Apoptóza
Apoptóza je jeden z hlavních typů programované buněčné smrti (PCD, někdy je s ní dokonce synonymizována),[1] probíhající především u živočichů, ale v určitém slova smyslu i u rostlin a dalších skupin.[2] Zahrnuje sled biochemických procesů vedoucích k typickým změnám vzhledu buňky (degradace cytoskeletu a následná změna tvaru cytoplazmatické membrány, smrštění buňky, fragmentace jádra i chromozomů uvnitř). Následně dochází k šetrnému odstranění zbytků této buňky (a nikoliv k zánětu), čímž se apoptóza v základech liší od nekrózy. Mezi nekrózou a apoptózou však není ostrá hranice a často se tyto jevy kombinují, čímž dochází k nejednoznačně pojmenovatelným typům buněčné smrti.[1]
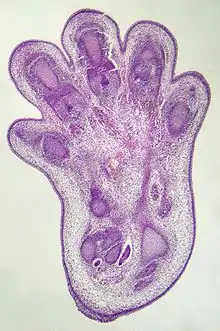
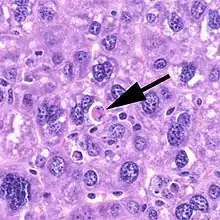
Apoptózu lze v mnohých případech vnímat jako pozitivní proces, ke kterému například lidské tělo sahá v průběhu svého vývoje (známým příkladem je proces vzniku prstů tak, že buňky mezi budoucími prsty spáchají apoptózu). Apoptoticky však umírají i různé poškozené buňky, a to buď na základě „vlastního rozhodnutí“ samotné buňky (vnitřní cesta), nebo díky buňkám imunitního systému.
Od počátku devadesátých let se množství publikací věnovaných tomuto tématu prudce zvýšilo. Mimo skutečnost, že je to významný biologický fenomén jako takový, je dnes známo, že porucha schopnosti apoptózy může u člověka způsobit poměrně široké spektrum onemocnění. Nadměrná apoptóza může vést k hypotrofii tkáně (některé typy ischemie), naopak nedostatečná apoptóza může obecně způsobit nadměrné pomnožení buněk (proliferaci), a tak například vést k rakovině.
Objev apoptózy a původ slova
Programovanou buněčnou smrt poprvé popsali vědci již v devatenáctém století. Německý přírodovědec Carl Vogt byl v roce 1842 pravděpodobně prvním, kdo popsal základní princip apoptózy. Podrobnější schéma tohoto procesu později v roce 1885 popsal německý cytogenetik Walther Flemming. Znovu se však vědci začali zabývat tímto fenoménem až v druhé polovině dvacátého století. John F. R. Kerr a jeho kolegové v roce 1972[3] poprvé použili termínu apoptóza (z řec. apoptosis — padání).[4] Samotné slovo apoptóza je totiž inspirováno skutečností, že při padání listů na podzim musí dojít k apoptotickému odumření vrstvy buněk v řapíku.[5] Není však bez zajímavosti, že termínu apoptóza již ve starověku používali Hippokratés či Galénos ve smyslu „plešatění“, „řídnutí kostí“ nebo „odpadávání strupů“.[6]
Apoptóza je v současnosti velmi intenzívně zkoumána. Počet publikací o apoptóze roste geometrickou řadou[7] a roku 2005 je o apoptóze více než 100 000 vědeckých článků. Roku 2002 dostali Sydney Brenner, H. Robert Horvitz a John E. Sulston Nobelovu cenu za fyziologii a lékařství za výzkum v oblasti genetické regulace vývoje orgánů a programované buněčné smrti.[8]
Morfologické změny buňky
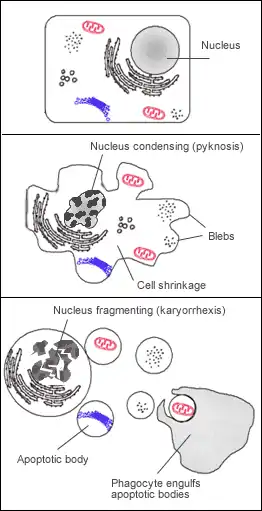
Buňka procházející apoptózou se odlišuje od běžných buněk zejména těmito rysy:
- Smrštění buňky[9]
- Blebbing membrány (váčkovité výrůstky z membrány),[10] způsobený rozkladem kortikálního cytoskeletu (tvořen aktinem a fodrinem[11])[9]
- Změny v struktuře membrány, např. vysunutí fosfatidylserinu na její vnější povrch („eat-me“ signál)[9]
- Poruchy propustnosti mitochondriální membrány a uvolňování cytochromu c[9]
- Pyknóza chromatinu (kondenzace do kompaktních shluků),[12][13] načež proběhne karyorrhexe (tzv. internukleozomální štěpení DNA do relativně malých úseků)[14] a následný rozpad jádra (karyolýza)[9]
- Rozpad buňky na apoptotická tělíska[9]
Funkce
Udržení rovnováhy
U dospělce je obvykle množství buněk v průběhu času relativně neměnné, a právě rovnováhu mezi buněčnou smrtí a buněčným dělením zajišťuje apoptóza. Když se buňky opotřebují či přestanou správně pracovat, jsou díky proliferaci okolní tkáně nahrazeny novými.[15] Tato rovnováha je jedním z principů udržení vnitřní homeostázy, díky níž udržuje každý živý organismus své vnitřní prostředí v určitých limitech. Pokud je narušena, mohou se objevit dva základní druhy problémů:
Usmrcení poškozených buněk
Apoptóza se jako proces uplatňuje například v likvidaci poškozených buněk, buněk napadených virem nebo například nedostatečně zásobených živinami. Ionizující záření, které způsobí poškození DNA, může rovněž nastartovat proces apoptózy, stejně jako některé jedovaté látky. Obvykle se o to v těchto dvou případech stará protein p53, který tím zamezí jakémukoliv nádorovému bujení poškozené buňky. U téměř poloviny všech lidských nádorů je gen pro tento protein zmutovaný.[17]
Usmrcení poškozených buněk jsou schopné docílit mimo jiné cytotoxické T-lymfocyty (typ bílých krvinek), které v cílové buňce vytvoří pomocí perforinu otvory a pomocí granzymů a různých proteáz nastartují v dané buňce apoptózu.[18]
Role ve vývinu a v dalších procesech
Apoptóza se uplatňuje i ve vývinu mnohobuněčných organizmů, zejména ve formaci určitých složitějších struktur, jichž by se jinak špatně docílilo prostým buněčným dělením v určitém směru. Je to ale v menší míře, než se dosud uvažovalo.[19] Díky skutečnosti, že při apoptóze dochází ke smrštění a fragmentaci buňky, může být tato velmi snadno fagocytována a jejich stavební látky mohou být tělem opět využity. Výzkum zejména na kuřecích zárodcích ukázal, jak cílená apoptóza určitých buněk společně s cíleným množením buněk vytváří embryonální tkáně živočichů. V průběhu této embryogeneze se z určitých částí embrya (zejména ze struny hřbetní) vylučuje signální molekula Shh („Sonic hedgehog“). Buňky, na jejichž receptor Patched1 se tato signální molekula naváže, pokračují v růstu a množení. Naopak na receptory buněk, na něž se žádné Shh nenavázalo, se místo toho naváže kaspáza 3 a vyvolá ve výsledku celý proces apoptózy, díky němuž tato buňka zahyne.[20][21]
Přebytečné mozkové buňky obratlovců (neurony) v průběhu embryogeneze spáchají apoptózu, pokud se na ně nenaváže dostatečné množství molekul NGF (nerve growth factor). Odstraňování ocasu pulců při jejich přeměně na dospělou žábu je zajištěno vylučováním hormonů ze štítné žlázy.[1] Apoptóza se v masovém měřítku uplatňuje i ve správném vývoji T-lymfocytů v brzlíku a B-lymfocytů v kostní dřeni. Na počátku tohoto procesu se vytvoří velké množství buněk s různorodými vazebnými místy pro různé nebezpečné antigeny, ale vznikají rovněž buňky, které jsou neúčinné nebo dokonce potenciálně nebezpečné pro tělo.[22] Také zmenšování prsou po odstavení kojence, zánik děložní sliznice při menstruaci, umírání pokožkových buněk, které je následováno jejich nahrazením buňkami novými, dobrovolná smrt neutrofilů během imunitní reakce, všechny tyto procesy existují díky apoptóze.[1]
Někdy se ve vývinu odlišných skupin organismů uplatňují různé jiné signální molekuly a faktory. Například pro tvorbu prstů u ptáků jsou zásadní BMP proteiny („Bone morphogenetic proteins“).[23] U octomilek (Drosophila) se na apoptóze významně podílí steroidní hormony. Vliv mohou mít i různé vývojové rozdíly mezi jedinci, jako je pohlaví. Díky němu páchají u samečků háďátek Caenorhabditis elegans buněčnou smrt určité neurony, konkrétně díky nízké aktivitě transkripčního faktoru TRA-1.[24]
Průběh apoptózy
Celý proces apoptózy buňky je spuštěn složitou soustavou různých látek. Tyto látky mohou být jak mimobuněčného původu, tak vnitrobuněčné, tedy produkované uvnitř buňky samotné. Z mimobuněčných molekul může apoptózu spustit vystavení buňky toxinům,[25] hormonům, růstovým faktorům, oxidu dusnatému[26] či cytokinům. Tyto látky buď proniknou přímo do buňky, nebo spustí biochemickou kaskádu z vnější části cytoplazmatické membrány. Jmenované látky mohou jak spouštět, tak zastavovat apoptózu v cílové buňce (tzv. negativní a pozitivní účinek). Vnitrobuněčná cesta se spouští převážně ve chvíli, kdy je buňka v jakékoliv formě stresu, jako je vystavení glukokortikoidům, teplu, radiaci, nedostatek živin, virová infekce buňky, nedostatek kyslíku[18] či také zvýšený obsah vápenatých iontů uvnitř buňky.[27] Všechny tyto jmenované signály jsou na počátku enzymatické kaskády, která je zodpovědná za provedení vlastní apoptózy.
Vnitřní cesta aktivace
Některé skutečnosti, jako například poškozená DNA, jedy přítomně v buňce nebo oxidativní stres,[28] mohou vyvolat vnitřní cestu aktivace apoptózy, která souvisí s mitochondriemi. Poškozením těchto organel se buňka vlastně vzdává poslední myšlenky na záchranu, protože mitochondrie představují nezbytné biochemické mašinérie, kde probíhá buněčné dýchání. Různé apoptotické proteiny (nebo možná i jiné látky, jako oxid dusnatý[26]) se mohou na základě výše zmíněných situací uvnitř buňky navázat na mitochondrii a způsobit vznik pórů v její membráně nebo zvýšení její propustnosti, čímž dochází k vylití různých látek do cytosolu. Tyto látky mnohdy představují další články v apoptotické kaskádě.[18] Patří k nim tzv. SMACs („Second mitochondria-derived activator of caspases“). Tyto se ihned navážou na další proteiny, tzv. IAPs (inhibitory apoptotických proteinů), deaktivují je a IAPs tím ztrácí svou schopnost bránit dalšímu pokračování apoptotické kaskády. Následně je aktivována skupina cystein proteáz, tzv. kaspázy, které způsobí vlastní degradaci buňky.[29]
Z mitochondrií se však nevylučují jen SMACs, ale protože se vytvoří ze speciálních molekul MAC v membráně mitochondrií určité kanály, může skrz membránu ven uniknout i cytochrom c,[18][30] protein, který se normálně podílí na transportu elektronů při buněčném dýchání. V tomto případě však funguje jako další z řady aktivátorů, protože se váže na protein Apaf-1 a za pomoci ATP také na pro-kaspázu 9. Z jmenovaných proteinů vzniká velký komplex zvaný apoptozom, který je schopný měnit pro-kaspázu 9 na kaspázu 3.
Pokud se ještě vrátíme k proteinovému kanálu MAC, je o něm známo, že je ovlivňován množstvím proteinů, jako je savčí Bcl-2 (bránící apoptóze), případně proteiny ced-9 u háďátka C. elegans.[31][32] Bcl-2 proteiny jsou schopné regulovat apoptózu buď právě pomocí kanálu MAC, nebo nepřímo pomocí jiných proteinů, případně dokonce i poté, co cytochrom c už unikl z mitochondrie.[18]
Vnější cesta aktivace
Vnější cesta aktivace apoptózy začíná na cytoplazmatické membráně, konkrétně na několika receptorech, zvaných „receptory smrti“.[28] Na tyto receptory se vážou různé ligandy, jako je TNF („Tumor necrosis factor“) či Fas ligand. Receptory jsou v podstatě oba ze skupiny TNF receptorů (TNFR).[33]
TNF je cytokin produkovaný zejména aktivovanými makrofágy. Obvykle bývají TNF receptory, přítomné na buňce, dvojího typu: TNF-R1 a TNF-R2. Vazba TNF na jeden z těchto receptorů má za následek aktivaci kaspáz, a to přes určité prostředníky (TRADD, FADD, TRAF2).[34][35] Fas receptor (také Apo-1 či CD95) se po navázání na Fas ligand[33] změní na tzv. DISC („death-inducing signaling complex“), který aktivuje kaspázu 8 a kaspázu 10. Kaspáza 8 pak obvykle aktivuje další kaspázy, které nevratně směřují k smrti buňky.[36] Fas ligandy se uplatňují ve velké roli v imunitním systému, protože se nachází na povrchu cytotoxických T-lymfocytů a mohou tak indukovat apoptózu u vybraných buněk. Mimoto mají tyto T-lymfocyty rovněž Fas receptor, který se může navázat na Fas ligand na té samé buňce a způsobit tak řízenou apoptózu opotřebovaných T-lymfocytů. Nádorové buňky toho někdy využívají, jelikož mohou obsahovat na svém povrchu Fas ligand, a tak mohou indukovat buněčnou smrt u T-lymfocytů, jejich „protivníků“.[28] Zvláštní způsob apoptózy, naprosto nezávislý na kaspázách, řídí molekula AIF (apoptosis-inducing factor).[37]
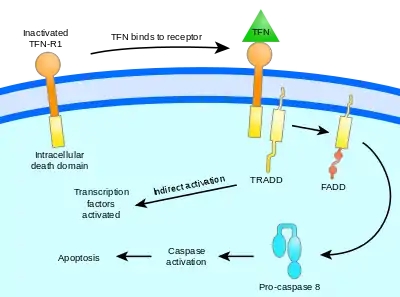 Znázornění cesty signálu přes TNF receptor |
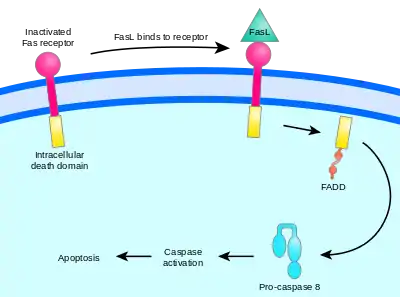 Znázornění cesty signálu přes Fas receptor |
Závěrečná fáze
Přestože, jak bylo uvedeno výše, cest vedoucích k apoptóze je více, existuje jeden hlavní mechanismus, jehož úkolem je provedení apoptózy jako takové. Po přijetí signálu totiž jsou aktivovány proteolytické enzymy, tzv. kaspázy. Kaspázy jsou cysteinové proteázy (enzymy rozkládající proteiny), které štěpí na karboxyskupině kyseliny asparagové.[9] Tzv. iniciační kaspázy katalyzují štěpení dalších druhů kaspáz a na konci tohoto řetězce vznikají tzv. exekutorové kaspázy. Exekutorové kaspázy se vážou na tzv. substráty smrti, mezi něž patří proteiny cytoskeletu, proteiny řídící opravu DNA, proteiny zabraňující apoptóze a podobně. V konečném důsledku vzniká z buňky několik váčků, tzv. apoptotická tělíska.[28]
Umírající buňky, které podstoupily závěrečnou fázi apoptózy a změnily se na apoptotická tělíska, vystavují na svém povrchu různé molekuly vyzývající k jejich pozření (fagocytóze). Mezi tyto signály patří například fosfatidylserin.[38] Tato molekula je v normální buňce pouze na vnitřní straně membrány, ale během apoptózy se díky proteinu skrambláze přemisťuje na vnější stranu.[39] Fosfatidylserin a jiné molekuly označují buňku, aby mohla být pohlcena, a to buď okolními buňkami, nebo buňkami imunitního systému[28] (jako zejména makrofágy).[40]
Onemocnění související s apoptózou
Nadměrná apoptóza má relevanci k onemocněním, jako je AIDS, aplastická anémie, různé degenerativní neurologické poruchy (Alzheimerova choroba, Huntingtonova choroba, Parkinsonova choroba), diabetes mellitus typu I, Hashimotova struma, chronická neutropenie, ischemie, lupus erythematodes, myelodysplastický syndrom, roztroušená skleróza, některá selhání jater, spinální svalová atrofie, ulcerózní kolitis, široké spektrum různých vývojových vad či Wilsonova choroba.[1]
Nedostatečná apoptóza v podstatě znamená, že může dojít ke vzniku nádorů nebo autoimunitních chorob (revmatoidní artritida)[41] Je klíčovým faktorem například u těchto onemocnění: autoimunitní lymfoproliferativní syndrom (Canale-Smithův syndrom). diabetes mellitus typu I. Gravesova choroba, hypereosinofilní syndrom, Hashimotova struma, leukémie, lupus erythematodes, různé lymfomy, osteoporóza, některé solidní nádory a neoplasie a také vývojové vady.[1]
Metody detekce apoptózy
- Nejspolehlivější metodou detekce apoptózy je pozorování fragmentace DNA pomocí elektronového mikroskopu.
- TUNEL (TdT-mediated dUTP-biotin nick end labelling, TUNEL assay) - enzymem terminální deoxynukleotidyl-transferasa (TdT) se naváže na 3'OH konce DNA označený dUTP. (Biotinylovaný dUTP se detekuje pomocí streptavidin-peroxidasy a zvizualizuje pomocí DAB. Když je dUTP označený fluoresceinem, detekuje se pomocí konjugátu alkalické fosfatázy a antifluoresceionové protilátky a zvizualizuje pomocí NBT/BCIP.) Metoda značí buňky na začátku a ve střední fázi apoptózy. Detekuje ale také nekrotické buňky. Používá se na tkáňové řezy.
- ISNTA (in situ nick translation assay) - syntetizuje DNA v místech zlomů pomocí DNA-polymerasy. Takto syntetizované úseky se označí pomocí biotinylovaného dUTP. Metoda označuje buňky v pozdní fázi apoptózy a čím pozdější fáze (čím více zlomů), tím je buňka označena intenzivněji. Používá se na tkáňové řezy i na buněčné kultury.
- Detekce kaspázy.
Seznamy molekul zahrnutých v apoptóze
Proapoptotické faktory
charakteristické rysy indukující apoptózu:
vnější vlivy schopné způsobit (vyvolávající) apoptózu:
- stres:[42] tepelný šok (hypertermie),[42] nefyziologická vlhkost, hypoxie
- poškození DNA a buňky[42]
- volné radikály
- chemoterapie[42]
- toxiny a toxické látky, např. H2O2, azid sodný
- NO
- hormony (steroidy)
- cytostatika (transformující růstové faktory – TNFα, TGF beta)
- (Fas-L, Reaper)[1]
- ozáření (radiace), např. rentgenové záření, ionizující záření[42]
- virová infekce, bakteriální infekce[42]
- interakce buněk
- růstové faktory (hormony a další regulační faktory), apoptózu indukuje jejich nadbytek i nedostatek[42]
- glukokortikoidy[42]
- cytokiny (cytokiny aktivují sfingomyelinový cyklus, produktem je ceramid, který blokuje buněčný cyklus a podporuje apoptózu)[42]
vnitřní faktory podporující apoptózu
- Ca2+[43]
- specifické molekuly, které se vážou na určité receptory
- TNF receptor
- např. cytokin (v tomto případě obecně FAS-ligand (FASL)) se naváže na trimer FAS receptoru. Trimer FAS se spojí s FADD (= Fas-associated death domain), FADD způsobí změnu procaspázy na caspázu.
Antiapoptotické faktory
- uplatňují se geny genové rodiny bcl-2, podrodina Bcl-2:
- onkogen bcl-2 (= B cell lymphomal leukemia-2) - produkuje protein BCL-2, který ve vnitřní membráně mitochondrií pravděpodobně omezuje vliv volných radikálů (detoxikuje volné radikály). V endoplazmatickém retikulu omezuje uvolňování Ca2+.
- onkogen bcl-xL[1]
- protein Bcl-w
- gen Mcl-1 (= myeloid cell leukemia-1)[1]
- gen A 1/bfl-1[1]
- NR-13
- BHRF1 (virus Epsteina a Barrové)
- LMW5-HL (african swine fewer virus)
- ORF16
- KS-bcl-2
- adenovirus protein E1B19kD[46]
- ced-9 (hlístice Caenorhabditis elegans)
- gen Rb (= retinoblastoma gene product)[1]
- gen brag-1[1]
- gen boo
- protein Sonic hedgehog (SSH) - význam ve vývoji struny hřbetní (chorda dorsalis)
Eat-me molekuly na apoptotické buňce
- Fosfatidylserin se z vnitřní strany cytoplazmatické membrány přesmykne na vnější stranu a na něj se vážou některé z těchto proteinů.
- ABC1 = ATP binding-cassette-transporter 1 ze skupiny ABC transportérů
- AnxI = Annexin I
- β2-GPI = β2-glycoprotein-I
- C1q = complement protein C1q
- C3b/bi = complement protein C3b/bi
- Gas6 = growth arrest-specific 6
- ICAM-3 = intercellular adhesion molecule 3
- MBL = mannose binding lectin
- MFG-E8 = milk-fat-globule-EGF-factor 8
- OxLDL = oxidized low-density lipoprotein particle
- SP-A/D = lung surfactant protein A or D
- TSP-1 = thrombospondin-1
- vitronektin (vitronectin, S-protein, protein S, zkratka: Prot S)[47]
Receptory na makrofágu:[48]
- CD14 = lipopolysaccharide receptor CD14
- CD91 = calreticulin/heat shock protein receptor; (CRT = calreticulin)
- CR3 = complement receptor 3
- CR4 = complement receptor 4
- LOX1 = lectin-like oxidized low-density lipoprotein particle receptor 1
- Mer = receptor-tyrosin-kinase (exprimuje se v monocytech a ve tkáních epiteliálního a rozmnožovacího původu)
- SR-A = class A macrophage scavenger receptor
- předpokládal se význam PS-receptor = phosphatidylserine receptor,[49] ale normální apoptóza funguje i bez PS-receptoru[50]
Odkazy
Reference
V tomto článku byl použit překlad textu z článku apoptosis na anglické Wikipedii.
- SAIKUMAR, P, Z Dong, V Mikhailov, M Denton, J M Weinberg, M A Venkatachalam. Apoptosis: definition, mechanisms, and relevance to disease. The American Journal of Medicine. 1999-11, roč. 107, čís. 5, s. 489–506. Dostupné online. ISSN 0002-9343. DOI 10569305.
- NOODÉN, Larry D. Cell Death in Plants. [s.l.]: Academic Press, 2000. ISBN 0125209150, ISBN 9780125209151.
- KERR, J F, A H Wyllie, A R Currie. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. British Journal of Cancer. 1972-08, roč. 26, čís. 4, s. 239–57. Dostupné online. ISSN 0007-0920. DOI 4561027.
- Webster.com dictionary entry. www.webster.com [online]. [cit. 2008-12-18]. Dostupné v archivu pořízeném dne 2007-07-03.
- FADEEL, B, S Orrenius. Apoptosis: a basic biological phenomenon with wide-ranging implications in human disease. Journal of Internal Medicine. 2005-12, roč. 258, čís. 6, s. 479–517. Dostupné online. ISSN 0954-6820. DOI JIM1570.
- NARULA, J, S Kharbanda, M Sluyser. Apoptosis in the eyes of a clinician. Apoptosis: An International Journal on Programmed Cell Death. Roč. 6, čís. 1–2, s. 5. Dostupné online [cit. 2008-12-14]. ISSN 1360-8185. DOI 11321041.
- Apoptosis and Cell Proliferation
- Nobelova cena za medicínu 2002 (anglicky)
- Programovaná buněčná smrt [online]. Masarykova univerzita. Dostupné online.
- Blebbing - Definiton [online]. Merriam Webster Dictionary. Dostupné online.
- GLENNEY, John R.; GLENNEY, Phyllis. Fodrin is the general spectrin-like protein found in most cells whereas spectrin and the TW protein have a restricted distribution. Cell. Roč. 34, čís. 2, s. 503–512. DOI 10.1016/0092-8674(83)90383-5.
- Santos A. Susin, et al.. Two Distinct Pathways Leading to Nuclear Apoptosis. Journal of Experimental Medicine. 2000, roč. 192, čís. 4, s. 571–80. Dostupné online. DOI 10.1073/pnas.191208598v1. PMID 10952727.
- Madeleine Kihlmark, et al.. Sequential degradation of proteins from the nuclear envelope during apoptosis. Journal of Cell Science. 2001, čís. 114, s. 3643–53. Dostupné online. PMID 11707516.
- Nagata S. Apoptotic DNA fragmentation. Exp. Cell Res.. April 2000, roč. 256, čís. 1, s. 12–8. DOI 10.1006/excr.2000.4834. PMID 10739646.
- Thompson, CB. Apoptosis in the pathogenesis and treatment of disease. Science. 1995, roč. 267, čís. 5203, s. 1456–62. Dostupné online. DOI 10.1126/science.7878464. PMID 7878464.
- FRIESS, H, Z Lu, H U Graber, A Zimmermann, G Adler, M Korc, R M Schmid, M W Büchler. bax, but not bcl-2, influences the prognosis of human pancreatic cancer. Gut. 1998-09, roč. 43, čís. 3, s. 414–21. Dostupné online [cit. 2008-12-18]. ISSN 0017-5749.
- Lennarz,W.J., Lane, M.D. ENCYCLOPEDIA OF BIOLOGICAL CHEMISTRY, FOUR-VOLUME SET, 1-4. [s.l.]: [s.n.]
- COTRAN, Kumar, Collins. Robbins Pathologic Basis of Disease. Philadelphia: W.B Saunders Company, 1989. Dostupné online. ISBN 0-7216-7335-X.
- PAZDERA, Josef. Podraz na embryo průhlednou „dělohou“. osel.cz [online]. 2014-02-17 [cit. 2021-05-16]. Dostupné online.
- Guerrero I, Ruiz i Altaba A. Development. Longing for ligand: hedgehog, patched, and cell death. Science. August 2003, roč. 301, čís. 5634, s. 774–6. DOI 10.1126/science.1088625. PMID 12907783.
- Thibert C, Teillet MA, Lapointe F, Mazelin L, Le Douarin NM, Mehlen P. Inhibition of neuroepithelial patched-induced apoptosis by sonic hedgehog. Science. August 2003, roč. 301, čís. 5634, s. 843–6. DOI 10.1126/science.1085405. PMID 12907805.
- Werlen G, et al.. Signaling life and death in the thymus: timing is everything. Science. 2003, roč. 299, čís. 5614, s. 1859–63. DOI 10.1126/science.1067833. PMID 12649474.
- PAJNI-UNDERWOOD, Sangeeta, Catherine P. Wilson, Cindy Elder, Yuji Mishina, Mark Lewandoski. BMP signals control limb bud interdigital programmed cell death by regulating FGF signaling. Development. 2007-06-15, roč. 134, čís. 12, s. 2359–2368. Dostupné online [cit. 2008-12-18]. DOI 10.1242/dev.001677.
- PORTMAN, Douglas S. Genetic control of sex differences in C. elegans neurobiology and behavior. Advances in Genetics. 2007, roč. 59, s. 1–37. Dostupné online [cit. 2008-12-18]. ISSN 0065-2660. DOI 10.1016/S0065-2660(07)59001-2.
- Popov SG, Villasmil R, Bernardi J, et al. Lethal toxin of Bacillus anthracis causes apoptosis of macrophages. Biochem. Biophys. Res. Commun.. April 2002, roč. 293, čís. 1, s. 349–55. DOI 10.1016/S0006-291X(02)00227-9. PMID 12054607.
- Brüne B. Nitric oxide: NO apoptosis or turning it ON?. Cell Death Differ.. August 2003, roč. 10, čís. 8, s. 864–9. DOI 10.1038/sj.cdd.4401261. PMID 12867993.
- Calcium orchestrates apoptosis. Mark P. Mattson & Sic L. Chan. Nature Cell Biology 5, 1041 - 1043 (2003). doi:10.1038/ncb1203-1041
- BALÁŽ, Vojtech, et al. Smrt jako součást života. Praha: NIDM, 2008. Dostupné v archivu pořízeném dne 2013-01-14. Archivováno 14. 1. 2013 na Wayback Machine
- Fesik SW, Shi Y. Controlling the caspases. Science. 2001, roč. 294, čís. 5546, s. 1477–8. DOI 10.1126/science.1062236. PMID 11711663.
- Laurent M. Dejean, Sonia Martinez-Caballero, Kathleen W. Kinnally. Is MAC the knife that cuts cytochrome c from mitochondria during apoptosis?. Cell Death and Differentiation. 2006, roč. 13, s. 1387–5. Dostupné online. DOI 10.1038/sj.cdd.4401949.
- Dejean LM, Martinez-Caballero S, Manon S, Kinnally KW. Regulation of the mitochondrial apoptosis-induced channel, MAC, by BCL-2 family proteins. Biochim. Biophys. Acta. February 2006, roč. 1762, čís. 2, s. 191–201. DOI 10.1016/j.bbadis.2005.07.002. PMID 16055309.
- LODISH, Harvey, et al.. Molecular Cell Biology. New York: W.H. Freedman and Company, 2004. Dostupné online. ISBN 0-7167-4366-3.
- Wajant H. The Fas signaling pathway: more than a paradigm. Science. 2002, roč. 296, čís. 5573, s. 1635–6. DOI 10.1126/science.1071553. PMID 12040174.
- Chen G, Goeddel DV. TNF-R1 signaling: a beautiful pathway. Science. 2002, roč. 296, čís. 5573, s. 1634–5. DOI 10.1126/science.1071924. PMID 12040173.
- Goeddel, DV et al. Connection Map for Tumor Necrosis Factor Pathway. Science. Dostupné v archivu pořízeném dne 2009-07-10. DOI 10.1126/stke.3822007tw132].
- Wajant, H. Connection Map for Fas Signaling Pathway. Science. Dostupné v archivu pořízeném dne 2009-05-03. DOI 10.1126/stke.3802007tr1.
- Susin SA, Lorenzo HK, Zamzami N, et al. Molecular characterization of mitochondrial apoptosis-inducing factor. Nature. February 1999, roč. 397, čís. 6718, s. 441–6. DOI 10.1038/17135. PMID 9989411.
- Li MO, et al.'. Phosphatidylserine receptor is required for clearance of apoptotic cells. Science. 2003, roč. 302, čís. 5650. DOI 10.1126/science.1087621. PMID 14645847.
- Wang X, et al.. Cell corpse engulfment mediated by C. elegans phosphatidylserine receptor through CED-5 and CED-12. Science. 2003, roč. 302, čís. 5650, s. 1563–1566. DOI 10.1126/science.1087641. PMID 14645848.
- Savill J, Gregory C, Haslett C. Eat me or die. Science. 2003, roč. 302, čís. 5650. DOI 10.1126/science.1092533. PMID 14645835.
- Apoptosis and Cell Proliferation
- Chaloupka J. (1996) Programovaná smrt buňky. - Biologické listy, 61(3-4): 2479-271.
- Kohlíková E. et al. (2003) Cytopatologie, patobiochemie a patofyziologie. - Praha, (strany 31-33).
- říjen 2005 apoptóza na stránkách firmy Roche - v levém menu např.: signální dráhy.
- Adams, J. M. & Cory, S. (1998) The Bcl-2 protein family: arbiters of cell survival. Science, 281: 1322-1326.
- Huang, D.C.S., Cory, S., Strasser, A.: Bcl-2, Bcl-xL and adenovirus protein E1B19kD are functionally equivalent in their ability to inhibit cell death. (1997) Oncogene, 14 pp. 405-414
- Archivovaná kopie. www.promega.com [online]. [cit. 2006-01-18]. Dostupné v archivu pořízeném z originálu dne 2005-08-29.
- Lauber, K., Blumenthal, S.G., Waibel, M., Wesselborg, S. (2004) Clearance of apoptotic cells: Getting rid of the corpses. - Molecular Cell, 14(3): 277-287.
- SAVILL, J. CELL BIOLOGY: Eat Me or Die. S. 1516–1517. Science [online]. 2003-11-28. Roč. 302, čís. 5650, s. 1516–1517. Dostupné online. DOI 10.1126/science.1092533. PMID 14645835. (anglicky)
- BÖSE, Jens; GRUBER, Achim D; HELMING, Laura; SCHIEBE, Stefanie; WEGENER, Ivonne; HAFNER, Martin; BEALES, Marianne. [No title found]. S. 15. Journal of Biology [online]. 2004. Roč. 3, čís. 4, s. 15. Dostupné online. DOI 10.1186/jbiol10. PMID 15345036.
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu apoptóza na Wikimedia Commons
Obrázky, zvuky či videa k tématu apoptóza na Wikimedia Commons - (česky) Programovaná buněčná smrt na Masarykově univerzitě - Pdf soubor
- Kušíková E.: Co je to apoptóza a proč nás zajímá?
- Topinková E.: Apoptóza - programovaná buněčná smrt a stárnutí. - Sanquis, 2002/20.
- Apoptóza - role kaspázy 3
- Apoptóza - role kaspázy 8
- Apoptóza - role kaspázy 7
- Biochemweb.org - rozcestník stránek o této tematice
- Apoptosis Info - Informace o průběhu apoptózy, odkazy, fóra, novinky
- Databáze proteinů zahrnutých v apoptóze
- Video apoptózy
- Přehledné vysvětlení molekulárního průběhu apoptózy
- Wikipathways - kompletní přehled známých biochemických cest při apoptóze u člověka ve formě obrázku
- signální dráhy tematicky roztříděné na jednotlivých slidech
- Finding Cancer’s Self-Destruct Button[nedostupný zdroj] CR magazine (Spring 2007). Article on apoptosis and cancer.