Paranthropus robustus
Paranthropus robustus (česky též Australopiték robustní)[1] je druh vyhynulého hominida, žijící ve starším pleistocénu, před 2–1,5 miliony let v jižní Africe, na území dnešní Jihoafrické republiky. Patří k zástupcům robustních australopitéků, řazených často do samostatného rodu Paranthropus. Žil již současně se zástupci rodu Homo (Homo habilis, Homo erectus).
Stratigrafický výskyt: pleistocén | |
|---|---|
.JPG.webp) Lebka druhu Paranthropus robustus | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Třída | savci (Mammalia) |
| Řád | primáti (Primates) |
| Čeleď | Hominidé (Hominidae) |
| Podčeleď | Homininae |
| Tribus | Hominini |
| Rod | Paranthropus Broom, 1938 |
| Binomické jméno | |
| Paranthropus robustus Broom, 1938 | |
| Některá data mohou pocházet z datové položky. | |
Objevy
Poměrně hojné ostatky druhu Paranthropus robustus jsou bez výjimky nalézány v jihoafrických krasových jeskyních. Všechny známé lokality – Kromdraai, Swartkrans, Drimolen, Cooper's a Gondolin – jsou přitom soustředěny na nevelkém území v údolí Blaauwbank nedaleko Johannesburgu.[2] Ačkoliv první kosti byly získány v Kromdraai, drtivá většina jich pochází ze Swartkrans.[3] Početnější jsou též ostatky z Drimolen, zatímco z ostatních lokalit je k dispozici jen hrstka nálezů.
Paranthropus robustus byl identifikován již v roce 1938 a představuje tudíž typový druh rodu Paranthropus. Zachycen byl krátce poté, co Raymond A. Dart představil veřejnosti Taungské dítě, nejstarší nález australopitéka (druh Australopithecus africanus), který byl ovšem ostatními badateli zprvu ostře odmítán. Takřka jediným, kdo názory R. A. Darta v té době přijal a podporoval, byl původem skotský lékař Robert Broom, který se po získání práce v Transvaalském muzeu v Pretorii roku 1934 sám pustil do pátrání po dalších ostatcích. Dne 8. června 1938 pak od správce vápencového lomu ve Sterkfontein zakoupil horní čelist hominida, kterou popsal pod katalogovým číslem TM 1517. Kvůli zřetelným odlišnostem od dosavadních nálezů australopitéků pro ni navrhl zcela nový druh Paranthropus robustus.[4] Jak později zjistil, čelist objevil školák Gert Terblanche v jeskyni Kromdraai. Tamtéž Broom poté dohledal i velkou část lebky a další ostatky.

Během druhé světové války byly odkryvy v jižní Africe přerušeny, zejména kvůli zastavení těžby vápence.[5] Pokračovaly však práce na vydání monografie o jihoafrických australopitécích, která nakonec vyšla v roce 1946.[6] Koncem čtyřicátých let 20. století již bylo nálezů druhu Australopithecus africanus i Paranthropus robustus tolik, že odborná veřejnost konečně začala Dartovy a Broomovy objevy brát vážně.[7][8]
V listopadu roku 1948 byly doklady robustních australopitéků zjištěny také ve Swartkrans, vzdáleném jen asi 1,5 km od Kromdraai, na protilehlém břehu údolí Blaauwbank. Během tří let intenzivní práce zde bylo získáno velké množství ostatků. Vzhledem k mírným odlišnostem od nálezů z Kromdraai je Robert Broom popsal jako nový druh Paranthropus crassidens.[9] Dnes je ovšem většina odborníků spojuje s nálezy z Kromdraai do jediného druhu (P. robustus).[10][11]
Po celou druhou polovinu 20. století pak v Kromdraai i Swartkrans probíhaly rozsáhlé systematické výzkumy, jimiž se zabýval především C. K. Brain. Odhaleny tak byly stovky kostí druhu P. robustus.[12]
Roku 1994 byl oznámen nález dvou zubů P. robustus z jeskyně Sterkfontein, která jinak vydává nálezy druhu Australopithecus africanus.[2]

V roce 1999 pak byl publikován také nezvykle velký zub hominida z jeskyně Gondolin.[13] Ačkoliv kvůli jeho značným rozměrům panují určité pochybnosti o taxonomickém určení, většina autorů jej řadí k druhu P. robustus.[2]
Výrazně bohatší jsou ovšem kolekce, získávané od roku 1994 v jeskyni Drimolen.[14] Významné místo mezi nimi zaujímá velmi dobře dochovaná lebka DNH 7, představující dosud nejkompletnější známou lebku australopitéka.[15]
Poslední nově odhalenou lokalitou je jeskyně Cooper's, kde byly ostatky P. robustus (izolované zuby, zlomky lebky a dolních čelistí, poškozený bederní obratel a několik článků prstů) odhaleny v červnu roku 2001.[16] V této jeskyni byla stolička hominida objevena již roku 1938.[17] Tehdy byla určena jako Plesianthropus transvaalensis (dnes Australopithecus africanus), přestože podle popisu by se mohlo jednat spíš o druh Paranthropus robustus. Nyní je ovšem zub ztracený a jeho taxonomické zařazení tedy nelze ověřit.[18]
V roce 2007 je tak pro druh Paranthropus robustus uváděno téměř 500 kusů jednotlivých nálezů. Na 350 zlomků pochází ze Swartkrans, 31 kusů z Kromdraai a více než 80 vzorků z Drimolen. Z velké části se jedná o jednotlivé zuby nebo zlomky čelistí a lebek, doložené jsou však i fragmenty postkraniální kostry.[19]
Datace
Jihoafrické krasové jeskyně jsou oproti východoafrickým lokalitám jen obtížně datovatelné. V jejich výplni chybí vrstvy sopečného popela, na něž lze dobře aplikovat radiometrické datovací metody. Uplatňují se tak především přístupy relativní chronologie. Tradiční je využití biostratigrafie, tedy datace na základě přítomnosti určitých živočišných druhů. Jejich výskyt je přitom srovnáván s dobře datovatelnými lokalitami východní Afriky.[20] Teprve v posledních letech technický rozvoj umožnil uplatnění absolutních datovacích metod i v jižní Africe – využíván je paleomagnetismus,[21] termoluminiscence[22] nebo metoda uran-olovo.[23] Tyto nové přístupy mnohdy výrazně zpřesnily starší údaje, které často představovaly značně široké časové intervaly.
Nejčastěji se tak výskyt druhu Paranthropus robustus uvádí v době před 2–1,5 miliony let,[24] vyloučit však nelze ani jeho přežívání až do doby asi před 600 000 lety.[25] P. robustus tak mohl podobně jako jiné druhy savců v jižní Africe přežívat výrazně déle než ve východní části kontinentu, doklady jsou však dosud nejasné.[26]

Jednotlivé lokality jsou k roku 2011[27] datovány takto:
- Swartkrans (Member 1) – před 2,3–1,8 miliony let
- Drimolen – před 2,0–1,5 miliony let
- Gondolin – před 1,78 miliony let
- Kromdraai (Member 3) – před 1,78–1,65 miliony let
- Swartkrans (Member 2) – před 1,65–1,07 miliony let
- Cooper's – před 1,5–1,4 miliony let
- Sterkfontein (Member 5) – před 1,38–1,07 miliony let
- Swartkrans (Member 3) – před 1,3–0,62 miliony let
Popis
Paranthropus robustus patří mezi velmi dobře doložené australopitéky, protože je znám z více než 500 jednotlivých vzorků. Poznání biologie a ekologie tohoto druhu však komplikuje problematické datování lokalit.[28] Poměrně hojně jsou v jihoafrických jeskyních nalézány zlomky kostí trupu a končetin. Ne všechny lze ovšem jednoznačně přiřadit druhu P. robustus, protože současně se zde nacházejí také ostatky časných zástupců rodu Homo. Zatímco lebky a zuby obou rodů jsou dostatečně odlišné a díky tomu jednoznačně identifikovatelné, zlomky postkraniální kostry se liší jen málo.[2]

Robustní stavbou se vyznačuje především lebka a zuby, zbytek těla je však jen o málo mohutnější než u ostatních australopitéků.[3] Odhady výšky a váhy z nalezených ostatků činí pro samice zhruba 110 cm při váze 32 kg, proti 132 cm a 40 kg u samců.[29] V celkové velikosti těla se tedy projevuje značný pohlavní dimorfismus, který se ale nijak výrazně nevymyká rozdílům u ostatních druhů australopitéků.[30] Velikost jihoafrických paranthropů však může být podhodnocená, jak naznačuje nápadně velká stolička z Gondolin, zlomky lebek z Drimolen[15] či hlavice stehenní kosti ze Swartkrans.[31] Spektrum nalézaných ostatků zřejmě bylo zkresleno aktivitou predátorů, kteří paranthropy lovili a kteří si záměrně mohli vybírat menší či slabší (tedy snazší) kořist.[32]
I přes přetrvávající malou kapacitu je patrné mírné zvětšení mozkovny a snad i mozku, celková stavba se ale stále blíží spíše lidoopům než lidem. Náznaky změn v uspořádání mozku zatím nebyly jednoznačně prokázány.[33]

- masivní stavba – robustnější než u A. africanus, ale lehčí než u P. boisei
- malý objem (450–650 cm³, průměrně 530 cm³)
- nápadné úpony mohutných šíjových svalů, vyvažujících hlavu s masivními čelistmi (šíjový hřeben crista nuchae)
- u větších jedinců – patrně samců – i nápadný úpon dobře vyvinutého spánkového svalu (šípový hřeben crista sagittalis)
- slabě pneumatizovaná spánková kost (os temporale), podobně jako u zástupců rodu Homo
- výrazné zúžení lebky za očnicemi, poskytující dostatek prostoru pro velký spánkový sval
- hluboká jamka pro připojení dolní čelisti (fossa mandibularis)
- velký, silně pneumatizovaný bradavkový výběžek (processus mastoideus)
- dlouhý, výrazně laterálně umístěný vnější zvukovod podobně jako u A. africanus, na rozdíl od krátkého zvukovodu moderních lidí
- vyklenutá spodina lebeční
- vpřed posunutý velký týlní otvor (foramen magnum) oválného tvaru
- týlní kosti s otisky žilních splavů sulcus sinus occipitalis a sulcus sinus marginalis, stejně jako u dalších robustních australopitéků a druhu Australopithecus afarensis

- vertikální, nápadně plochý profil bez výrazně vystupujících čelistí
- robustní, zpředu nápadně široká, miskovitá tvář s vystupujícími lícními kostmi a plochým nosem (lícní kosti vystupují méně než u P. boisei)
- nízké, vzad ubíhající čelo
- výrazný, nepřerušený nadočnicový oblouk
- „frontal trigone“ – trojúhelníkovitý útvar tvořený vkleslinou čelní kosti nad očnicovým valem, vymezenou výraznými náběhy k šípovému hřebeni
- nosní otvor hruškovitého tvaru, se zaoblenými okraji
- „přední pilíře“ – zpevňující útvary po stranách nosního otvoru, které pomáhají odolávat tlaku a namáhání lebky při skusu
- široké, vpřed silně vyklenuté lícní kosti, ukazující na dobře vyvinutý zevní žvýkací sval (musculus masseter)
- silně pneumatizovaná horní čelist
- velká, masivní dolní čelist s nápadně silným tělem a vysokým ramenem (ramus mandibulae)
- chybějící bradový výběžek dolní čelisti
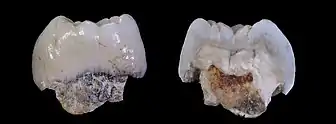
- celkově velmi robustní žvýkací aparát
- klenuté, vzad ubíhající patro, tvořené výrazně zesílenými kostmi
- malé řezáky a špičáky
- redukované špičáky, nevyčnívající z linie ostatních zubů
- extrémně velké zadní zuby – stoličky a třenové zuby (menší než u P. boisei)
- třenové zuby výrazně molarizované (utvářené podobně jako stoličky – symetrické, s dvěma hrbolky)
- první třenové zuby obvykle se třemi kořeny
- extrémně silná, rychle se tvořící vrstva skloviny (již brzy po prořezání zubů se opotřebovává a obrušuje do téměř rovné skusové plochy)
Znaky postkraniální kostry:[2][3]
- robustní paže, v poměru k nohám ještě poměrně dlouhé – kratší než u starších australopitéků, delší než u lidí
- dobře osvalené předloktí, schopné silného úchopu
- loketní kloub stabilní při otáčení podél dlouhé osy – podobně jako u lidoopů, ale odlišně od lidí
- rovné články prstů, ne zakřivené jako u ostatních australopitéků a moderních lidoopů
- karpometakarpální kloub palce velký, pohyblivý, sedlovitého typu jako u lidí
- první metakarp pro skloubení s palcem dlouhý, s širokou hlavicí – podobně jako u lidí
- koncové články prstů ruky opatřené drsnatinou pro masitá bříška (tuberositas phalangis distalis) jako u lidí – odlišně od starších australopitéků a moderních lidoopů
- bederní obratle tvořící bederní lordózu
- stehenní kost s malou hlavicí a dlouhým, předozadně zploštělým krčkem jako u ostatních australopitéků
- palec nohy přitažený k ostatním prstům, ale částečně ještě schopný pohybu do strany
Způsob života
Nalézané kosti trupu a končetin ukazují, že Paranthropus robustus využíval plně vzpřímenou dvojnohou chůzi, ačkoliv tento pohyb se podobně jako u ostatních australopitéků ještě mírně lišil od chůze moderních lidí, například kvůli méně stabilní pánvi nebo pohyblivějšímu palci nohy. Chůze mohla být ve srovnání s lidmi méně výkonná, ale pro způsob života paranthropů zcela dostačující.[34]
Naproti tomu mírně prodloužené paže, tvořené mohutnými, dobře osvalenými kostmi a stabilním loketním kloubem svědčí o přetrvávajícím šplhání ve větvích.[35] Adaptace pro pobyt v korunách stromů ovšem nejsou tak výrazné jako u starších australopitéků – prsty na rukou již například netvoří prohnuté, ale rovné kůstky, méně vhodné pro úchop větví.[2]

Zajímavé údaje o životě paranthropů nabízí nové analýzy zubů a lebek. Srovnání velikosti lebek a stáří jedinců podle stavu zubů ukazuje, že lebky samců s nejopotřebovanějšími zuby jsou největší, zatímco lebky mladých jedinců s dobře dochovanými zuby jsou malé. U druhého pohlaví tomu tak ale není, lebky různě starých samic jsou přibližně stejně velké. To naznačuje, že u samců docházelo k růstu ještě po dosažení dospělosti (tj. pohlavní zralosti) podobně jako u goril, pro které jsou typičtí stříbrohřbetí samci, rostoucí a vybarvující se ještě několik let po dosažení dospělosti.[36] Výjimečné to však není ani pro další moderní primáty s výraznějším pohlavním dimorfismem.[37] Současně analýza ukázala, že právě mladí samci paranthropů byli typickou kořistí predátorů ze Swartkrans – snad proto, že žili osamoceně nebo v malých skupinkách na okraji společnosti, než dosáhli plné zralosti vůdčích samců a založili vlastní skupinu. Byli tak zranitelnější než ostatní.[36]
Další z analýz studovala izotopy stroncia v zubní sklovině homininů. Malí jedinci (samice) druhu P. robustus přitom – na rozdíl od větších jedinců (samců) – vykazovaly hodnoty, které neodpovídaly místu nálezu. To znamená, že samice v době tvorby skloviny (tj. v mládí a dospívání) musely žít jinde. Na základě těchto výsledků autoři studie předpokládají, že samice podobně jako u moderních šimpanzů a goril opouštěly rodné skupiny a přecházely jinam, zatímco samci zůstávali přibližně v místě svého narození.[38] Přesto nelze předpokládat, že by sociální struktura paranthropů plně odpovídala moderním gorilám. Některé prvky mohou být společné, ale přesná moderní analogie pro život druhu P. robustus patrně neexistuje.[38]
Životní prostředí
.jpg.webp)
Stejně jako v případě východoafrického druhu Paranthropus boisei, i na jihu kontinentu je přítomnost P. robustus spojená s vysušováním prostředí, šířením suchomilné vegetace a změnou vlhkých, hustších porostů v suchou savanu.[39]
Přestože okolí krasových jeskyní kolem Johannesburgu mohlo být v pleistocénu o něco vlhčí než v současnosti, žil P. robustus v celkově sušším prostředí než východoafričtí paranthropové.[40] Tvořily je především rozsáhlé travnaté pláně, jen s občasnými skupinkami keřů či stromů. Stejně jako u P. boisei je přitom i v jižní Africe pozorována úzká vazba na vodní zdroje.[41] Na přetrvávající ostrůvky mokřin a podmáčených luk v jinak suché krajině ukazují například nálezy kostí vodušek ze Swartkrans. V těchto vlhčích místech se pak mohly udržet i ostrůvky hustší vegetace, křovin a řídkých lesíků, skýtající paranthropům kromě potravy i možnost úkrytu před dravci.[42] Zatím nelze spolehlivě prokázat, zda jihoafričtí paranthropové upřednostňovali otevřenou travnatou krajinu nebo spíše mokřady a lesíky. Zdá se, že podobně jako východoafrické druhy byli schopní využívat větší spektrum prostředí.[42] Moderní statistické zhodnocení ostatků fauny z dostupných lokalit ale naznačuje spíše na spojení s živočichy lesních porostů.[42]
Strava
Robustní spodní čelist, velké stoličky se silnou sklovinou, zesílení lebky i silné žvýkací svaly ukazují na schopnost vyvinout při skusu velký tlak a konzumovat tak potravu, vyžadující náročné zpracování.[3] Kvůli tomu byl Paranthropus robustus zhruba do 90. let 20. století považován za výhradního býložravce, úzce specializovaného na značně tuhou, vláknitou nebo tvrdou potravu, zatímco Australopithecus africanus i časní zástupci rodu Homo měli být spíše všežraví a málo specializovaní.[43] Objevily se však i názory, že P. robustus své čelisti potřeboval k drcení kostí ostatních savců.[44] Malé řezáky a špičáky vedly k předpokladu, že P. robustus jedl spíše menší plody, semena či ořechy, které nebylo nutné před rozdrcením zadními zuby dlouze dělit na menší kusy. Zmenšování předních zubů však může být kromě jejich menšího významu způsobeno i zvětšováním stoliček a třenových zubů a omezenou velikostí čelistí.[3]
V novější době jsou starší závěry zřetelně upravovány díky výsledkům moderních analýz. Tak výzkum poměru izotopů stroncia a vápníku v kostech P. robustus ukázal již na počátku 90. let 20. století hodnoty blízké spíše lokálním jihoafrickým masožravcům než výhradním býložravcům. To naznačuje, že homininé mohli využívat i živočišné zdroje.[45] Složení kostí ovšem při fosilizaci podléhá změnám a novější analýzy provedené na vzorcích, odebraných ze zubní skloviny se hodnotami blíží spíše moderním paviánům a neukazují na výrazný příjem masa.[46] Obsah stroncia a vápníku je navíc u většiny jihoafrických potravních zdrojů dosud jen málo probádán. Zjištěné hodnoty tak mohou odrážet konzumaci některých plodů.[47] Zároveň byl novými testy ve sklovině zjištěn nízký podíl barya, což by mohlo při zvýšeném obsahu stroncia odpovídat rozsáhlejšímu příjmu hmyzu nebo semen a oddenků některých travin či šáchorovitých.[47]
Mikroskopické poškození skloviny na nalezených zubech tvoří především množství malých důlků a odštěpků. Takové stopy odpovídají moderním konzumentům malých, tvrdých až křehkých objektů, jako jsou mangabej kouřový (Cercocebus atys), mangabej pláštíkový (Lophocebus albigena) nebo malpa hnědá (Cebus apella).[48] Proto se předpokládá, že Paranthropus robustus vyhledával různé plody, lusky či ořechy a semena, ale například i měkkýše.[43] I vnitřní struktura zubní skloviny naznačuje, že P. robustus byl dobře přizpůsoben konzumaci tvrdé potravy.[49] Nevýrazné mikroskopické poškození řezáků a špičáků potvrzuje předpoklad jejich malého významu pro konzumaci, vyloučit ale nelze ani jejich nahrazení jednoduchými nástroji.[41]
Analýza izotopů uhlíku ze zubní skloviny ukazuje na značnou závislost na potravě z lesních porostů (v Africe C3-rostliny), která byla ale z 25–35 % doplňována zdroji otevřených prostor (v Africe C4-rostliny).[50] To odpovídá výše naznačené oblibě prosvětlených lesů, ale současně pronikání do otevřené krajiny.[42] Poměr využití zdrojů z lesních porostů a z otevřených travnatých prostor přitom značně kolísal. Zajímavé také je, že se liší nejen mezi jedinci téhož druhu, ale dokonce i u jednoho jedince v průběhu let, jak bylo zjištěno odebráním vzorků napříč pravidelnými přírůstky skloviny.[29]
C4-rostliny v jižní Africe představují především traviny a šáchorovité. Samotné čepele trav nebo šáchorů nejsou jako potrava pravděpodobné vzhledem k mikroskopickým stopám na zubech paranthropů, uvažuje se proto o konzumaci různých oddenků či hlíz, typických zejména pro šáchory.[43] Takové zdroje se mohly hojně nacházet v mokřinatých oblastech při vodních zdrojích.[51] Na rozdíl od východní Afriky však jen asi třetina jihoafrických druhů šáchorovitých využívá při fotosyntéze C4-cyklus a většina z nich navíc nemá dostatečně vyvinuté nebo poživatelné oddenky.[46] Vyloučena není ani konzumace býložravého hmyzu nebo malých obratlovců.[52][43] Často jsou proto jako potrava pro P. robustus uvažováni termiti. Velká část jihoafrických druhů termitů se však živí dřevem nebo jinými C3-rostlinami, nikoliv trávou.[50]
Pouze termiti a šáchorovité rostliny tedy nemohly zajistit dostatečný podíl C4-zdrojů ve stravě jihoafrických paranthropů, proto se uvažuje i o dalších složkách potravy, jako jsou kobylky, vejce, ještěrky, hlodavci, aj.[50]
Podle veškeré evidence tak Paranthropus robustus nebyl úzce vyhraněný na konkrétní druh obživy, takže utváření lebky a zubů odpovídá spíše rozšíření spektra přijímané potravy než specializaci.[43] Výsledky ukazují, že jihoafričtí paranthropové byli (podobně jako jiní australopitéci i časní zástupci rodu Homo) spíše oportunističtí všežravci s širokou ekologickou tolerancí a rozsáhlou potravinovou základnou.[41] Složení jídelníčku patrně kolísalo v závislosti na sezónní dostupnosti zdrojů, přičemž zdroje z otevřených travnatých prostor byly velmi důležitou, ale silně variabilní součástí přijímané potravy.[53] Některé byly konzumovány po celý rok, zatímco ostatní – především tvrdé a na zpracování náročné kusy – mohly být získávány jen sezónně, jako záložní zdroje v dobách nedostatku jiné potravy.[48]
Kultura
Z jeskyně Swartkrans pochází poměrně početné nálezy kostěných a kamenných nástrojů a kostěné nástroje vydala i jeskyně Drimolen.[54] Vzhledem k současnému výskytu ostatků rodů Homo a Paranthropus na těchto lokalitách je ovšem obtížné přiřadit tyto nástroje konkrétnímu druhu. V počátcích výzkumů byli paranthropové v porovnání s člověkem vnímáni jako primitivní, býložraví tvorové s velkými zuby a malým mozkem, jejichž inteligence nestačila na nástrojovou činnost. Veškeré nalézané artefakty tak byly automaticky přisuzovány soudobým zástupcům rodu Homo a spojovány se získáváním masité potravy.[55]
Nově se však ukazuje, že také P. robustus mohl být schopen používat, ne-li přímo vyrábět nástroje.[3] Utváření ruky se v mnohém blíží spíše člověku než australopitékům – dobře vyvinutý a osvalený, pohyblivý palec i prsty s rovnými články a masitými bříšky odpovídají schopnosti jemné manipulace a tzv. precizního gripu,[56] které jsou klíčové pro efektivní užívání a výrobu nástrojů.[57] Tento názor ovšem nepřijímají všichni odborníci a opírají se přitom i o fakt, že nalezené ostatky nelze vždy s jistotou přiřknout druhu P. robustus.[29] Pravděpodobnost, že tento tvor měl ruce vhodné pro jemnou manipulaci s nástroji je přesto poměrně velká. Užívání nástrojů se navíc předpokládá i u jiných druhů australopitéků (A. afarensis, A. garhi), stejně jako je známé u všech moderních lidoopů i některých primátů.[58]
Zvláště kostěné nástroje ze Swartkrans jsou spojovány s paranthropy, protože část z nich byla nalezena poblíž kostí druhu P. robustus. Již v době nálezu s nimi C. K. Brain prováděl úspěšné experimenty, prokazující vhodnost nástrojů k získávání podzemních hlíz.[55] Nověji jsou kostěné hroty pokládány spíše za nástroje, sloužící k rozbíjení termitišť a získávání termitů.[54] Každopádně na využití kostěných nástrojů mohli být jedinci P. robustus alespoň sezónně poměrně závislí – mohlo se jednat o významný prostředek k získávání obživy.
Fylogeneze
Převážná část odborníků se shoduje, že Paranthropus robustus je blízce příbuzný s východoafrickým Paranthropus boisei a že oba druhy tvoří monofyletickou skupinu se společným předkem. Tomu zatím nasvědčuje i většina srovnávacích studií.[59][60]
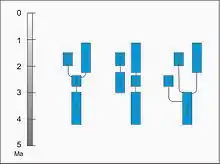
Starší studie předpokládaly, že se P. robustus vyvinul v jižní Africe z druhu Australopithecus africanus a poté se stal předkem východoafrického Paranthropus boisei.[29] Po nálezu Černé lebky (Paranthropus aethiopicus) roku 1985 je však zřejmé, že předka robustních australopitéků je nutné hledat spíše ve východní Africe. Paranthropus aethiopicus totiž utvářením lebky i dobou výskytu velmi dobře překlenuje rozdíly mezi Paranthropus robustus a Australopithecus afarensis a pojí je do jedné vývojové linie. Podle nejčastějšího názoru z druhu P. aethiopicus (navazujícího na druh A. afarensis) vyšel jak východoafrický P. boisei, tak jihoafrický P. robustus.[61] Přesto nelze vyloučit, že P. boisei a P. robustus společného předka nesdílí. To by ovšem znamenalo, že robustní žvýkací aparát se u těchto homininů vyvinul nezávisle na sobě, paralelním vývojem.[62]
Někteří autoři také kvůli mírným odlišnostem ostatků stále doporučují užívání názvu Paranthropus crassidens pro nálezy ze Swartkrans a Paranthropus robustus pro ostatky z Kromdraai, přičemž předpokládají, že P. crassidens se vyvinul z P. robustus. Většina odborníků však shrnuje nálezy do jediného druhu P. robustus.[63] Tento názor podporují také nové objevy v Drimolen, které rozšířily známou variabilitu ostatků a které svými rozměry i morfologií překlenují rozdíly mezi nálezy ze Swartkrans a Kromdraai.[2]
Odkazy
Reference
- VANČATA, V. Paleoantropologie - přehled fylogeneze člověka a jeho předků. Brno: Akademické nakladatelství CERM, 2003. 212 s. ISBN 80-7204-272-6. S. 18.
- WOOD, B. A.; RICHMOND, B. G. Human evolution: taxonomy and paleobiology. Journal of Anatomy. 2000, roč. 196, s. 19–60.
- CONROY, G. C. Reconstructing human origins. New York: W. W. Norton & company, 1997. 547 s. Dostupné online.
- BROOM, R. The genera and species of the South African fossil ape man. American Journal of Physical Anthropology. 1950, roč. 8, s. 1–13.
- Ostatky hominidů v jižní Africe byly v první polovině 20. století nalézány především v důsledku intenzivní těžby vápence, který se získával z výplně starých krasových jeskyní.
- BROOM, R.; SCHEPERS, G. W. H. The South-African ape-men: the Australopithecineae. 1. vyd. Pretoria: Transvaal Museum, 1946.
- KEITH, A. Australopithecines or Dartians. Nature. 1947, roč. 159, s. 377.
- LE GROS CLARK, W. Observation on the anatomy of the fossil Australopithecinae. Journal of Anatomy. 1947, roč. 83, s. 300–333.
- BROOM, R. Another new type of fossil ape-man. Nature. 1949, roč. 163, s. 57.
- ROBINSON, J. T. The genera and species of the Australopithecinae. American Journal of Physical Anthropology. 1954, roč. 12, s. 181–200.
- STRAIT, D. S.; GRINE, F. E.; MONIZ, M. A. A reappraisal of early hominid phylogeny. Jounal of Human Evolution. 1997, roč. 32, s. 17–82.
- GRINE, F. E. New craniodental fossils of Paranthropus from the Swartkrans Formation and their significance in ‘‘robust’’ australopithecine evolution. In: GRINE, F. E. Evolutionary History of the ‘‘Robust’’ Australopithecines. [s.l.]: [s.n.], 1988. S. 223–243.
- MENTER, C. G., KUYKENDALL, K. L.; KEYSER, A. W.; CONROY, G. C. First record of hominid teeth from the Plio-Pleistocene site of Gondolin, South Africa. Journal of Human Evolution. 1999, roč. 37, s. 299–307.
- KEYSER, A. W., MENTER, C. G.; MOGI-CECCHI, J.; PICKERING, T. R.; BERGER, L. R. Drimolen: A new hominid-bearing site in Guateng, South Africa. South African Journal of Science. 2000, roč. 96, s. 193–197. Dostupné online.
- KEYSER, A. W. The Drimolen skull: the most complete australopithecine cranium and mandible to date. South African Journal of Science. 2000, roč. 96, s. 189–193. Dostupné online.
- BERGER, L. R., DE RUITER, D. J.; STEININGER, C. M.; HANCOX, J. Preliminary results of excavations at the newly discovered Coopers D deposit, Gauteng, South Africa. South African Journal of Science. 2003, roč. 99, s. 276–278.
- SHAW, J. C. M. Further remains of a Sterkfontein ape. Nature. 1939, roč. 143, s. 117.
- BERGER, L. R.; PICKFORD, M.; THACKERAY, J. F. A Plio-Pleistocene hominid upper central incisor from the Cooper’s site, South Africa. South African Journal of Science. 1995, roč. 91, s. 541–542. Dostupné online.
- KIMBLE, W. H. The species and diversity of australopiths. In: HENKE, W. C.; TATTERSALL, I. Handbook of palaeoanthropology 3. Berlín: Springer, 2007. S. 1539–1573.
- BOAZ, N. T. Paleoecology of early Hominidae in Africa. Kroeber Anthropological Society Papers. 1977, roč. 50, s. 37–62.
- THACKERAY, J. F.; KIRSCHVINK, J. L.; RAUB, T. D. Palaeomagnetic analysis of calcified deposits from the Plio-Pleistocene hominid site of Kromdraai, South Africa. South African Journal of Science. 2002, roč. 98, s. 537–540.
- VOGEL, J. C. Further attempts at dating the Taung tufas. In: TOBIAS, P. V. Ancestors: The Hard Evidence. [s.l.]: [s.n.], 1985. S. 189–194.
- HERRIES, A. I. R.; CURNOE, D.; ADAMS, J. W. A multi-disciplinary seriation of early Homo and Paranthropus bearing palaeocaves in southern Africa. Quaternary International. 2009, roč. 202, s. 14–3 28.
- KIMBEL, W. H. The species and diversity of australopiths. In: HENKE, W.; TATTERSALL, I. Handbook of Palaeoanthropology 3. Berlín: Springer, 2007. S. 1539–1573.
- HERRIES, A. I. R.; ADAMS, J. W. Clarifying the context, dating and age range of the Gondolin hominins and Paranthropus in South Africa. Journal of Human Evolution. 2013, roč. 65, s. 676–681.
- KLEIN, R. G. The causes of “robust” australopithecine extinction. In: GRINE, F. E. Evolutionary History of the “Robust” Australopithecines. [s.l.]: Aldine transactions, 2007. S. 499–505.
- PICKERING, R., KRAMERS, J.D.; HANCOX, P. J.; DE RUITER, D. J.; WOODHEAD, J. D. Synchronous flowstone development links early hominin bearing cave deposits in South Africa. Earth and Planetary Science Letters. 2013, roč. 306, s. 23–32.
- DE RUITER, D. J., PICKERING, R.; STEININGER, C. M.; KRAMERS, J. D.; HANCOX, P. J.; CHURCHILL, S. E. et al. New Australopithecus robustus fossils and associated U-Pb dates from Cooper's Cave (Gauteng, South Africa). Journal of Human Evolution. 2009, roč. 56, s. 497–513.
- CARTMILL, M.; SMITH, F. H. The Human Lineage. 2. vyd. Oxford: Wiley-Blackwell, 2011.
- MCHENRY, H. M. Petite bodies of the ‘‘robust’’ australopithecines. American Journal of Physical Anthropology. 1991, roč. 86, s. 445–454.
- PICKERING, T. R., HEATON, J. L.; CLARKE, R. J.; SUTTON, M. B.; BRAIN, C. K.; KUMAN, K. New hominid fossils from Member 1 of the Swartkrans formation, South Africa. Journal of Human Evolution. 2012, roč. 62, s. 618–628.
- GRINE, F. E., JACOBS, R. L.; REED, K. E.; PLAVCAN, J. M. The enigmatic molar from Gondolin, South Africa: implications for Paranthropus paleobiology. Journal of Human Evolution. 2012, roč. 63, s. 597–609.
- DESOUSA, A.; CUNHA, E. Hominins and the emergence of the modern human brain. In: HOFMAN, M. A.; FALK, D. Evolution of the primate brain. [s.l.]: Elsevier, 2012. S. 293–322.
- HARCOURT-SMITH, W. E. The Origins of Bipedal Locomotion. In: HENKE, W. C.; TATTERSALL, I. Handbook of palaeoanthropology 3. Berlín: Springer, 2007. S. 1483–1518.
- COLLARD, M.; WOOD, B. Grades among the African Early Hominids. In: BROMAGE, T. G.; SCHRENK, F. African Biogeography. Climate Change & Human Evolution. Oxford: Oxford University Press, 1999. S. 316–327.
- LOCKWOOD, C. A., MENTER, C. G.; MOGI-CECCHI, J., KEYER, A. W. Extended male growth in a fossil hominin species. Science. 2007, roč. 318, s. 1443–1446.
- MOGGI-CECCHI, J. The life histories of fossil hominins. Journal of Anthropological Sciences. 2009, roč. 87, s. 3–5.
- COPELAND, S. R., SPONHEIMER, M.; DE RUITER, D. J.; LEE-THORP, J. A.; CODRON, D.; LE ROUX, P. J.; GRIMES, V.; RICHARDS, M. P. Strontium isotope evidence for landscape use by early hominins. Nature. 2011, roč. 474, s. 76–78.
- STEWART, K. M. Environmental change and hominin exploitation of C4-based resources in wetland/savanna mosaics. Journal of Human Evolution. 2014, roč. 77, s. 1–16.
- REED, K. E. Early hominid evolution and ecological change through the African Plio-Pleistocene. Journal of Human Evolution. 1997, roč. 32, s. 289–322.
- WOOD, B. A.; STRAIT, D. S. Patterns of resource use in early Homo and Paranthropus. Journal of Human Evolution. 2004, roč. 46, s. 119–162.
- DE RUITER, D. J.; SPONHEIMER, M.; LEE-THORP, J. A. Indications of habitat association of Australopithecus robustus in the Bloubank Valley, South Africa. Journal of Human Evolution. 2008, roč. 55, s. 1015–1030.
- LEE-THORP, J. A.; VAN DER MERWE, N. J.; BRAIN, C. K. Diet of Australopithecus robustus at Swartkrans from stable carbon isotopic analysis. Journal of Human Evolution. 1994, roč. 27, s. 361–372.
- SZALAY, F. S. Hunting-scavenging protohominids: a model for hominid origins. Man. 1975, roč. 10, s. 420–429.
- SILLEN, A. Strontium-calcium ratios (Sr/Ca) of Australopithecus robustus and associated fauna from Swartkrans. Journal of Human Evolution. 1992, roč. 23, s. 495–516.
- LEE-THORP, J. A.; SPONHEIMER, M. Contributions of biogeochemistry to understanding hominin dietary ecology. Yearbook of physical anthropology. 2006, roč. 49, s. 131–148.
- SPONHEIMER, M., DE RUITER, D. J., LEE-THORP, J. A., SPÄTH, A. Sr/Ca and early hominin diets revisited: new data from modern and fossil tooth enamel. Journal of Human Evolution. 2005, roč. 48, s. 147–156.
- UNGAR, P. S.; SPONHEIMER, M. J. The diets of early hominins. Science. 2011, roč. 334, s. 190–193.
- MACHO, G. A.; SHIMIZU, D. Dietary adaptations of South African australopiths: inference from enamel prism attitude. Journal of Human Evolution. 2009, roč. 57, s. 241–247.
- SPONHEIMER, M., LEE-THORP, J., DE RUITER, D. J., CODRON, D., CODRON, J., BAUGH, A., THACKERAY, J. F. Hominins, sedges, and termites: new carbon isotope data from the Sterkfontein Valley and Kruger National Park. Journal of Human Evolution. 2005, roč. 48, s. 301–312.
- YEAKEL, J. D., BENNETT, N. C., KOCH, P. L., DOMINY, N. J. The isotopic ecology of African mole rats informs hypotheses on the evolution of human diet. Proceedings of the Royal Society of London B. 2007, roč. 274, s. 1723–1730.
- Při analýzách izotopů uhlíku nelze odlišit přímou konzumaci C4-rostlin od konzumace živočichů, kteří se těmito rostlinami živí.
- STRAIT, D. S. The evolutionary history of the australopiths. Evolution Education Outreach. 2010, roč. 3, s. 341–352.
- BACKWELL, L. R.; D'ERRICO, F. Early hominid bone tools from Drimolen. Journal of Archaeological Science. 2008, roč. 35, s. 2880–2894.
- SUSMAN, R. L. New postcranial remains from Swartkrans and their bearing on the functional morphology and behavior of Paranthropus robustus. In: GRINE, F. E. Evolutionary History of the Robust Australopithecines. [s.l.]: [s.n.], 1988. S. 149–172.
- To znamená, že Paranthropus mohl pevně spojit ukazovák a palec bříšky k sobě, což mu umožnilo obratně manipulovat i s drobnými předměty
- CROMPTON, R. H.; VEREECKE, E. E.; THORPE, S. K. S. Locomotion and posture from the common hominoid ancestor to fully modern hominins with special reference to the common panin/hominin ancestor. Journal of Anatomy. 2008, roč. 212, s. 501–543.
- CONSTANTINO, P.; WOOD, B. A. Paranthropus paleobiology. Miscelánea en homenaje a Emiliano Aguirre. 2004, roč. 3, s. 136–151.
- WOOD, B. A.; CONSTANTINO, P. Paranthropus boisei: fifty years of evidence and analysis. American journal of physical anthropology. 2007, roč. 134/S45, s. 106–132.
- VILLMOARE, B.; KIMBEL, W. H. CT-based study of internal structure of the anterior pillar in extinct hominins and its implications for the phylogeny of robust Australopithecus. Proceedings of the National Academy of Sciences. 2011, roč. 108, s. 16 200 – 16 205.
- COLLARD, M.; WOOD, B. Homoplasy and the early hominid masticatory system: inferences from analyses of extant hominoids and papionins. Journal of Human Evolution. 2001, roč. 41, s. 167–194.
- ALEMSEGED, Z.; COPPENS, Y.; GERAADS, D. Hominid cranium from Omo: description and taxonomy of Omo-323-1976-896. American Journal of Physical Anthropology. 2002, roč. 117, s. 103–112. Dostupné online.
- WOOD, B. A.; CONSTANTINO, P. Human origins: Life at the top of the tree. In: CRACRAFT, J.; DONOGHUE, M. Assembling the tree of life. Oxford: Oxford University Press, 2004. S. 517–535.
Literatura
- ROBERTSOVÁ, A. Evoluce: Příběh člověka. Praha: Knižní klub, 2012. 256 s.
- SVOBODA, J. Čas lovců: Aktualizované dějiny paleolitu. Brno: Akademické nakladatelství CERM, 2009. 298 s.
- SVOBODA, J. Předkové. Evoluce člověka. Praha: Academia, 2014. 480 s.
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu Paranthropus robustus na Wikimedia Commons
Obrázky, zvuky či videa k tématu Paranthropus robustus na Wikimedia Commons - Trojrozměrný model lebky druhu Paranthropus robustus