Paranthropus
Paranthropus je rod vyhynulých homininů, žijící v pliocénu a pleistocénu (asi před 2,7–1,3 miliony let) ve východní a jižní Africe. Jeho zástupci byli současníky prvních lidí a do vývoje moderního člověka nezasáhli. Takto byli vnímáni již od prvních objevů, jak ukazuje rodové jméno, odvozené z řeckých slov para – vedle a ánthropos – člověk.[1]
Stratigrafický výskyt: pliocén, pleistocén | |
|---|---|
 Forenzní rekonstrukce tváře z lebky (2013) | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Třída | savci (Mammalia) |
| Řád | primáti (Primates) |
| Čeleď | hominidé (Hominidae) |
| Podčeleď | Homininae |
| Tribus | Hominini |
| Rod | Paranthropus Broom, 1938 |
| Druhy | |
| Některá data mohou pocházet z datové položky. | |
V širším slova smyslu patří paranthropové mezi australopitéky, mnozí odborníci pro ně používají i rodové jméno Australopithecus a řadí je k tak zvaným robustním australopitékům. Kvůli specifickým znakům ovšem bývají jinými autory vyčleňováni do samostatného rodu (Paranthropus). Proto se alternativně používají obě rodová označení a vždy záleží na postoji příslušného badatele k taxonomii a fylogenezi hominidů.
Paranthropové představují slepou vývojovou větev, přežívali však poměrně dlouho, až do spodního pleistocénu a lze je tak považovat za velmi úspěšné hominidy.
Objevy

Rodové jméno Paranthropus poprvé navrhl Robert Broom již v roce 1938 pro nálezy z krasové jeskyně Kromdraai, nacházející se nedaleko Johannesburgu v jižní Africe.[1] Objev patřil druhu Paranthropus robustus a přišel jen krátce poté, co Raymond A. Dart roku 1925 zveřejnil lebku Taungského dítěte, první známý nález australopitéka (druh Australopithecus africanus).[2] Odborná veřejnost v té době neuznávala australopitéky jako lidské předky a Robert Broom byl prakticky jediným aktivním Dartovým zastáncem, pátrajícím v jižní Africe po dalších nálezech. Dne 8. června 1938 tak získal horní čelist z Kromdraai, ovšem odlišnou od dosavadních nálezů rodu Australopithecus, a proto pro ni navrhl zcela nový druh Paranthropus robustus.[1]

V listopadu roku 1948 byly doklady paranthropa zjištěny také ve Swartkrans, vzdáleném jen asi 1,5 km západně od Kromdraai. Bylo zde získáno velké množství ostatků, vzhledem k mírným odlišnostem od nálezů z Kromdraai je ale Robert Broom popsal jako nový druh Paranthropus crassidens.[3] Dnes je však většina odborníků spojuje s nálezy z Kromdraai do jediného druhu (P. robustus).[4][5]
Koncem čtyřicátých let 20. století pak již bylo nálezů druhu Australopithecus africanus i Paranthropus robustus tolik, že odborná veřejnost konečně začala Dartovy a Broomovy objevy brát vážně.[6][7]
Roku 1955 byly podobně robustní zuby poprvé zachyceny i ve východní Africe, konkrétně v Olduvajské rokli v Tanzanii.[8] Jejich určení ale nebylo jasné až do roku 1959, kdy Mary Leakey na stejné lokalitě objevila velmi dobře dochovanou lebku s podobně utvářenými zuby. Původně byl nález popsán jako Zinjanthropus boisei.[9] Většina odborníků se však domnívala, že jde o variantu jihoafrického druhu Paranthropus robustus, a proto se již od 60. let 20. století začal uplatňovat název Paranthropus boisei, případně Australopithecus boisei.[10]
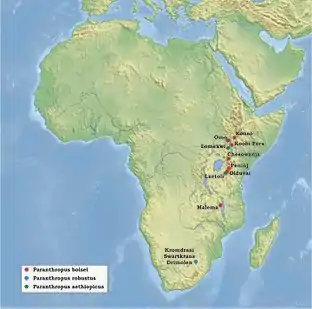
Po celou druhou polovinu 20. století pak probíhaly rozsáhlé systematické výzkumy jihoafrických lokalit Kromdraai a Swartkrans, které vydaly stovky ostatků.[11] Množství nálezů bylo současně získáno také ve východní Africe, především v okolí jezera Turkana – nejznámější lokality představuje dolní tok řeky Omo na jihu Etiopie a oblast Koobi Fora v Keni.[12]
_Mary_Douglas_Nicol_Leakey_(1913-1996)_and_her_husband_Louis_Seymour_Bazett_Leakey_(1903-1972).jpg.webp)
V roce 1967 byla navíc v údolí řeky Omo zachycena obtížně určitelná bezzubá dolní čelist.[10] Protože se svým tvarem mírně lišila od ostatních nálezů robustních australopitéků, navrhli pro ni nálezci zcela nový taxon Paraustralopithecus aethiopicus.[13] Většina tehdejších paleoantropologů ovšem nový druh neuznávala.[10] Až nález dobře dochované lebky na lokalitě Lomekwi při západním břehu jezera Turkana roku 1985[14] vedl k uznání nového druhu, ovšem pod označením Paranthropus aethiopicus.[15]
Přelom 20. a 21. století patřil v jižní Africe objevům nových jeskynních systémů, vydávajících ostatky P. robustus – jedná se o Gondolin,[16] Cooper's[17] a především o velmi bohatou lokalitu Drimolen,[18] z níž pochází i dosud nejkompletnější známá lebka australopitéka.[19] Všechny lokality se však nacházejí na relativně malé ploše v údolí Blaauwbank u Johannesburgu.[8]
Naproti tomu oblast výskytu druhu P. boisei byla novými objevy podstatně rozšířena.Tak byla v říjnu roku 1993 na etiopské lokalitě Konso zjištěna velmi dobře dochovaná lebka včetně spodní čelisti. Tím se známý areál výskytu robustních australopitéků posunul na sever.[20] Naopak jižní okraj rozšíření druhu Paranthropus boisei tvoří lokalita Malema až na severu Malawi, odkud byl v roce 1999 popsán nález části horní čelisti se silně opotřebenými stoličkami.[21]
Také areál výskytu druhu P. aethiopicus se významně rozšířil směrem k jihu, když byly jeho ostatky v roce 2001 zachyceny v tanzanském Laetoli.[22] Celkově však zůstává tento druh nejméně známým zástupcem paranthropů, doloženým jen jednotlivými zuby nebo silně poškozenými zlomky čelistí s minimem diagnostických znaků.[22]
Někteří autoři řadí k paranthropům i nově objevený a dosud málo známý druh Australopithecus garhi, odhalený v letech 1997–1998 na lokalitě Bouri v etiopské oblasti Střední Awaš.[23]
Datace
Nálezy druhu Paranthropus boisei a Paranthropus aethiopicus pocházejí z lokalit, jejichž sedimenty lze velmi dobře datovat pomocí radiometrických metod.[24] Největšího stáří dosahují ostatky P. aethiopicus – v údolí Omo je lze datovat do doby před 2,7–2,3 miliony let a ojedinělé nálezy z Lomekwi a Laetoli jsou staré asi 2,5–2,6 milionu let.[14][22]
Nejstarší nálezy P. boisei jsou známé z Malawi a dosahují stáří 2,3–2,5 milionu let.[21] Naopak nejmladší doklady tohoto druhu z etiopské lokality Konso jsou staré jen 1,4 milionu let[20] a pozůstatky částečné kostry z tanzanského Olduvai dokonce jen 1,34 milionu let.[25] V Koobi Fora a na dolním toku řeky Omo jsou ostatky P. boisei nalézány téměř z celého období výskytu tohoto druhu, tedy z doby před 2,3–1,4 milionu let.[24]
Jihoafrické krasové jeskyně jsou oproti východoafrickým lokalitám jen obtížně datovatelné. V jejich výplni chybí vrstvy sopečného popela, na něž lze dobře aplikovat radiometrické datovací metody. Uplatňují se tak především přístupy relativní chronologie. Tradiční je využití biostratigrafie, tedy datace na základě přítomnosti určitých živočišných druhů. Jejich výskyt je přitom srovnáván s dobře datovatelnými lokalitami východní Afriky.[26] Teprve v posledních letech technický rozvoj umožnil uplatnění absolutních datovacích metod i v jižní Africe, čímž se výrazně zpřesnily starší údaje, představující často značně široké časové intervaly.[27]
Nejčastěji se tak výskyt druhu Paranthropus robustus uvádí v době před 2–1,5 miliony let,[28] vyloučit však nelze ani jeho přežívání až do doby asi před 600 000 lety.[29] P. robustus tak mohl podobně jako jiné druhy savců v jižní Africe přežívat výrazně déle než ve východní části kontinentu, doklady jsou však dosud nejasné.[30]
Popis

Paranthropové jsou svébytnou skupinou hominidů, u nichž se v extrémní míře rozvinuly některé typické znaky rodu Australopithecus. Přestože jsou k dispozici stovky nálezů z východní a jižní Afriky, většinu z nich tvoří zuby nebo zlomky lebek, zatímco ostatky trupu a končetin jsou vzácné.[8] Robusticita paranthropů se týká především jejich žvýkacího aparátu, potažmo celé lebky.[31] Nález částečně dochované kostry OH 80 v Olduvai (Paranthropus boisei) však naznačuje, že i kosti končetin mohly být masivnější než u gracilních druhů.[32]
Celková stavba těla se v porovnání s ostatními australopitéky nijak zásadně neliší. Typický je výrazný pohlavní dimorfismus, srovnatelný se současnými gorilami i ostatními australopitéky.[33] Samci v průměru dosahovali výšky 130–140 cm a samice 110–124 cm. Hmotnost samců činila asi 40–50 kg, zatímco samice vážily kolem 30–35 kg.[34] Odhady celkové velikosti jsou ovšem odvozeny z velmi nekompletních ostatků, takže je nové objevy mohou změnit.[19]
I přes přetrvávající malou kapacitu je patrné mírné zvětšení mozkovny a snad i mozku (na cca 500 cm³), celková stavba se ale stále blíží spíše lidoopům než lidem. Náznaky změn v uspořádání mozku zatím nebyly jednoznačně prokázány.[35] Proto nelze předpokládat, že by duševní schopnosti paranthropů výrazně přesahovaly možnosti šimpanzů, goril nebo orangutanů. Vzhledem k velikosti mozku i k průběhu prořezávání zubů lze soudit, že ani délka a průběh ontogeneze se od moderních lidoopů příliš nelišila.[36]
Utváření lebky je v mnoha znacích podobné zástupcům rodu Homo, zatímco ostatní australopitéci jsou bližší lidoopům. Velký týlní otvor je posunutý více vpřed, spodina lebeční více klenutá. Kosti jsou silně pneumatizované, aby se snížila váha zvětšující se lebky. Mozkovnu doplňují nápadné úpony dobře vyvinutých svalů. Jedná se o šíjový hřeben crista nuchae pro šíjové svaly, vyvažující hlavu s masivními čelistmi a u větších jedinců (patrně samců) i o sagitální hřeben crista sagittalis pro spánkový sval (musculus temporalis), který umožňuje silnou práci předních zubů. Také výrazné zúžení lebky za očnicemi vytváří dostatek prostoru pro zvětšený spánkový sval.[37]
Robustní tvář je široká a díky vystupujícím lícním kostem a plochému nosu získává nápadně miskovitý tvar. Nízké, vzad ubíhající čelo je odděleno výraznými nadočnicovými oblouky.[37] Široké, vpřed silně vyklenuté lícní kosti ukazují na dobře vyvinutý zevní žvýkací sval (musculus masseter). Nosní otvor má zaoblené okraje a u druhu Paranthropus robustus jej navíc po stranách zpevňují tzv. přední pilíře, které pomáhají odolávat tlaku a namáhání lebky při skusu. Tvrdé patro tvoří značně zesílené kosti. Čelisti již nevyčnívají tak dopředu jako u starších homininů, takže celý obličej má z profilu nápadně svislý průběh, podobně jako u lidí.[38]
Masivní dolní čelist s nápadně silným tělem a vysokým ramenem (ramus mandibulae) vyplňují vedle redukovaných řezáků a špičáků především rozměrné (megadontní) třenové zuby a stoličky s velmi silnou a rychle se tvořící zubní sklovinou. Špičáky již téměř nevyčnívají z linie ostatních zubů a třenové zuby jsou molarizované, to znamená, že se svým utvářením blíží stoličkám.[5]

Málo známá je postkraniální kostra – většina poznatků je odvozována z jihoafrických nálezů.[38] Zatímco kosti nohy nesou jednoznačné doklady bipední chůze, prodloužené a svalnaté předloktí svědčí o přetrvávajícím využívání stromového patra.[39] Palec na noze, byť přitažený k ostatním prstům, si ještě zachovával větší rozsah pohybu než u lidí. Utváření ruky se však již značně přiblížilo člověku, zejména pro druh Paranthropus robustus jsou doložené rovné, nikoliv primitivně zahnuté články prstů. To spolu s dobře vyvinutým a osvaleným, pohyblivým palcem odpovídá zvýšeným manipulačním schopnostem.[40]
Rekonstrukce Forenzní trojrozměrnou rekonstrukci tváře paranthropa boisei podle 3D skenu lebky od Dr. Moacira Eliase Santose provedl roku 2013 brazilský specialista Cicero Moraes.
Způsob života
Životní prostředí
Zástupci rodu Paranthropus jsou ve východní i jižní Africe spojováni s vysušováním prostředí, šířením suchomilné vegetace a změnou vlhkých, prosvětlených porostů v otevřenou suchou savanu.[41] Klimatické změny vyvrcholily zhruba před 2,8 – 2,5 miliony lety současně s nástupem první doby ledové v Evropě.[42]
Přestože převažujícím ekosystémem v okolí pleistocénních lokalit byly bezlesé travnaté pláně, velká část badatelů předpokládá, že paranthropové využívali i přetrvávající ostrůvky podmáčených luk nebo mokřin při březích jezer a vodních toků.[43] V těchto vlhčích místech se místy mohla udržet hustší vegetace, křoviny a řídké lesíky, skýtající kromě potravy i možnost úkrytu před dravci.[44] Zatím nelze spolehlivě prokázat, zda paranthropové upřednostňovali otevřenou travnatou krajinu nebo spíše mokřady a lesíky. Zdá se, že byli schopní využívat větší spektrum prostředí, přičemž určujícím faktorem nebyla přítomnost nebo absence stromového porostu, ale blízkost vodního zdroje.[44]
Strava
Nápadně mohutná, specializovaná stavba čelistí a zubů i unikátně rychlá tvorba skloviny od prvních objevů soustředila pozornost odborníků na stravu paranthropů. Zpočátku převládal názor, že paranthropové byli na rozdíl od gracilních australopitéků a prvních lidí výhradně býložraví a úzce specializovaní na konzumaci tvrdé a křehké nebo tuhé potravy. Také přezdívka Louskáček, která se vžila pro lebku OH 5 z Olduvai, vychází z tohoto předpokladu.[45]

Od konce 20. století jsou ovšem starší závěry zřetelně upravovány díky výsledkům moderních analýz. Mezi ně patří studium poměru izotopů stroncia a vápníku, izotopů uhlíku nebo studium mikroskopického poškození zubní skloviny. Ukazuje se, že i přes podobnou morfologii tváře, lebky a zubů se strava východoafrických a jihoafrických paranthropů zřejmě nápadně lišila a že P. robustus a P. boisei využívali odlišné potravní zdroje.[45] Paranthropus robustus získával téměř 75% potravy v lesních porostech a v křovinách. Poměr využití zdrojů ze stromů či keřů a z otevřených travnatých prostor u něj přitom značně kolísal.[10] Paranthropus boisei naopak 75–80 % potravy vyhledával v prostoru savany a tento podíl se neměnil ani v průběhu dlouhého výskytu druhu.[45] Paranthropus robustus má sklovinu poškozenou především početnými odštěpky a důlky, odpovídajícími tvrdé stravě, zatímco P. boisei nese na zubech spíše jemné rýhy a škrábance, ukazující na žvýkání tuhé, vláknité stravy.[43]
Proto se nejčastěji uvažuje o tom, že Paranthropus boisei se v pobřežních mokřinách živil převážně rostlinami z čeledi šáchorovitých, na něž se zčásti specializoval.[41] Jen menší část potravy pak mohlo tvořit ovoce, semena, ořechy, případně hmyz nebo drobní obratlovci. Naproti tomu Paranthropus robustus byl (podobně jako jiní australopitéci i časní zástupci rodu Homo) spíše oportunistickým všežravcem s širokou ekologickou tolerancí a rozsáhlou potravinovou základnou.[46]
Paranthropové tak různými způsoby reagovali na klimatické změny, způsobující šíření nekvalitní, tuhé a tvrdé nebo málo výživné potravy na úkor snadno dostupných a měkkých plodů. Základem jídelníčku přitom nemusela být vždy potrava s velkými nároky na zpracování. Konzumováno mohlo být i snadno dostupné, na živiny bohaté ovoce nebo jiné plody, přičemž tvrdé a tuhé kusy mohly být získávány jen jako záložní zdroje v dobách nedostatku jiné potravy.[45] Všichni zástupci rodu Paranthropus však již více než starší australopitékové pronikali do otevřené bezlesé krajiny, kde byli schopní využívat nové potravní zdroje jako například termitiště nebo podzemní zásobní orgány různých rostlin (hlízy, oddenky, cibulky).[38]
Kultura
V počátcích výzkumů byli paranthropové v porovnání s člověkem vnímáni jako primitivní, býložraví tvorové s velkými zuby a malým mozkem, jejichž inteligence nestačila na nástrojovou činnost. Veškeré nalézané artefakty tak byly automaticky přisuzovány soudobým zástupcům rodu Homo a spojovány se získáváním masité potravy.[47] Takové vnímání se však postupně mění, zejména vlivem jihoafrických nálezů. Utváření ruky (alespoň u P. robustus) odpovídá schopnosti jemné manipulace.[38] Užívání nástrojů se navíc předpokládá i u jiných druhů australopitéků (A. afarensis, A. garhi), stejně jako je známé u všech moderních lidoopů i některých primátů.[48]
Dosud neexistují jednoznačné doklady, že by Paranthropus aethiopicus a Paranthropus boisei používali nebo vyráběli nástroje. Louis Leakey sice po nálezu Louskáčka v roce 1959 pokládal P. boisei za tvůrce oldovanských nástrojů, později však bylo prokázáno, že v oblasti žil ve stejnou dobu také Homo habilis a je pravděpodobnější, že právě on je původcem těchto artefaktů. Přesto nelze vyloučit, že P. boisei k získávání podzemních oddenků a hlíz nějaké hole nebo kopáče využíval.[10]
Druhu Paranthropus robustus jsou často přisuzovány kostěné hroty, nalezené ve Swartkrans a Drimolen. Úspěšné experimenty prokázaly vhodnost nástrojů k vykopávání podzemních hlíz nebo k rozbíjení termitišť a získávání termitů. Vzhledem k současnému výskytu ostatků rodů Homo a Paranthropus na těchto lokalitách je ovšem obtížné přiřadit tyto nástroje konkrétnímu druhu.[49]
.JPG.webp)
Fylogeneze
Převážná část odborníků se shoduje, že paranthropové tvoří monofyletickou skupinu se společným předkem. Tomu zatím nasvědčuje i většina srovnávacích studií.[24][50] Geologicky nejstarší druh Paranthropus aethiopicus byl ovšem rozeznán jako poslední, a proto jeho nález zásadním způsobem změnil představy o vývoji celé skupiny.[51] Starší studie předpokládaly, že Paranthropus robustus se vyvinul v jižní Africe z druhu Australopithecus africanus a poté se stal předkem východoafrického Paranthropus boisei, který později vyhynul bez potomků.[10] Po nálezu Černé lebky (Paranthropus aethiopicus) roku 1985 je však zřejmé, že předka robustních australopitéků je nutné hledat spíše ve východní Africe. Paranthropus aethiopicus totiž utvářením lebky i dobou výskytu velmi dobře překlenuje rozdíly mezi Paranthropus robustus a Australopithecus afarensis a spojuje tak gracilní a robustní australopitéky do jedné vývojové linie.
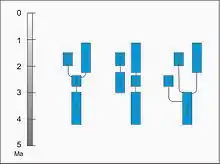
Proto podle nejčastějšího názoru na druh A. afarensis navázal P. aethiopicus, z něhož vyšel jak východoafrický P. boisei, tak jihoafrický P.robustus.[52] Méně časté jsou představy, že se P. aethopicus odštěpil ještě dříve než P. boisei a P. robustus, kteří by tak sice rovněž měli původ ve východní Africe, ale nenavazovali by na P. aethiopicus.[53] Třetí možností je, že robustní australopitékové nesdílejí společného předka, tedy že P. aethiopicus a P. boisei navazují ve východní Africe na druh Australopithecus afarensis, zatímco P. robustus se v jižní Africe vyvinul samostatně z druhu Australopithecus africanus.[54] Obě dvě poslední varianty ovšem předpokládají, že robustní žvýkací aparát se u těchto homininů vyvinul nezávisle na sobě, paralelním vývojem.[55]
Robustní australopitékové a první zástupci rodu Homo se ve fosilním záznamu objevují téměř současně a podporují tak předpoklad štěpení pliocenních homininů na dvě souběžné vývojové linie, představující patrně dva různé způsoby adaptace na klimatické změny.[42] Podle tradičního náhledu došlo k rozdělení na specializované býložravce s malým mozkem (Paranthropus) a generalizované, přizpůsobivé všežravce s velkým mozkem (Homo).[56] Zároveň se předpokládalo, že paranthropové se v pleistocénu již nedokázali přizpůsobit dalšímu vysychání a souvisejícím změnám prostředí, nedokázali najít dostatek potravy, a proto vyhynuli. Ukazuje se však, že byli velmi úspěšnou skupinou, přežívající dlouho do spodního pleistocénu a rozhodně je nelze považovat za zaostalé. Ostatky P. boisei jsou dokonce na známých lokalitách výrazně početnější než nálezy soudobých zástupců rodu Homo. Alespoň jihoafričtí paranthropové dokázali využít široké spektrum potravních zdrojů, byli tedy schopní najít obživu v mnoha různých ekosystémech podobně jako lidé.[46] Je možné, že potravní niky obou skupin se částečně překrývaly a lišit se mohly jen záložní zdroje, využívané v nepříznivých obdobích roku. Zatímco Homo mohl častěji využívat živočišné zdroje, Paranthropus se spoléhal na tuhé nebo tvrdé jídlo, pro jiné živočichy nedostupné.[46]
Důvod vymizení robustních australopitéků proto dosud nebyl uspokojivě vysvětlen.[38] Problémem mohl být kromě potravy také nedostatek vody, protože paranthropové vykazují úzkou vazbu na vodní zdroje a vlhčí biotopy.[30] Za vyhynutím paranthropů (respektive australopitéků) ovšem nemusely stát jen samotné klimatické změny, ale i neschopnost konkurovat prasatům, opicím či mladším formám rodu Homo (především Homo erectus).[57] Někteří dokonce spekulují, že Homo erectus australopitéky lovil a vyhubil.[30] Vyhynutí paranthropů ale nemusí být nutně důsledkem cíleného lovu, nýbrž jen zvýšené kompetice. Homo erectus mohl jen častěji a s větším úspěchem využívat stejné potravní zdroje. Například díky lepší schopnosti plánování mohl záměrně stopovat budoucí kořist a nespoléhat jen na náhodné nálezy nebo díky kamenným nástrojům dokázal rozbít zvířecí kosti a využít morek, nejen maso.[58] Doklady pro kontakty obou skupin homininů však zatím chybí.[38]
Odkazy
Reference
- BROOM, R. The genera and species of the South African fossil ape man. American Journal of Physical Anthropology. 1950, roč. 8, s. 1–13.
- DART, R. A. Australopithecus africanus: The man-ape of South Africa. Nature. 1925, roč. 115, s. 195–199.
- BROOM, R. Another new type of fossil ape-man. Nature. 1949, roč. 163, s. 57.
- ROBINSON, J. T. The genera and species of the Australopithecinae. American Journal of Physical Anthropology. 1954, roč. 12, s. 181–200.
- STRAIT, D. S.; GRINE, F. E.; MONIZ, M. A. A reappraisal of early hominid phylogeny. Jounal of Human Evolution. 1997, roč. 32, s. 17–82.
- KEITH, A. Australopithecines or Dartians. Nature. 1947, roč. 159, s. 377.
- LE GROS CLARK, W. Observation on the anatomy of the fossil Australopithecinae. Journal of Anatomy. 1947, roč. 83, s. 300–333.
- WOOD, B. A.; RICHMOND, B. G. Human evolution: taxonomy and paleobiology. Journal of anatomy. 2000, roč. 196, s. 19–60.
- LEAKEY, L. S. B. A new fossil skull from Olduvai. Nature. 1959, roč. 184, s. 491–493.
- CARTMILL, M.; SMITH, F. H. The Human Lineage. 1. vyd. Oxford: Wiley-Blackwell, 2009. 624 s. Dostupné online. ISBN 978-0471214915.
- GRINE, F. E. New craniodental fossils of Paranthropus from the Swartkrans Formation and their significance in ‘‘robust’’ australopithecine evolution. In: GRINE, F. E. Evolutionary History of the ‘‘Robust’’ Australopithecines. [s.l.]: [s.n.], 1988. S. 223–243.
- CONSTANTINO, P.; WOOD, B. A. The evolution of Zinjanthropus boisei. Evolutionary Anthropology: Issues, News, and Reviews. 2007, roč. 16, s. 49–62.
- ARAMBOURG, C.; COPPENS, Y. Découverte d’un australopithécien nouveau dans les gisements de l’Omo (Éthiopie). South African Journal of Science. 1968, roč. 64, s. 58–59.
- WALKER, A., LEAKEY, R. E. F., HARRIS, J. M., BROWN, F. H. 2.5 Myr Australopithecus boisei from west of Lake Turkana, Kenya. Nature. 1986, roč. 322, s. 517–522.
- KIMBEL, W. H.; WHITE, T. D.; JOHANSON, D. C. Implications of KNM-WT 17000 for the evolution of ‘‘robust’’ Australopithecus. In: GRINE, F. E. Evolutionary History of the ‘‘Robust’’ Australopithecines. [s.l.]: Aldine Transaction, 2007. S. 259–268.
- MENTER, C. G., KUYKENDALL, K. L.; KEYSER, A. W.; CONROY, G. C. First record of hominid teeth from the Plio-Pleistocene site of Gondolin, South Africa. Journal of Human Evolution. 1999, roč. 37, s. 299–307.
- BERGER, L. R., DE RUITER, D. J.; STEININGER, C. M.; HANCOX, J. Preliminary results of excavations at the newly discovered Coopers D deposit, Gauteng, South Africa. South African Journal of Science. 2003, roč. 99, s. 276–278.
- KEYSER, A. W., MENTER, C. G.; MOGI-CECCHI, J.; PICKERING, T. R.; BERGER, L. R. Drimolen: A new hominid-bearing site in Guateng, South Africa. South African Journal of Science. 2000, roč. 96, s. 193–197. Dostupné online.
- KEYSER, A. W. The Drimolen skull: the most complete australopithecine cranium and mandible to date. South African Journal of Science. 2000, roč. 96, s. 189–193. Dostupné online.
- SUWA, G., ASFAW, B., BEYENE, Y., WHITE, T. D., KATOH, S., NAGAOKA, S., NAKAYA, H., UZAWA, K., RENNE, P., WOLDEGABRIEL, G. The first skull of Australopithecus boisei. Nature. 1997, roč. 389, s. 489–492.
- KULLMER, O., SANDROCK, O., ABEL, R., SCHRENK, F., BROMAGE, T. G., JUWAYEYEI, Y. M. The first Paranthropus from the Malawi Rift. Journal of Human Evolution. 1999, roč. 37, s. 121–127.
- HARRISON, T. Hominins from the Upper Laetolil and Upper Ndolanya Beds, Laetoli. In: HARRISON, T. Paleontology and Geology of Laetoli: Human Evolution in Context 2. [s.l.]: Springer, 2011. S. 141–188.
- WOOD, B. A.; LONERGAN, N. L. The hominin fossil record: taxa, grades and clades. Journal of Anatomy. 2008, roč. 212, s. 354–376.
- WOOD, B. A.; CONSTANTINO, P. Paranthropus boisei: fifty years of evidence and analysis. American journal of physical anthropology. 2007, roč. 134/S45, s. 106–132.
- DOMÍNGUEZ-RODRIGO, M., PICKERING, T. R., BAQUEDANO, E., MABULLA, A., MARK, D. F., MUSIBA, C., et al. First partial skeleton of a 1.34-million-year-old Paranthropus boisei from Bed II, Olduvai Gorge, Tanzania. PloS one. 2013, roč. 8, s. e80347.
- BOAZ, N. T. Paleoecology of early Hominidae in Africa. Kroeber Anthropological Society Papers. 1977, roč. 50, s. 37–62.
- HERRIES, A. I. R.; CURNOE, D.; ADAMS, J. W. A multi-disciplinary seriation of early Homo and Paranthropus bearing palaeocaves in southern Africa. Quaternary International. 2009, roč. 202, s. 14–3 28.
- KIMBEL, W. H. The species and diversity of australopiths. In: HENKE, W.; TATTERSALL, I. Handbook of Palaeoanthropology 3. Berlín: Springer, 2007. S. 1539–1573.
- HERRIES, A. I. R.; ADAMS, J. W. Clarifying the context, dating and age range of the Gondolin hominins and Paranthropus in South Africa. Journal of Human Evolution. 2013, roč. 65, s. 676–681.
- KLEIN, R. G. The causes of “robust” australopithecine extinction. In: GRINE, F. E. Evolutionary History of the “Robust” Australopithecines. [s.l.]: Aldine transactions, 2007. S. 499–505.
- SVOBODA, J. A. Čas lovců: Aktualizované dějiny paleolitu. 1. vyd. Brno: Akademické nakladatelství CERM, 2009. 298 s.
- DOMÍNGUEZ-RODRIGO, M., PICKERING, T. R., BAQUEDANO, E., MABULLA, A., MARK, D. F., MUSIBA, C., et al. First partial skeleton of a 1.34-million-year-old Paranthropus boisei from Bed II, Olduvai Gorge, Tanzania. PloS one. 2013, roč. 8, s. e80347.
- WOOD, B. A. Human evolution. Bioessays. 1996, roč. 18, s. 945–954. Dostupné online.
- ROBERTSOVÁ, A. Evoluce: Příběh člověka. Praha: Knižní klub, 2012. 256 s.
- DESOUSA, A.; CUNHA, E. Hominins and the emergence of the modern human brain. In: HOFMAN, M. A.; FALK, D. Evolution of the primate brain. [s.l.]: Elsevier, 2012. S. 293–322.
- BOGIN, B.; SMITH, B. H. The evolution of the human life cycle. American Journal of Human Biology. 1996, roč. 8, s. 703–716.
- CONROY, G. C. Reconstructing human origins. New York: W. W. Norton & company, 1997. 547 s. Dostupné online.
- CONSTANTINO, P. The “robust” australopiths. Nature Education. 2013, roč. 4, s. 1.
- COLLARD, M.; WOOD, B. Grades among the African Early Hominids. In: BROMAGE, T. G.; SCHRENK, F. African Biogeography. Climate Change & Human Evolution. Oxford: Oxford University Press, 1999. S. 316–327.
- CROMPTON, R. H.; VEREECKE, E. E.; THORPE, S. K. S. Locomotion and posture from the common hominoid ancestor to fully modern hominins with special reference to the common panin/hominin ancestor. Journal of Anatomy. 2008, roč. 212, s. 501–543.
- STEWART, K. M. Environmental change and hominin exploitation of C4-based resources in wetland/savanna mosaics. Journal of Human Evolution. 2014, roč. 77, s. 1–16.
- DE MENOCAL, P. B. Plio-Pleistocene African climate. Science. 1995, roč. 270, s. 53–59.
- GRINE, F. E., SPONHEIMER, M. J., UNGAR, P. S., LEE-THORP, J., TEAFORD, M. F. Dental microwear and stable isotopes inform the paleoecology of extinct hominins. American Journal of Physical Anthropology. 2012, roč. 148, čís. 285–317.
- DE RUITER, D. J.; SPONHEIMER, M.; LEE-THORP, J. A. Indications of habitat association of Australopithecus robustus in the Bloubank Valley, South Africa. Journal of Human Evolution. 2008, roč. 55, s. 1015–1030.
- UNGAR, P. S.; SPONHEIMER, M. J. The diets of early hominins. Science. 2011, roč. 334, s. 190–193.
- WOOD, B. A.; STRAIT, D. S. Patterns of resource use in early Homo and Paranthropus. Journal of Human Evolution. 2004, roč. 46, s. 119–162.
- SUSMAN, R. L. New postcranial remains from Swartkrans and their bearing on the functional morphology and behavior of Paranthropus robustus. In: GRINE, F. E. Evolutionary History of the Robust Australopithecines. [s.l.]: [s.n.], 1988. S. 149–172.
- CONSTANTINO, P.; WOOD, B. A. Paranthropus paleobiology. Miscelánea en homenaje a Emiliano Aguirre. 2004, roč. 3, s. 136–151.
- BACKWELL, L. R.; D'ERRICO, F. Early hominid bone tools from Drimolen. Journal of Archaeological Science. 2008, roč. 35, s. 2880–2894.
- VILLMOARE, B.; KIMBEL, W. H. CT-based study of internal structure of the anterior pillar in extinct hominins and its implications for the phylogeny of robust Australopithecus. Proceedings of the National Academy of Sciences. 2011, roč. 108, s. 16 200 – 16 205.
- DELSON, E. Human phylogeny revised again. Nature. 1986, roč. 322, s. 496–497.
- COLLARD, M.; WOOD, B. Homoplasy and the early hominid masticatory system: inferences from analyses of extant hominoids and papionins. Journal of Human Evolution. 2001, roč. 41, s. 167–194.
- STRAIT, D. S.; GRINE, F. E.; MONIZ, M. A. A reappraisal of early hominid phylogeny. Journal of Human Evolution. 1997, roč. 32, s. 17–82.
- POTTS, R. Environmental hypotheses of Pliocene human evolution. In: BOBE, R.; ALEMSEGED, Z.; BEHRENSMEYER, A. K. Hominin Environments in the East African Pliocene: An Assessment of the Faunal Evidence. [s.l.]: Springer, 2007. S. 25–49.
- LEWIN, R. New fossil upsets human family. Science. 1986, roč. 233, s. 720–721.
- COPPENS, Y. Hominid evolution and the emergence of the genus Homo. Pontifical Academy of Sciences Scripta Varia. 2013, roč. 121, s. 1–15.
- ŠMAHEL, Z. Příběh lidského rodu. 1. vyd. Brno: Moravské zemské muzeum, 2005. 85 s. ISBN 80-7028-262-2.
- SPONHEIMER, M.; LEE-THORP, J. A. Differential resource utilization by extant great apes and australopithecines: towards solving the C-4 conundrum. Comparative Biochemistry and Physiology A. 2003, roč. 136, s. 27–34.
Literatura
- MAZÁK, V. Sága rodu Homo. Praha: Práce, 1986. 424 s.
- ROBERTSOVÁ, A. Evoluce: Příběh člověka. Praha: Knižní klub, 2012. 256 s.
- SVOBODA, J. Čas lovců: Aktualizované dějiny paleolitu. Brno: Akademické nakladatelství CERM, 2009. 298 s.
- SVOBODA, J. Předkové. Evoluce člověka. Praha: Academia, 2014. 480 s.
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu Paranthropus na Wikimedia Commons
Obrázky, zvuky či videa k tématu Paranthropus na Wikimedia Commons