Cytoplazmatická membrána
Cytoplazmatická membrána (také plazmatická membrána, plazmalema) je tenký semipermeabilní obal ohraničující vnitřek buňky od vnějšího prostředí a kontrolující pohyb látek do buňky a ven z buňky.[1][2] Buněčná membrána je selektivně permeabilní pro ionty a organické molekuly, a v zásadě chrání buňku před vnějšími vlivy. Skládá se z lipidové dvouvrstvy s vmezeřenými molekulami bílkovin. Buněčná membrána se podílí na různých buněčných procesech jako buněčná adheze, výměna iontů a buněčná signalizace, také slouží jako cílová struktura pro několik extrabuněčných struktur, včetně buněčné stěny, glykokalyxu a vnitřního buněčného cytoskeletu. Cytoplazmatická membrána může být uměle vytvořena.[3][4][5]
Funkce
Buněčná membrána obklopuje cytoplazmu buňky, u živočišných buněk, a fyzicky odděluje intracelulární komponenty od extracelulárního prostředí. Houby, bakterie a rostliny také mají buněčnou stěnu, která uskutečňuje mechanickou oporu pro buňku a zabraňuje průniku velkých molekul. Buněčná membrána hraje roli při ukotvení cytoskeletu, dává buňce tvar, má vliv na uchycení k extracelulární matrix a k jiným buňkám, pomáhá skupinám buněk formovat se společně do tkání.
Membrána je selektivně permeabilní a může regulovat, které látky vstoupí do buňky a nebo naopak uniknou ven z buňky. Tím usnadňuje transport materiálů potřebných pro přežívání. Průnik látek přes membránu může být pasivní, uskutečňující se bez dodání energie, nebo aktivní, vyžadující pro transport látky dodání (vynaložení) energie. Membrána také udržuje buněčný potenciál. Buněčná membrána pracuje jako selektivní filtr tím, že dovolí jen určitým látkám vstoupit dovnitř nebo opustit buňku. Buňka využívá počet transportních mechanismů, které zahrnuje biologická membrána:
- Pasivní difúze a osmóza: některé látky (malé molekuly, ionty) tak jako oxid uhličitý, kyslík a voda mohou pronikat skrz plazmatickou membránu difúzí, která patří mezi pasivní transportní procesy. Membrána působí jako bariéra pro určité molekuly a ionty, které se mohou vyskytovat v odlišných koncentracích na obou stranách membrány. Koncentrační gradient přes semipermeabilní membránu umožňuje osmotický tok vody skrz membránu.
- Transmembránové proteinové kanály a transportéry: živiny, jako cukry nebo aminokyseliny, musí vstoupit do buňky a určité produkty metabolismu musí opustit buňku. Takové molekuly jsou pumpovány skrz membránu transmembránovým transportem nebo difúzí proteinovými kanály. Tyto proteiny, také zvané permeasy jsou obvykle docela specifické, rozpoznávají a transportují pouze limitované skupiny chemických látek, často pouze jedinou substanci.
- Endocytóza: je proces, kdy buňka absorbuje molekuly jejich pohlcením. Plazmatická membrána utvoří malé deformace směřující dovnitř, zvaná invaginace, ve kterých je látka určená k transportu pohlcena. Deformace pak zaštípnutím membrány dovnitř buňky utvoří váček obsahující pohlcenou látku. Endocytóza je cesta pro pohlcování pevných částic (buněčné jezení nebo fagocytoza), malých molekul a iontů (buněčné pití nebo pinocytoza), a makromolekul. Endocytóza vyžaduje dodání energie a tím se řadí do aktivního transportu.
- Exocytóza: materiál může být dopraven do buňky invaginací a formováním váčku, membrána váčku může fúzovat s plazmatickou membránou a vypudit obsah váčku do okolního media. Tento proces se nazývá exocytoza. Exocytozu využívají různé buňky k odstranění zbytků látek získaných endocytozou, k sekreci látek jako hormony a enzymy, k transportu látek skrz celulární barieru. Při procesu exocytozy vakuoly obsahující odpad z potravy nebo sekreční vakuoly vypučené z Golgiho aparátu se nejdříve přemístí po cytoskeletu z vnitřní části buňky k povrchu. Membrána váčku se dostane do kontaktu s plazmatickou membránou. Lipidové molekuly z lipidové dvouvrstvy se přesunou, přeskupí se a dvě membrány navzájem fúzují. Přechod skrz membránu je uskutečněn fúzováním membrány s váčkem a vyprázdněním obsahu váčku ven z buňky.
Bakterie
Gramnegativní bakterie mají plazmatickou membránu a vnější membránu oddělené periplazmatickým prostorem. Jiné prokaryotické organizmy mají jen plazmatickou membránu. Prokaryotické buňky jsou také obklopeny buněčnou stěnou složenou z peptidoglykanu (aminokyseliny a cukry). Některé eukaryotické buňky také mají buněčné stěny, ale ty neobsahují peptidoglykan.
Struktura
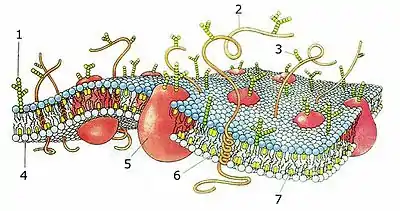
1. glykolipid
2. alfa-helix protein
3. oligosacharidový boční řetězec
4. fosfolipid
5. globulární protein
6. hydrofobní část alfa-helix proteinu
7. cholesterol
Model fluidní mozaiky
Podle modelu fluidní mozaiky S. J. Singera a G. L. Nicolsona mohou být biologické membrány považovány za dvoudimenzionální tekutinu, ve které všechny molekuly lipidů a proteinů více či méně snadno difundují.[6] Ačkoliv trojrozměrná je již samotná lipidová dvouvrstva, k 3D uspořádání přispívá dále velké množství proteinů. Tyto proteiny umožňují vznik komplexů protein-protein, pickets and fences formace vycházející z aktinového cytoskeletu, ale také lipidové rafty.
Lipidová dvouvrstva
Lipidová dvouvrstva se formuje procesem self-assembly, což znamená, že se formuje spontánně. Buněčná membrána primárně obsahuje tenkou vrstvu amfipatických fosfolipidů, které se spontánně sestavují do tzv. hydrofobního ocasu, který je z obou stran izolován polárními tekutinami. To má za následek, že se oblast více hydrofilní hlavičky asociuje s intracelulárním a extracelulárním do tvaru výsledné spojité lipidové dvouvrstvy. Na formaci lipidové dvouvrstvy se podílejí různé síly jako van der Waalsovy, elektrostatické, hydrogenní a nekovalentní interakce. Přesto jsou hlavní silou formující lipidovou dvojvrstvu hydrofobní interakce.
Lipidové dvojvrstvy jsou v zásadě nepropustné pro polární molekuly. Seskupení do hydrofilní hlavičky a hydrofobní ho ocasu lipidové dvouvrstvy brání polárním rozpuštěným látkám (aminokyseliny, nukleové kyseliny, cukry, proteiny, ionty) v difúzi skrz membránu, ale všeobecně připouští pasivní difúzi hydrofobních molekul. To poskytuje buňce schopnost kontrolovat pohyb těchto látek skrz transmembránové proteinové komplexy, jako jsou póry, kanály a brány. Flipázy a skramblázy soustředí fosfatidylserin, který nese negativní náboj, na vnitřní straně membrány. Společně s kyselinou sialovou (NANA) tvoří vnější bariéru pro látky s nabitými skupinami procházející skrz membránu.
Membrány zajišťují odlišnou funkci u eukaryotických a u prokaryotických buněk. Jednou z důležitých rolí je regulace prostupu materiálu do a z buňky. Struktura fosfolipidové dvouvrstvy (model fluidní mozaiky) se specifickými membránovými proteiny vysvětluje selektivní propustnost (permeabilitu) membrány a pasivní a aktivní transportní mechanismus. Navíc membrány prokaryot a v mitochondriích a chloroplastech eukaryot usnadňují syntézu ATP pomocí chemiosmózy.
Polarita membrány
Apikální membrána na polarizované buňce je na povrchu plazmatické membrány, která tvoří lumen. To je zvláště evidentní v epiteliálních a endoteliálních buňkách, ale také popisuje jiné polarizované buňky, jako jsou neurony.
Bazolaterální membrána polarizované buňky je částí plazmatické membrány, která vytváří její bazální a laterální povrchy. Nachází se směrem k intersticiu a pryč od lumen. Bazolaterální membrána se skládá z bazální membrány a laterální (postranné, boční) membrány, jejíchž aktivita a složení u epiteliálních buněk jsou identické. Proteiny (např., iontové kanály a pumpy) se volně pohybují z bazálního na laterální povrch a naopak v souladu s modelem fluidní mozaiky. Tight junction (tzv. těsný spoj) spojují epiteliální buňky těsně blízko jejich apikálních povrchů, a tím zabraňuje průchodu proteinů z bazolaterální membrány k apikální membráně. Bazální a laterální povrchy zůstávají zhruba ekvivalentní a rovnocenné k sobě navzájem a zároveň jasně odlišné od apikálního povrchu.
Nadmembránové struktury
Buněčná membrána může vytvářet různé typy „nadmembránových“ struktur jako kaveoly, postsynaptická těsnění (postsynaptic density), podozomy, invadopodia, fokální adheze a různé typy buněčných spojů. Tyto struktury jsou obvykle odpovědné za buněčnou adhezi, komunikaci, endocytózu a exocytózu. Mohou být zobrazeny pomocí elektronové mikroskopie a fluorescenčního mikroskopu. Jsou sestaveny ze speciálních proteinů – integrinů a kadherinů.
Cytoskelet
Cytoskelet vytváří podklad buněčné membrány v cytoplazmě a vytváří lešení pro membránové proteiny, stejně jako pro tvořící se organely, které vyčnívají z buňky. Prvky cytoskeletu interagují velmi intenzivně a úzce s buněčnou membránou.[7] Kotvící proteiny omezují cytoskelet jen na určitý povrch buněk – např. apikální povrch epiteliální buňky, který lemuje střevo obratlovců. Cytoskelet na povrchu buněk může formovat přívěškovité organely, jako jsou cílie (mikrotubulárního původu) a filopodia, která jsou z aktinového materiálu. Tato rozšíření jsou zakomponována v membráně a vyčnívají z buněčného povrchu za účelem vnímání vnějšího prostředí a zároveň umožňují kontakt se substráty nebo jinými buňkami. Apikální povrchy epiteliálních buněk jsou hustě pokryté aktinovými výběžky, tzv. mikroklky, které zvětšují buněčný povrch a tím zvyšují rychlost vstřebávání živin. Místní rozpojení cytoskeletu a buněčné membrány vytváří dále různé puchýřky (bubliny).
Složení
Buněčná membrána obsahuje různé biologické molekuly, především lipidy a proteiny. Materiál je zakomponován do membrány nebo z ní odstraněn různými mechanismy:
- Fúzí intracelulárních vezikulů s membránou (exocytóza) dochází nejen k exkreci obsahu vezikulů, ale také k zabudování vezikulárních membránových komponent do buněčné membrány. Membrána může formovat puchýřky okolo extracelulárního materiálu, který se uštípnutím stane vezikulem (endocytóza).
- Jestliže je membrána spojená s tubulárními strukturami vytvořenými z membránového materiálu, pak materiál z tubulů může být nepřetržitě vpravován do membrány.
- Ačkoli koncentrace membránových komponent ve vodné fázi je nízká (stabilní membránové komponenty mají nízkou rozpustnost ve vodě), dochází zde k výměně molekul mezi lipidy a vodnou fází.
Lipidy
Příklady hlavních membránových fosfolipidů a glykolipidů: fosfatidylcholin (PtdCho), fosfatidylethanolamin (PtdEtn), fosfatidylinositol (PtdIns ), fosfatidylserin (PtdIns). Cytoplazmatická membrána obsahuje tři třídy amfipatických lipidů: fosfolipidy, glykolipidy a cholesteroly. Množství jednotlivých typů lipidů závisí na druhu buňky, ale ve většině případů jsou nejhojněji zastoupeny fosfolipidy.[8] V červených krvinkách tvoří 30 % plazmatické membrány lipidy.
Mastné řetězce fosfolipidů a glykolipidů obvykle obsahují sudý počet atomů uhlíku, typicky mezi 16 a 20 atomy uhlíku. Ty 16 a 18 uhlíkaté mastné kyseliny jsou nejčastější. Mastné kyseliny jsou buď nasycené nebo nenasycené s konfigurací dvojné vazby téměř vždy „cis“. Délka a stupeň nenasycenosti řetězců mastných kyselin má nesmírný vliv na fluiditu membrány.[9] Nenasycené lipidy vytvářejí ohyby (klinks) a tak předcházejí společnému pevnému sbalení mastných kyselin, čímž se snižuje teplota tání (vzrůst fluidity) membrány. Schopnost organismu regulovat fluiditu buněčné membrány změnou složení lipidů se nazývá homeovisciósní adaptace.
Celá membrána drží pohromadě pomocí nekovalentní interakce hydrofobních konců, nicméně struktura je docela pohyblivá a není fixována pevně na místě. Za fyziologických podmínek jsou fosfolipidové molekuly v membráně tekutého krystalického stavu. To znamená, že lipidové molekuly volně difundují a vykazují rychlé boční šíření podél vrstvy, kde se vyskytují. Nicméně výměna fosfolipidových molekul mezi intracelulárním a extracelulárním listem dvouvrstvy je velmi zdlouhavý proces. Lipidové rafty a kaveoly jsou příkladem cholesterolem obohacených mikrodomén v buněčné membráně. V živočišných buňkách se cholesterol běžně vyskytuje rozptýlený v různé míře po celé membráně, v nepravidelných prostorech mezi hydrofobními konci membránových lipidů, kde vytváří ztužující a zesilující účinek na membránu [2]
Sacharidy
Plazmatická membrána také obsahuje karbohydráty, převážně glykoproteiny, ale s některými glykolipidy (cerebrosidy a gangliosidy). Z velké části, bez glykosylace na membráně z vnitřní části buňky; spíše obecně glykosylace probíhá na extracelulárním povrchu plazmatické membrány. Glykokalyx je důležitý prvek ve všech buňkách, zvláště epitelií s mikrovili. Nedávná data naznačují, že se glykokalyx podílí na buněčné adhezi, při homingu lymfocytů a mnoha jiných procesech. Předposlední cukr je galaktóza a poslední cukr je kyselina sialová, cukerná kostra se vytváří v golgiho aparátu. Kyselina sialová nese negativní náboj poskytuje vnější bariéru nabitých částic.
Proteiny
| Typ | Popis | Příklad |
|---|---|---|
| Integrální proteiny nebo transmembránové proteiny | Mají hydrofilní cytosolickou doménu,která interaguje s vnitřními molekulami,pak hydrofobní doménu roztahující membránu, která je ukotvuje uvnitř buněčné membrány, a hydrofilní extracelulární doménu, která interaguje s vnějšími molekulami. Hydrofobní domény se skládají z jedné, více nebo kombinace α-helix a β listů proteinových motivů | Iontové kanály, protonové pumpy, receptory spřažené s G proteiny |
| Proteiny ukotvené s lipidy | Jsou kovalentně vázané na jednu nebo více lipidových molekul, hydrofobně vložené do buněčné membrány. Proteiny samy o sobě nejsou v kontaktu s membránou. | G proteiny |
| Periferní proteiny | Spojené s integrálními membránovými proteiny nebo asociováné s periferními oblastmi lipidové dvouvrstvy. Tyto proteiny mají tendenci mít pouze dočasné interakce s biologickými membránami a jednou reagující molekula se odloučí a pokračuje ve své práci v cytoplazmě. | Některé enzymy a některé hormony. |
Buněčná membrána obsahuje velké množství proteinů, typicky kolem 50 % membránového objemu.[9] Tyto proteiny jsou pro buňku důležité, protože jsou odpovědné za různé biologické aktivity. Přibližně třetina genů kvasinky je specificky kóduje, a jejich počet je dokonce vyšší u vícebuněčných organismů.[8] Buněčná membrána je vystavena vnějšímu prostředí a je důležitým místem pro mezibuněčnou komunikaci. Existuje velké množství proteinových receptorů a identifikačních proteinů, jako jsou antigeny, které jsou přítomné na povrchu membrány. K funkcím membránových proteinů také patří buněčný kontakt, povrchové rozpoznání, kontakt cytoskeletu, signalizace, enzymatické aktivity nebo transport látek přes membránu. Nejvíce membránových proteinů musí být nějak vloženo do membrány. Proto se naskytuje N-koncová signální sekvence aminokyselin směřující proteiny do endoplazmatického retikula, kde se vkládají proteiny do lipidové dvouvrstvy. Jednou vložené proteiny jsou pak transportovány do jejich cílové destinace ve váčku, kde váček fúzuje s cílovou membránou.
Variace
Buněčná membrána má odlišné složení lipidů a proteinů v různých typech buněk a smí proto mít specifická jména v určitých buněčných typech.
Historicky byla plazmatická membrána také obecně označována jako plazmalema.
U sinic mořského planktonu v Sargasovém moři byly v r. 2009 popsány plazmatické membrány bez fosfolipidů – náhradním membránovým lipidem je tzv. SQDG (sulfoquinovosyldiacylglycerol). Na objevu se podíleli i vědci Akademie věd České republiky. Výskyt byl zjištěn i na jiných místech oceánů s nedostatkem fosforečných a dusíkatých živin.[10]
Propustnost
Propustnost membrány je míra pasivní difuze molekul membránou. Molekuly schopné difuzi dostaly název „permeant molecules“ (pronikavé molekuly). Propustnost závisí hlavně na elektrickém náboji a polaritě molekul a v menší míře na molární hmotnosti molekuly. Vzhledem k tomu, že membrána je hydrofilní povahy, malé elektricky neutrální molekuly projdou skrz membránu snáze než nabité, velké molekuly. Neschopnost nabitých molekul projít skrz buněčnou membránu závisí na pH rozdělení látek v celé tekutině části těla.
Odkazy
Reference
V tomto článku byl použit překlad textu z článku cell membrane na anglické Wikipedii.
- Kimball's Biology Pages Archivováno 25. 1. 2009 na Wayback Machine, Cell Membranes
- Alberts B, Johnson A, Lewis J, et al.. Molecular Biology of the Cell. 4th. vyd. New York: Garland Science, 2002. Dostupné online. ISBN 0-8153-3218-1. (anglicky)
- BUDIN, Itay; DEVARAJ, Neal K. Membrane Assembly Driven by a Biomimetic Coupling Reaction. Journal of the American Chemical Society. December 29, 2011, s. 751–753. Dostupné online [cit. February 18, 2012]. DOI 10.1021/ja2076873. (anglicky)
- Staff. Chemists Synthesize Artificial Cell Membrane [online]. ScienceDaily, January 25, 2012 [cit. 2012-02-18]. Dostupné online.
- Staff. Chemists create artificial cell membrane [online]. kurzweilai.net, January 26, 2012 [cit. 2012-02-18]. Dostupné online.
- Singer SJ, Nicolson GL. The fluid mosaic model of the structure of cell membranes. Science. 1972, s. 720–731. Dostupné online. DOI 10.1126/science.175.4023.720. PMID 4333397. (anglicky)
- Doherty GJ and McMahon HT. Mediation, Modulation and Consequences of Membrane-Cytoskeleton Interactions. Annual Review of Biophysics. 2008, s. 65–95. Dostupné online. DOI 10.1146/annurev.biophys.37.032807.125912. PMID 18573073. (anglicky)
- Lodish H, Berk A, Zipursky LS, et al.. Molecular Cell Biology. 4th. vyd. New York: Scientific American Books, 2004. Dostupné online. ISBN 0-7167-3136-3. (anglicky)
- Jesse Gray, Shana Groeschler, Tony Le, Zara Gonzalez. Membrane Structure [SWF]. Davidson College [cit. 2007-01-11]. Dostupné online.
- Phytoplankton cell membranes challenge fundamentals of biochemistry. phys.org [online]. 2009-02-02 [cit. 2020-12-13]. Dostupné online. (anglicky)
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu cytoplazmatická membrána na Wikimedia Commons
Obrázky, zvuky či videa k tématu cytoplazmatická membrána na Wikimedia Commons