site, South Africa. Journal of Human Evolution. 2010, roč. 59, s. 70–86.
- CONROY, G. C. Reconstructing human origins. New York: W. W. Norton & company, 1997. 547 s. Dostupné online.
- BERGER, L. R.; LACRUZ, R.; DE RUITER, D. J. Revised age estimates of Australopithecus-bearing deposits at Sterkfontein, South Africa. American Journal of Physical Anthropology. 2002, roč. 119, s. 192–197. Dostupné online.
- GRANGER, D. E., GIBBON, R. J., KUMAN, K., CLARKE, R. J., BRUXELLES, L, CAFFEE, M. W. New cosmogenic burial ages for Sterkfontein Member 2 Australopithecus and Member 5 Oldowan. Nature. 2015, roč. 522, s. 85–88.
- CROMPTON, R. H.; VEREECKE, E. E.; THORPE, S. K. Locomotion and posture from the common hominoid ancestor to fully modern hominins with special reference to the common panin/hominin ancestor. Journal of Anatomy. 2008, roč. 212, s. 501–543.
- HOLLOWAY, R. L.; CLARKE, R. J.; TOBIAS, P. V. Posterior lunate sulcus in Australopithecus africanus: was Dart right?. Comptes Rendus Palevol. 2004, roč. 3, s. 287–293.
- DESOUSA, A.; CUNHA, E. Hominins and the emergence of the modern human brain. In: HOFMAN, M. A.; FALK, D. Evolution of the primate brain. Amsterdam: Elsevier, 2012. S. 293–322.
- WOOD, B. A.; RICHMOND, B. G. Human evolution: taxonomy and paleobiology. Journal of anatomy. 2000, roč. 196, s. 19–60.
- HÄUSLER, M.; MARTELLI, S.; BOENI, T. Vertebrae numbers of the early hominid lumbar spine. Journal of Human Evolution. 2002, roč. 43, s. 621–643.
- WHITCOME, K. K. Functional implications of variation in lumbar vertebral count among hominins. Journal of Human Evolution. 2012, roč. 62, s. 486–497.
- ŠMAHEL, Zbyněk. Příběh lidského rodu. Brno: Moravské zemské muzeum, 2005. 85 s. ISBN 80-7028-262-2.
- Velké horní špičáky lidoopů jsou ostřeny o třenové zuby ve spodní čelisti
- BEHRENSMEYER, A. K.; REED, K. E. Reconstructing the Habitats of Australopithecus: Paleoenvironments, Site Taphonomy, and Faunas. In: REED, K. E.; FLEAGLE, J. G.; LEAKEY, R. E. The Paleobiology of Australopithecus. [s.l.]: Springer, 2013. S. 41–60.
- REYNOLDS, S. C.; BAILEY, G. N.; KING, G. C. Landscapes and their relation to hominin habitats: case studies from Australopithecus sites in eastern and southern Africa. Journal of Human Evolution. 2011, roč. 60, s. 281–298.
- BRAIN, C. K. The hunters or the hunted?: an introduction to African cave taphonomy. 1. vyd. Chicago: University of Chicago Press, 1983. 376 s.
- DAEGLING, D. J., JUDEX, S., OZCIVICI, E., RAVOSA, M. J., TAYLOR, A. B., GRINE, F. E., TEAFORD, M. F., UNGAR, P. S. Viewpoints: Feeding mechanics, diet, and dietary adaptations in early hominins. American Journal of Physical Anthropology. 2013, roč. 151, s. 356–371.
- DAEGLING, D. J., MCGRAW, W. S., UNGAR, P. S., PAMPUSH, J. D., VICK, A. E., BITTY, E. A. Hard-object feeding in sooty mangabeys (Cercocebus atys) and interpretation of early hominin feeding ecology. PLOS ONE.. 2011, roč. 8, s. e23095.
- TEAFORD, M. F.; UNGAR, P. S. Diet and the evolution of the earliest human ancestors. Proceedings of the National Academy of Sciences. 2000, roč. 97, s. 13 506 – 13 511.
- SPONHEIMER, M.; LEE-THORP, J. A. Isotopic evidence for the diet of an early hominid, Australopithecus africanus. Science. 1999, roč. 283, s. 368–370.
- UNGAR, P. S.; SPONHEIMER, M. J. The diets of early hominins. Science. 2011, roč. 334, s. 190–193.
- https://phys.org/news/2019-07-extinct-human-species-breast-fed.html - Extinct human species likely breast fed for a year after birth, study suggests
- https://phys.org/news/2019-07-maternal-secrets-earliest-ancestors.html - Maternal secrets of our earliest ancestors unlocked
- REED, K. E.; FISH, J. L. Tropical and temperate seasonal influences on human evolution. In: BROCKMAN, D. K.; VAN SCHAIK, C. P. Seasonality in Primates: Studies of Living and Extinct Human and Non-Human Primates. Cambridge: Cambridge University Press, 2005. S. 491–520.
- SPONHEIMER, M., DE RUITER, D. J., LEE-THORP, J. A., SPÄTH, A. Sr/Ca and early hominin diets revisited: new data from modern and fossil tooth enamel. Journal of Human Evolution. 2005, roč. 48, s. 147–156.
- LEE-THORP, J. A.; SPONHEIMER, M. Contributions of biogeochemistry to understanding hominin dietary ecology. Yearbook of Physical Anthropology. 2006, roč. 49, s. 131–148.
- D'ANASTASIO, R., ZIPFEL, B., MOGGI CECCHI, I., STANYON, R., CAPASSO, L. Possible brucellosis in an early hominin skeleton from Sterkfontein, South Africa. PLOS ONE. 2009, roč. 4, s. e6439.
- SUSSMAN, R. W.; HART, D. Modeling the Past: The Primatological Approach. In: HENKE, W. C.; TATTERSALL, I. Handbook of palaeoanthropology 2. Berlín: Springer, 2007. S. 702–721.
- HARCOURT-SMITH, W. E. The Origins of Bipedal Locomotion. In: HENKE, W. C.; TATTERSALL, I. Handbook of palaeoanthropology 3. Berlín: Springer, 2007. S. 1483–1518.
- COLLARD, M.; WOOD, B. Grades among the African Early Hominids. In: BROMAGE, T. G.; SCHRENK, F. African Biogeography. Climate Change & Human Evolution. Oxford: Oxford University Press, 1999. S. 316–327.
- SANDERS, W. J. Comparative morphometric study of the australopithecine vertebral series Stw-H8/H41. Journal of Human Evolution. 1998, roč. 34, s. 249–302.
- GREEN, D. J.; GORDON, A. D.; RICHMOND, B. G. Limb-size proportions in Australopithecus afarensis and Australopithecus africanus. Journal of Human Evolution. 2007, roč. 52, s. 187–200.
- HÄUSLER, M.; SCHMID, P. Comparison of the pelvis of Sts 14 and AL 288–1: implications for birth and sexual dimorphism in australopithecines. Journal of Human Evolution. 1995, roč. 29, s. 363–383.
- BERGE, C.; GOULARAS, D. A new reconstruction of Sts 14 pelvis (Australopithecus africanus) from computed tomography and three-dimensional modeling techniques. Journal of Human Evolution. 2010, roč. 58, s. 262–272.
- MCHENRY, H. M. Mrs Ples now and then. South African Journal of Science. 1997, roč. 93, s. 165–167.
- COLLARD, M.; WOOD, B. Homoplasy and the early hominid masticatory system: inferences from analyses of extant hominoids and papionins. Journal of Human Evolution. 2001, roč. 41, s. 167–194.
- VILLMOARE, B.; KIMBEL, W. H. CT-based study of internal structure of the anterior pillar in extinct hominins and its implications for the phylogeny of robust Australopithecus. Proceedings of the National Academy of Sciences. 2011, roč. 108, s. 16 200 – 16 205.
- SPOOR, F. Malapa and the genus Homo. Nature. 2011, roč. 478, s. 44–45.
- BERGER, L. R. Brief communication: Predatory bird damage to the Taung type‐skull of Australopithecus africanus Dart 1925. American Journal of Physical Anthropology. 2006, roč. 131, s. 166–168. Dostupné online.
- TOBIAS, P. V. Some little known chapters in the early history of the Makapansgat fossil hominid site. Palaeontologia Africana. 1997, roč. 33, s. 67–79.
- GRINE, F. E., WEBER, G. W., PLAVCAN, J. M., BENAZZI, S. Sex at Sterkfontein:‘Mrs. Ples’ is still an adult female. Journal of Human Evolution. 2012, roč. 62, s. 593–604.
- THACKERAY, J. F.; GOMMERY, D.; BRAGA, J. Australopithecine postcrania (Sts 14) from the Sterkfontein Caves, South africa: the skeleton of “Mrs Ples”?. South African Journal of Science. 2002, roč. 98, s. 211–212.
- BROOM, R. Discovery of a new skull of the South African ape-man, Plesianthropus. Nature. 1947, roč. 159, s. 672.
- LOCKWOOD, C. A.; TOBIAS, P. V. A large male hominin cranium from Sterkfontein, South Africa, and the status of Australopithecus africanus. Journal of Human Evolution. 1999, roč. 36, s. 637–685.
- TOUSSAINT, M., MACHO, G. A., TOBIAS, P., PARTRIDGE, T., HUGHES, A. R. The third partial skeleton of a late Pliocene hominin (Stw 431) from Sterkfontein, South Africa. South African Journal of Science. 2003, roč. 99, s. 215–223.
- CONROY, G. C., WEBER, G. W., SEIDLER, H., TOBIAS, P. V., KANE, A., BRUNDSEN, B. Endocranial capacity in an early hominid cranium form Sterkfontein, South Africa. Science. 1998, roč. 280, s. 1730–1731.
- PICKERING, T. R.; CLARKE, R. J.; HEATON, J. L. The context of Stw 573, an early hominid skull and skeleton from Sterkfontein Member 2: taphonomy and paleoenvironment. Journal of Human Evolution. 2004, roč. 46, s. 277–295.
- CLARKE, R. J.; TOBIAS, P. V. Sterkfontein Member 2 foot bones of the oldest South African hominid. Science. 1995, roč. 269, s. 521–524.
- ACKERMANN, R. R.; SMITH, R. J. The macroevolution of our ancient lineage: what we know (or think we know) about early hominin diversity. Evolutionary Biology. 2007, roč. 34, s. 72–85.
- CLARKE, R. J. Latest information on Sterkfontein´s Australopithecus skeleton and a new look at Australopithecus. South African Journal of Science. 2008, roč. 104, s. 443–449.
- BALTER, M. Little Foot, Big Mystery. Science. 2011, roč. 333, s. 1374.
- SUBSOL, G., MORENO, B., JESSEL, J. P., BRAGA, J., BRUXELLES, L., THACKERAY, F., CLARKE, R. In situ 3D digitization of the ‘Little Foot’Australopithecus skeleton from Sterkfontein. PaleoAnthropology. 2015, čís. 44–53.
- PARTRIDGE, T. C., SHAW, J., HESLOP, D., CLARKE, R. J. The new hominid skeleton from Sterkfontein, South Africa: age and preliminary assessment. Journal of Quaternary Science. 1999, roč. 14, s. 293–298.
- BRUXELLES, L., CLARKE, R. J., MAIRE, R., ORTEGA, R., STRATFORD, D. Stratigraphic analysis of the Sterkfontein StW 573 Australopithecus skeleton and implications for its age. Journal of Human Evolution. 2014, roč. 70, s. 36–48.
- CLARKE, R. J. Discovery of complete arm and hand of the 3.3 million-year-old Australopithecus skeleton from Sterkfontein. South African Journal of Science. 1999, roč. 95, s. 477–480. Dostupné online.
- CLARKE, R. J. Newly revealed information on the Sterkfontein Member 2 Australopithecus skeleton. South African Journal of Science. 2002, roč. 98, s. 523–526.
- KUMAN, K.; CLARKE, R. J. Stratigraphy, artefact industries and hominid associations for Sterkfontein, Member 5. Journal of Human Evolution. 2000, roč. 38, s. 827–847.
- REYNOLDS, S. C.; KIBII, J. M. Sterkfontein at 75: review of palaeoenvironments, fauna and archaeology from the hominin site of Sterkfontein (Gauteng Province, South Africa). Palaeontologia Africana. 2011, roč. 46, s. 59–88.
Literatura
- ROBERTSOVÁ, A. Evoluce: Příběh člověka. Praha: Knižní klub, 2012. 256 s.
- SVOBODA, J. Čas lovců: Aktualizované dějiny paleolitu. Brno: Akademické nakladatelství CERM, 2009. 298 s.
- SVOBODA, J. Předkové. Evoluce člověka. Praha: Academia, 2014. 480 s.
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu Australopithecus africanus na Wikimedia Commons
Obrázky, zvuky či videa k tématu Australopithecus africanus na Wikimedia Commons - Trojrozměrné modely vybraných nálezů druhu Australopithecus africanus