Paranthropus boisei
Paranthropus boisei (česky též Australopiték východoafrický)[1] je druh vyhynulého hominida, žijící ve starším pleistocénu, před 2,3 – 1,3 miliony let ve východní Africe, na území dnešní Etiopie, Keni, Tanzanie a Malawi. Je nejmladším, posledním zástupcem australopitéků, přičemž patří ke skupině robustních druhů, řazených často do samostatného rodu Paranthropus. Žil již současně se zástupci rodu Homo (Homo habilis, Homo erectus).
Stratigrafický výskyt: pleistocén | |
|---|---|
 | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Třída | savci (Mammalia) |
| Řád | primáti (Primates) |
| Čeleď | Hominidé (Hominidae) |
| Podčeleď | Homininae |
| Tribus | Hominini |
| Rod | Paranthropus Broom, 1938 |
| Binomické jméno | |
| Paranthropus boisei Leakey, 1959 | |
| Některá data mohou pocházet z datové položky. | |
Objevy
Paranthropus boisei byl velmi úspěšným druhem hominidů, který obýval rozsáhlé prostory východní Afriky téměř po milion let. I díky tomu je doložen vcelku značným množstvím kosterních ostatků. Známo je téměř 150 vzorků, většinou se ovšem jedná o zlomky čelistí nebo jednotlivé zuby. Poměrně hojně jsou nalézány i téměř celé nebo částečně dochované lebky, velmi málo jsou naopak dochované pozůstatky trupu a končetin.[2]
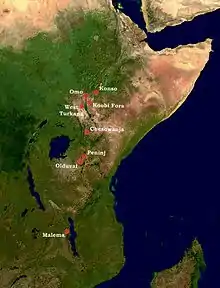
Kosti druhu Paranthropus boisei jsou nalézány v dlouhém pásu od střední Etiopie až po severní Malawi. Nejvíce dokladů ovšem pochází z oblasti Koobi Fora na východním břehu jezera Turkana.[3]

První pozůstatky zachytil Louis Leakey již roku 1955 v rokli Olduvai v Tanzanii. Jednalo se však pouze o dva samostatné zuby – mléčný špičák a stoličku – takže jejich přesné taxonomické určení bylo prakticky nemožné.[2]
To se změnilo 17. července 1959, když Mary Leakey v Olduvajské rokli odhalila téměř celou, dobře dochovanou lebku s podobně utvářenými zuby. Tento výjimečný nález dostal kromě katalogového čísla OH 5 (tj. Olduvai hominid 5) přezdívku Louskáček (Nutckracker) a stal se ve své době velkou senzací.[4] Tehdy se jednalo o nejstaršího známého hominida z východní Afriky[5] a i přes další nálezy, které ji svým stářím nebo významem později překonaly, je lebka OH 5 stále jedním z nejznámějších nálezů z Olduvajské rokle. Louis Leakey pro ni původně vyčlenil zcela nový rod Zinjanthropus s druhem Zinjanthropus boisei – „Zinj“ je staré označení pro východní Afriku, zatímco druhový přívlastek připomíná Charlese Boise, jednoho ze sponzorů výprav do Olduvai.[4] Celek lze tudíž přeložit jako „východoafrický člověk Boiseův“.[6]
Většina odborníků se však domnívala, že jde o variantu jihoafrického druhu Paranthropus robustus, a proto se již od 60. let začal uplatňovat název Paranthropus boisei nebo Australopithecus boisei.[7]
V roce 1964 byla na lokalitě Peninj u tanzanského jezera Natron zachycena dolní čelist, která dobře doplnila lebku Louskáčka.[8] Množství ostatků bylo zjištěno v 70. letech na dolním toku řeky Omo a především v oblasti Koobi Fora na východním břehu jezera Turkana. Několik zlomků bylo získáno také při západním břehu jezera Turkana a na lokalitě Chesowanja u jezera Baringo v Keni.[9]
Další významné nálezy pocházejí z přelomu 20. a 21. století. Tak byla v říjnu roku 1993 na etiopské lokalitě Konso zjištěna velmi dobře dochovaná lebka, a to včetně spodní čelisti. Tím se známý areál výskytu robustních australopitéků posunul dále na sever.[10] Naopak jižní okraj rozšíření druhu Paranthropus boisei tvoří lokalita Malema až na severu Malawi, odkud byl v roce 1999 popsán nález části horní čelisti se silně opotřebenými stoličkami.[11]
_Mary_Douglas_Nicol_Leakey_(1913-1996)_and_her_husband_Louis_Seymour_Bazett_Leakey_(1903-1972).jpg.webp)
Velmi důležitý objev byl učiněn také v letech 2010–2011, kdy byla v Olduvajské rokli zachycena částečná kostra OH 80. Ta představuje první zlomky postkraniálního skeletu, které bylo možné s jistotou přiřknout druhu P. boisei, protože se nalezly spolu s devíti zuby.[12] U jiných ostatků končetin, především z oblastí Omo a Koobi Fora však nelze spolehlivě rozhodnout, zda patří druhu Paranthropus boisei nebo jiným, soudobým hominidům (Homo habilis, Homo erectus).[9]
Datace
Nálezy druhu Paranthropus boisei pocházejí z lokalit, jejichž sedimenty lze velmi dobře datovat pomocí radiometrických metod.[13] Právě při nálezu Louskáčka v Olduvajské rokli byla přitom poprvé aplikována metoda draslík/argon. Ta ukázala, že vývoj člověka trval mnohem déle než asi půl milionu let, jak se v té době často předpokládalo.[14]
Nejstarší nálezy P. boisei jsou nyní známé z Malawi a dosahují stáří 2,3–2,5 milionu let.[11] Naopak nejmladší doklady druhu z etiopské lokality Konso jsou staré jen 1,4 milionu let[10] a nově zachycené pozůstatky částečné kostry z Olduvai dokonce jen 1,34 milionu let.[12] Jen o málo starší jsou další nálezy z tanzanských lokalit Olduvai a Peninj (před 1,7–1,4 milionu let). V Koobi Fora a na dolním toku řeky Omo jsou ostatky P. boisei nalézány téměř z celého období výskytu tohoto druhu, tedy z doby před 2,3–1,4 milionu let.[13]
Popis
Morfologicky byl Paranthropus boisei nejodvozenějším ze všech australopitéků. Jeho lebka byla mohutnější než u ostatních druhů, včetně jihoafrického Paranthropus robustus.[4] Měl ze všech homininů největší stoličky a třenové zuby, nejsilnější sklovinu a nejrobustnější čelisti, zatímco řezáky a špičáky byly nezvykle silně redukované.[15] Proto je často označován jako hyperrobustní australopiték.[1] Díky mnoha unikátním znakům je navíc jedním z nejsnáze identifikovatelných vyhynulých homininů.[16]
Stejně jako pro ostatní australopitéky je i pro tento druh typický výrazný pohlavní dimorfismus, srovnatelný se současnými gorilami.[17] Samci v průměru dosahovali výšky 137 cm a samice 124 cm. Hmotnost samců dosahovala asi 50 kg, zatímco samice vážily kolem 34 kg.[14] Odhady celkové velikosti jsou ovšem odvozeny z velmi nekompletních ostatků, takže je nové objevy mohou změnit.
Objem mozku nalezených lebek se v závislosti na velikosti jedince pohybuje v rozmezí 400–545 cm³ s průměrem 488 cm³.[13] V porovnání se staršími druhy australopitéků lze tudíž pozorovat mírný nárůst průměrné velikosti, může se ovšem jednat jen o důsledek celkově větších tělesných rozměrů a výrazného pohlavního dimorfismu.[18] V porovnání s rodem Homo je případné zvětšování mozku jen nepatrné. Změny v uspořádání mozku zatím nebyly jednoznačně prokázány, celkově se struktura blíží spíše lidoopům než lidem.[19]

- předozadně krátký, úzký tvar
- malý objem (475–545 cm³, průměrně 488 cm³)
- nápadné úpony mohutných šíjových svalů, vyvažujících hlavu s masivními čelistmi (šíjový hřeben crista nuchae)
- u větších jedinců – patrně samců – i nápadný úpon dobře vyvinutého spánkového svalu (šípový hřeben crista sagittalis)
- velká a silně pneumatizovaná spánková kost (os temporale) podobně jako u šimpanzů
- unikátní zesílení (překrytí) švu mezi spánkovou a temenní kostí jako adaptace na tlak silných spánkových a žvýkacích svalů
- výrazné zúžení lebky za očnicemi, poskytující dostatek prostoru pro velký spánkový sval
- hluboká, laterálně rozšířená jamka pro připojení dolní čelisti (fossa mandibularis)
- kratší, vpřed posunutý velký týlní otvor (foramen magnum) s unikátním srdčitým tvarem, odlišný od delšího, oválného či kruhového otvoru ostatních australopitéků
- týlní kosti s otisky žilních splavů sulcus sinus occipitalis a sulcus sinus marginalis, stejně jako u dalších robustních australopitéků a druhu Australopithecus afarensis

- vertikální, nápadně plochý profil bez výrazně vystupujících čelistí
- robustní, zpředu nápadně široká, miskovitá tvář s vystupujícími lícními kostmi a plochým nosem
- nízké, vzad ubíhající čelo
- výrazné nadočnicové oblouky
- „frontal trigone“ - trojúhelníkovitý útvar tvořený vkleslinou čelní kosti nad očnicovým valem, vymezenou výraznými náběhy k šípovému hřebeni
- kulaté očnice, posazené daleko od sebe
- nosní otvor obvykle hruškovitého tvaru, se zaoblenými postranními hranami
- chybějící zpevňující prvky obličeje (přední pilíře), typické pro jihoafrické australopitéky
- široké, vpřed silně vyklenuté lícní kosti, začínající již nad třenovými zuby – ukazují na dobře vyvinutý zevní žvýkací sval (musculus masseter)
- masivní dolní čelist, větší a širší než u jakéhokoliv jiného hominina
- chybějící bradový výběžek dolní čelisti
- celkově velmi robustní žvýkací aparát
- klenuté patro, tvořené výrazně zesílenými kostmi
- velmi malé řezáky a špičáky v porovnání se zadními zuby, menší než u druhů P. aethiopicus a P. robustus
- extrémně velké zadní zuby – stoličky, třenové zuby – s rozměry až 2 x 2 cm (největší v rámci všech známých homininů)
- třenové zuby výrazně molarizované (utvářené podobně jako stoličky - symetrické, s dvěma hrbolky)
- extrémně silná, rychle se tvořící vrstva skloviny – korunky zubů rostou rychleji než u ostatních homininů
Způsob života
Studie se díky unikátní morfologii lebky a zubů od počátku soustředily především na stravu a životní prostředí východoafrických paranthropů, což bylo na přelomu 20. a 21. století ještě posíleno moderními metodami. Mnohé stránky života lze však studovat jen s obtížemi i kvůli chybějícím nálezům postkraniální kostry.
Jen málo odborníků pochybuje o tom, že se Paranthropus boisei pohyboval vzpřímenou dvounohou chůzí. Dochované fragmenty dolních končetin naznačují plnou adaptaci k bipedii, zatímco prodloužené předloktí, tvořené mohutnými kostmi se silnými svaly svědčí o častém využívání stromového patra, a to i přes nápadně mohutnou stavbu těla.[2]
Dosud neexistují doklady, že by P. boisei používal nebo vyráběl nástroje. Louis Leakey jej sice po nálezu Louskáčka v roce 1959 pokládal za tvůrce oldovanských nástrojů, později však bylo prokázáno, že v oblasti žil ve stejnou dobu také Homo habilis a je pravděpodobnější, že právě on je původcem těchto artefaktů.[7]
Životní prostředí
Velká část badatelů předpokládá, že Paranthropus boisei dával přednost vlhčím místům, podmáčeným loukám nebo mokřinám při březích jezer a vodních toků.[20] Zatímco starší analýzy často ukazovaly spíše na uzavřené prostředí hustších lužních porostů, novější výzkumy naznačují život i v otevřeném prostředí savany, jen s řídkými ostrůvky otevřených lesíků a křovin.[2] Ekologická tolerance tedy mohla být poměrně široká podobně jako například u druhu Australopithecus afarensis.[21] Zdá se, že určujícím faktorem nebyla přítomnost nebo absence stromového porostu, ale dostatek vody, protože i sušší travnaté lokality (Malema, Konso) se vždy nachází v blízkosti vodního toku nebo jezera.[20] Také analýza izotopů kyslíku odpovídá silné závislosti na vodních zdrojích – Paranthropus boisei nepatřil k druhům, které jsou schopné potřebu vody pokrýt konzumovanou potravou.[20]
V každém případě Paranthropus boisei obýval otevřenější prostředí než starší P. aethiopicus. Dokonce nelze vyloučit, že právě šíření travnatých plání mohlo stimulovat přerod P. aethiopicus v P. boisei.[22]
Strava
Mohutné čelisti a zuby mají silně odvozenou, specializovanou stavbu, která se navíc v průběhu dlouhého výskytu druhu prakticky nezměnila.[23] Malé přední „krájecí“ zuby doplňovaly extrémně velké zadní „mlecí“ zuby, uložené v téměř paralelních řadách.[24] Unikátní byla velmi rychlá tvorba zubní skloviny na korunkách stoliček, která mohla souviset s nároky konzumované potravy.[25]
Od prvních objevů tak převládal názor, že paranthropové byli na rozdíl od gracilních australopitéků a prvních lidí výhradně býložraví, úzce specializovaní na konzumaci tvrdé a křehké potravy. Také přezdívka Louskáček, která se vžila pro lebku OH 5 z Olduvai, vychází z tohoto předpokladu.[26]

Až v novější době se objevily názory, že čelisti a zuby, schopné vyvinout a snést značnou sílu při žvýkání, nemusí být nutně důsledkem konzumace tvrdé stravy. Zvýšenou odolnost si mohlo vyžádat i zpracování měkčích, ale málo výživných zdrojů. Ty samy o sobě nevyžadovaly takovou sílu, bylo však nutné je důkladně rozžvýkat. Kvůli nízkému obsahu živin jich navíc bylo třeba přijímat velké množství.[26] Současně někteří badatelé uvažují o tom, že specifické rysy na lebce a na zubech nemusely být způsobeny úzkou specializací druhu, ale naopak rozšířením spektra přijímané potravy.[27]
Velké stoličky a třenové zuby odpovídají výraznému namáhání a potřebě velké skusové plochy. Naproti tomu velmi malé řezáky ukazují, že přední zuby nebyly v porovnání se stoličkami tak důležité. To znamená, že východoafričtí paranthropové mohli jíst drobné plody, listy, semena či ořechy, které nebylo nutné před rozdrcením zadními zuby dlouze dělit na menší kusy (pokud k tomuto účelu nepoužívali nástroje, což zatím nebylo doloženo).[27] Korunky stoliček ovšem postrádají ostré hrany, jaké mají moderní gorily i jiní listožraví primáti k důkladnému zpracování listů nebo travních čepelí.[20]
Analýza izotopů uhlíku ze zubní skloviny odpovídá výrazně zvýšené konzumaci rostlin z otevřeného prostoru savany, tvořících 75–80 % všech přijímaných potravních zdrojů.[26] Tím se P. boisei nápadně liší od starších homininů, kteří z velké části využívali i potravu z lesních porostů (Ardipithecus ramidus vyhledával dokonce 80–95 % veškeré potravy na stromech nebo v křovinách).[20]

Překvapivé výsledky přineslo i studium mikroskopických poškození zubní skloviny, které se nápadně liší od jihoafrického druhu Paranthropus robustus a podobají se spíše starším australopitékům (A. anamensis, A. afarensis) či moderním primátům (mandril, pavián, šimpanz, orangutan).[27] Chybí množství odštěpků a důlků, které vznikají v důsledku konzumace tvrdých objektů a naopak se hojně vyskytují jemné rýhy a škrábance, ukazující na žvýkání tuhé, vláknité stravy.[20] Stejně jako morfologie zubů a celé lebky se přitom ani struktura mikroskopických stop v průběhu celého výskytu druhu P. boisei nijak výrazně neměnila.[28] U mnoha nalezených ostatků byly také zjištěny výrazně obroušené korunky zubů, což může svědčit o silně abrazivní potravě.[14] Obrušování skloviny mohly způsobovat zbytky zeminy na podzemních zásobních orgánech, ale také křemíkaté fytolity, které se hojně nacházejí v travinách i šáchorovitých.[29]
Proto se oproti původním předpokladům nejčastěji uvažuje o tom, že Paranthropus boisei se v pobřežních mokřinách živil převážně rostlinami z čeledi šáchorovitých.[18] Kromě nejčastěji uváděného šáchoru papírodárného, tj. papyru (Cyperus papyrus) se uvažuje o konzumaci druhů Cyperus articulatus, Cyperus esculentus, Cyperus immensus, Cyperus laevigatus, Cyperus longus, Cyperus rotundus nebo Cyperus usitatus.[18] Jen menší část potravy pak mohlo tvořit ovoce, semena, ořechy, případně hmyz nebo drobní obratlovci. Stébla i podzemní zásobní orgány šáchorovitých (oddenky, cibulky nebo hlízy) jsou tuhé a vláknité. Nutriční hodnota u některých druhů (Cyperus papyrus) sice přibližně odpovídá domestikovaným bramborám, jedlé části jsou však značně dřevnaté a náročné na konzumaci. Šáchorovité jsou navíc v mokřinatých oblastech východní Afriky hojné a dostupné po celý rok, zatímco traviny jsou silně sezónní.[20]
Fylogeneze
Paranthropus boisei je nejmladším zástupcem paranthropů i všech australopitéků. Právě jím tyto druhy homininů vymírají. Ve východní Africe schází dobře prostudované lokality z doby před 1–1,4 miliony let, a proto nelze vyloučit, že robustní australopitékové se zde vyskytovali ještě v této době. Podle většiny autorů je to ovšem málo pravděpodobné.[13]
Důvod zániku druhu dosud nebyl uspokojivě vysvětlen. Obvykle se předpokládá, že paranthropové se již nedokázali přizpůsobit dalšímu vysychání a souvisejícím změnám prostředí, v němž nedokázali najít dostatek potravy. To ovšem není plně v souladu s velmi dlouhým výskytem tohoto druhu bez výraznějších změn v morfologii a také s faktem, že ostatky P. boisei jsou na známých lokalitách výrazně početnější než nálezy soudobých zástupců rodu Homo. Důvodem vyhynutí paranthropů (respektive australopitéků) ovšem nemusely být jen samotné klimatické změny, ale i neschopnost konkurovat mladším formám rodu Homo (především Homo erectus), využívajícím stejné potravní zdroje.[30]
Paranthropus boisei morfologicky velmi těsně navazuje na druh Paranthropus aethiopicus, který jej utvářením lebky i dobou výskytu spojuje s druhem Australopithecus afarensis do jedné vývojové linie. Morfologická blízkost P. aethiopicus a P. boisei je dokonce tak úzká, že někteří odborníci starší z druhů neuznávají a veškeré východoafrické nálezy robustních čelistí a zubů určují jako P. boisei.[31] Většinou se však dva druhy alespoň formálně vydělují, přičemž se často předpokládá, že jsou důsledkem vývoje anagenezí. Hranice mezi oběma taxony se klade zhruba do doby před 2,3 miliony let.[32] Předpoklad anageneze podporují nálezy z dolního toku řeky Omo, které na sebe morfologicky i časově dobře navazují. Zhruba před 2,3 miliony let zde podle dosavadních výzkumů vrcholily změny přírodního prostředí (šíření otevřených travin na úkor lesů), které mohly stimulovat změny i mezi homininy.[22]
Převážná část odborníků se shoduje, že P. boisei je blízce příbuzný také s jihoafrickým Paranthropus robustus a že oba druhy tvoří monofyletickou skupinu. Tomu zatím nasvědčuje i většina srovnávacích studií.[13] Přesto nelze vyloučit, že P. boisei a P. robustus společného předka nesdílí. To by ovšem znamenalo, že robustní žvýkací aparát se u těchto homininů vyvinul nezávisle na sobě, paralelním vývojem.[32]
Významné nálezy

KNM-ER 406
Téměř kompletní lebka dospělého samce z Koobi Fora, stará 1,7 milionu let.
KNM-ER 732
Zčásti dochovaná lebka samice z Koobi Fora, stará 1,7 milionu let. Oproti jiným ostatkům je méně robustní, má menší tvář a postrádá sagitální hřeben na temeni. Lebka tak názorně ukazuje rozdíly mezi pohlavími u druhu P. boisei.
KNM-ER 1500
Zlomkovitě dochovaná kostra z Koobi Fora, která se na základě fragmentu dolní čelisti obvykle řadí k druhu Paranthropus boisei. Kostra, dosahující stáří 1,8–1,9 milionu let byla nalezena v roce 1972. Kromě zlomku dolní čelisti zahrnuje špatně dochované pozůstatky vřetenní, loketní, stehenní, holenní i lýtkové kosti a článek prstu.
.JPG.webp)
OH 5 (Louskáček, Zinj)
Velmi dobře dochovaná lebka, typový nález druhu Paranthropus boisei. Lebku, starou 1,85 milionu let zachytila 17. července 1959 Mary Leakey v tanzanské roklině Olduvai. Nález vzbudil velký ohlas odborné i laické veřejnosti, protože byl prvním dobře dochovaným pozůstatkem hominida z východní Afriky. Kvůli robustní stavbě získal přezdívku Louskáček (Nutckracker).[6] Odhalen byl ve stejné úrovni jako artefakty oldovanské kultury a početné zvířecí kosti. Protože lebka zůstala na rozdíl od ostatních kostí téměř neporušená, považoval Louis Leakey Louskáčka za tvůrce nástrojů, který dotyčná zvířata lovil a jedl. V té době by byl nejstarším známým výrobcem nástrojů, a proto jej Leakey původně zařadil do nového rodu jako Zinjanthropus boisei, aby jej odlišil od australopitéků, u nichž se výroba nástrojů nepředpokládala.[4] Až pozdějšími výzkumy bylo zjištěno, že nástroje z Olduvai vyráběl spíše Homo habilis, který se zde vyskytoval souběžně s paranthropy.[7]

OH 80
Dosud nejúplněji dochovaná kostra, která může být s jistotou přiřazena druhu P. boisei, protože současně s ostatky končetin (kost pažní, vřetenní, stehenní, holenní) se dochovalo také devět zubů. Určení je tak na rozdíl od kostry KNM-ER 1500 mnohem spolehlivější. Zlomky kostí jsou staré jen 1,34 milionu let a jsou tak zároveň nejmladším dokladem druhu. Ukazují, že robustní stavbu neměly pouze čelisti a lebka, ale také končetiny. Dochované zlomky jsou přesto velmi nekompletní, navíc nesou nápadné stopy po ohryzu masožravci.[12]
RC 911
Fragment horní čelisti se dvěma stoličkami z Malema, starý 2,3–2,5 milionu let patří k nejstarším pozůstatkům druhu. Lokalita Malema rozšiřuje areál výskytu P. boisei výrazně k jihu a může naznačovat komunikační propojení jihoafrických a východoafrických nalezišť. Dosud jsou lokality z jižní i východní Afriky geograficky striktně oddělené rozsáhlým územím bez nálezů, takže jejich vzájemné vztahy jsou nejasné.[1]
Odkazy
Reference
- VANČATA, V. Paleoantropologie - přehled fylogeneze člověka a jeho předků. Brno: Akademické nakladatelství CERM, 2003. 212 s. ISBN 80-7204-272-6. S. 18.
- WOOD, B. A.; RICHMOND, B. G. Human evolution: taxonomy and paleobiology. Journal of anatomy. 2000, roč. 196, s. 19–60.
- SILVERMAN, N.; RICHMOND, B. G.; WOOD, B. Testing the taxonomic integrity of Paranthropus boisei sensu stricto. American Journal of Physical Anthropology. 2001, roč. 115, s. 167–178. Dostupné online.
- LEAKEY, L. S. B. A new fossil skull from Olduvai. Nature. 1959, roč. 184, s. 491–493.
- Starší nálezy představovaly jen nepočetné, malé zlomky čelistí nebo jednotlivé zuby, které nebylo možné blíže určit.
- CONROY, G. C. Reconstructing human origins. New York: W. W. Norton & company, 1997. 547 s. Dostupné online.
- CARTMILL, M.; SMITH, F. H. The Human Lineage. 1. vyd. Oxford: Wiley-Blackwell, 2009. 624 s. Dostupné online. ISBN 978-0471214915.
- JELÍNEK, J. Velký obrazový atlas pravěkého člověka. 1. vyd. Praha: Artia, 1972. 560 s.
- CONSTANTINO, P.; WOOD, B. A. The evolution of Zinjanthropus boisei. Evolutionary Anthropology: Issues, News, and Reviews. 2007, roč. 16, s. 49–62.
- SUWA, G., ASFAW, B., BEYENE, Y., WHITE, T. D., KATOH, S., NAGAOKA, S., NAKAYA, H., UZAWA, K., RENNE, P., WOLDEGABRIEL, G. The first skull of Australopithecus boisei. Nature. 1997, roč. 389, s. 489–492.
- KULLMER, O., SANDROCK, O., ABEL, R., SCHRENK, F., BROMAGE, T. G., JUWAYEYEI, Y. M. The first Paranthropus from the Malawi Rift. Journal of Human Evolution. 1999, roč. 37, s. 121–127.
- DOMÍNGUEZ-RODRIGO, M., PICKERING, T. R., BAQUEDANO, E., MABULLA, A., MARK, D. F., MUSIBA, C., et al. First partial skeleton of a 1.34-million-year-old Paranthropus boisei from Bed II, Olduvai Gorge, Tanzania. PloS one. 2013, roč. 8, s. e80347.
- WOOD, B. A.; CONSTANTINO, P. Paranthropus boisei: fifty years of evidence and analysis. American journal of physical anthropology. 2007, roč. 134/S45, s. 106–132.
- ROBERTSOVÁ, A. Evoluce: Příběh člověka. Praha: Knižní klub, 2012. 256 s.
- STRAIT, D. S. The evolutionary history of the australopiths. Evolution Education Outreach. 2010, roč. 3, s. 341–352.
- KIMBEL, W. H. The species and diversity of australopiths. In: HENKE, W.; TATTERSALL, I. Handbook of Palaeoanthropology 3. Heidelberg: Springer, 2007. S. 1539–1573.
- WOOD, B. A. Human evolution. Bioessays. 1996, roč. 18, s. 945–954. Dostupné online.
- STEWART, K. M. Environmental change and hominin exploitation of C4-based resources in wetland/savanna mosaics. Journal of Human Evolution. 2014, roč. 77, s. 1–16.
- FALK, D., REDMOND, J. C. J., GUYER, J., CONROY, C., RECHEIS, W., WEBER, G. W., SEIDLER, H. Early hominid brain evolution: a new look at old endocasts. Journal of Human Evolution. 2000, roč. 38, s. 695–717.
- GRINE, F. E., SPONHEIMER, M. J., UNGAR, P. S., LEE-THORP, J., TEAFORD, M. F. Dental microwear and stable isotopes inform the paleoecology of extinct hominins. American Journal of Physical Anthropology. 2012, roč. 148, čís. 285–317.
- BOCHERENS, H., SANDROCK, O., KULLMER, O., SCHRENK, F. Hominin palaeoecology in Late Pliocene Malawi: First insights from isotopes (13C, 18O) in mammal teeth. South African Journal of Science. 2011, roč. 107, s. #331.
- ALEMSEGED, Z. An integrated approach to taphonomy and faunal change in the Shungura Formation (Ethiopia) and its implication for hominid evolution. Journal of Human Evolution. 2003, roč. 44, s. 1–28.
- WOOD, B. A.; LIEBERMAN, D, E. Craniodental variation in Paranthropus boisei: a developmental and functional perspective. American Journal of Physical Anthropology. 2001, roč. 116, s. 13–25. Dostupné online.
- COPPENS, Y. Hominid evolution and the emergence of the genus Homo. Pontifical Academy of Sciences Scripta Varia. 2013, roč. 121, s. 1–15.
- ROBSON, S. L.; WOOD, B. A. Hominin life history: reconstruction and evolution. Journal of Anatomy. 2008, roč. 212, s. 394–425.
- UNGAR, P. S.; SPONHEIMER, M. J. The diets of early hominins. Science. 2011, roč. 334, s. 190–193.
- WOOD, B. A.; STRAIT, D. S. Patterns of resource use in early Homo and Paranthropus. Journal of Human Evolution. 2004, roč. 46, s. 119–162.
- DAEGLING, D. J, JUDEX, S., OZCIVICI, E., RAVOSA, M. J., TAYLOR, A. B., GRINE, F. E., TEAFORD, M. F., UNGAR, P. S. Feeding mechanics, diet, and dietary adaptations in early hominins. American Journal of Physical Anthropology. 2013, roč. 151, s. 356–371.
- RABENOLD, D.; PEARSON, O. M. Abrasive, silica phytoliths and the evolution of thick molar enamel in primates, with implications for the diet of Paranthropus boisei. PLoS ONE. 2011, roč. 6, s. e28379.
- ŠMAHEL, Z. Příběh lidského rodu. 1. vyd. Brno: Moravské zemské muzeum, 2005. 85 s. ISBN 80-7028-262-2.
- STRAIT, D. S.; GRINE, F. E.; MONIZ, M. A. A reappraisal of early hominid phylogeny. Journal of Human Evolution. 1997, roč. 32, s. 17–82.
- ALEMSEGED, Z.; COPPENS, Y.; GERAADS, D. Hominid cranium from Omo: description and taxonomy of Omo-323-1976-896. American Journal of Physical Anthropology. 2002, roč. 117, s. 103–112. Dostupné online.
Literatura
- ROBERTSOVÁ, A. Evoluce: Příběh člověka. Praha: Knižní klub, 2012. 256 s.
- SVOBODA, J. Čas lovců: Aktualizované dějiny paleolitu. Brno: Akademické nakladatelství CERM, 2009. 298 s.
- SVOBODA, J. Předkové. Evoluce člověka. Praha: Academia, 2014. 480 s.