Membránový potenciál
Membránový potenciál je rozdíl elektrického potenciálu mezi dvěma stranami biologické membrány. Z fyzikálního hlediska je to vlastně napětí na polarizované polopropustné membráně. Vzniká jako důsledek působení elektrochemického gradientu malých iontů a protonů.
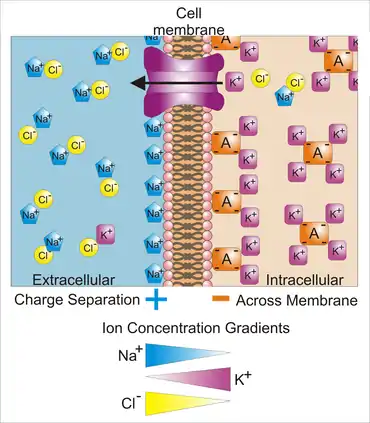
Z vlastností polopropustné membrány totiž vyplývá, že průchod většiny látek není ve většině případů možný volně. Volně mohou přes membránu procházet pouze ty molekuly, které jsou rozpustné v tucích, a takové molekuly, které jsou pouze slabě polarizovány (voda, močovina, glycerol, oxid uhličitý) – a to pouze v místech silného zakřivení nebo při změně elektrického pole. Nabité částice mohou přes membránu projít za předpokladu, že pro ně existuje buď kanál, kterým by mohly projít, nebo specifický přenašeč.
Z toho plyne i nerovnoměrné rozdělení iontů vně a uvnitř membrány.
Klidový membránový potenciál
Mezi vnější a vnitřní částí cytoplasmatické membrány naprosté většiny živých buněk existuje klidový membránový potenciál, jehož hodnota je –30 až –90 mV (u většiny neuronů v lidském těle: -70 až -90 mV). Rozdíly v koncentracích iontů vně a uvnitř buňky způsobí, že vnitřní povrch membrány nese záporný náboj díky organickým iontům (glutamát, aspartát), vnější povrch pak náboj kladný. Klidový membránový potenciál je třeba chápat jako výsledek rovnováhy, která se ustaví na základě koncentračního a elektrického gradientu jednotlivých iontů. Uvnitř buňky je značné množství bílkovin, které nemohou procházet přes membránu a které nesou záporný náboj. To je výchozí stav způsobující zápornost vnitřku buňky.
Koncentrace iontů v extracelulárním prostoru se velmi liší od koncentrace iontů uvnitř buňky:
| Intracelulární koncentrace [mmol/l] | Extracelulární koncentrace [mmol/l] | ||
|---|---|---|---|
| Na+ | 12 | Na+ | 145 |
| K+ | 155 | K+ | 4 |
| Ca2+ | 10−5 – 10−4 | Ca2+ | 2 |
| Cl− | 4 | Cl− | 120 |
| HCO3− | 8 | HCO3− | 27 |
| bílkoviny (A−) | 155 | bílkoviny (A−) | 0 |
Důsledky jsou následující:
- Kationty jsou elektricky nuceny ke vstupu do nitra buňky.
- To je možné prakticky pouze pro K+ ionty. Na+ nemůže vzhledem k velikosti svého hydratačního obalu volně procházet „stále otevřenými“ iontovými kanály, což K+ může (dáno specifitou draselného kanálu, který může přenášet pouze K+ s větším iontovým poloměrem).
- Anionty (Cl−) jsou naopak negativním vnitřkem buňky odpuzovány.
- Koncentrační gradient naopak žene K+ z nitra ven.
- Na+ je aktivně čerpán ven (3 ionty) a K+ dovnitř buňky (2 ionty) Na+/K+ATP–ázou, což elektronegativitu membrány teoreticky zvyšuje o 10 mV (kdyby toho nebylo, byla by membrána méně negativní).
Konečný klidový membránový potenciál můžeme tedy chápat jako výsledek protichůdného (nebo souhlasného) působení výše uvedených sil. Tato situace platí pro většinu buněk. Existují však buňky, u nichž není možné hovořit o klidovém membránovém potenciálu – tedy takové, které svoji polaritu neustále mění – pacemakerové buňky. Pro ně by pak byly výše uvedené podmínky změněny a hovoří se o prostupech iontů „stále otevřenými“ iontovými kanály. Podle tohoto kritéria, tj. zda buňka mění či nemění v klidu rozdíl napětí mezi vnější a vnitřní stranou membrány, je možné buňky rozdělit na:
- buňky zcela polarizované (většina);
- buňky neúplně polarizované (pacemakerové buňky).
U buněk neúplně polarizovaných je chybné hovořit o klidovém membránovém potenciálu, neboť neustále mění rozdíl polarity mezi vnější a vnitřní stranou membrány.
Změny potenciálu
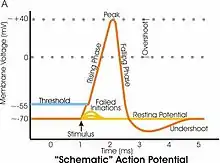
Hyperpolarizace a depolarizace
Hyperpolarizace je zvyšování membránového potenciálu (zvyšování rozdílu potenciálů), vzniká tedy více negativní membránový potenciál. Hyperpolarizace u nervových buněk zabraňuje vzniku nervového impulzu (akčního potenciálu) a má tedy uklidňující účinek na nervové buňky.
Depolarizace je snižování membránového potenciálu směrem k nulovým hodnotám. Depolarizace typicky probíhá při vzrušení nervových buněk jako pokles potenciálu směrem k nule – taková depolarizace je nutná pro vznik akčních potenciálů a tedy i pro šíření vzruchů.
Akční potenciál
Na membránách nervových a svalových buněk dochází k dočasným, vratným a rychlým změnám membránového potenciálu, které umožňují přenos vzruchu a svalový stah. To se označuje jako akční potenciál.
Stanovení
Měřením membránového potenciálu je možné nepřímo sledovat fyziologické a biochemické pochody uvnitř buňky, případně její interakci s vnějším prostředím. Za podmínek rovnováhy lze ze známé koncentrace vybraného iontu (membrána pro něj musí být propustná) uvnitř a vně buňky přímo spočítat hodnotu membránového potenciálu Nernstova rovnice:
- Ψ = k . ln (ce/ci),
kde Ψ je membránový potenciál ve voltech, k je konstanta závislá na teplotě a na náboji daného iontu; druhý výraz v rovnici je přirozený logaritmus podílu extracelulární a intracelulární koncentrace iontu. Konstanta k je definována vztahem k = R T/z F, kde R je univerzální plynová konstanta, T je absolutní teplota, z je valence iontu a F Faradayova konstanta. V případě neurálních buněk lze užít Goldman-Hodgkin-Katzova rovnice:
