Escherichia coli
Escherichia coli (původním názvem Bacterium coli) je gramnegativní, fakultativně anaerobní, spory netvořící tyčinkovitá bakterie, pohybující se pomocí bičíků. Náleží do čeledi Enterobacteriaceae, jež také zahrnuje mnoho rodů patogenních mikroorganismů.
 | |
| Vědecká klasifikace | |
| Doména | bakterie (Bacteria) |
| Kmen | Proteobacteria |
| Třída | Gammaproteobacteria |
| Řád | Enterobacterales |
| Čeleď | Enterobacteriaceae |
| Rod | Escherichia |
| Binomické jméno | |
| Escherichia coli Theodor Escherich, 1885 | |
| Některá data mohou pocházet z datové položky. | |
E. coli patří ke střevní mikroflóře teplokrevných živočichů včetně člověka, proto je její přítomnost v pitné vodě indikátorem fekálního znečištění. Člověku je jako součást přirozené mikroflóry prospěšná, neboť produkuje řadu látek, které brání rozšíření patogenních bakterií (koliciny), a podílí se i na tvorbě některých vitamínů (např. vitamín K). Existují však i patogenní kmeny tohoto druhu, které mohou způsobit nebezpečná onemocnění.
E. coli objevil německo-rakouský pediatr a bakteriolog Theodor Escherich v roce 1885[1]. Patří k nejlépe prostudovaným mikroorganismům, neboť je modelovým organismem pro genové a klinické studie. Joshua Lederberg jako první r. 1947 pozoroval a popsal na bakterii E. coli výměnu genetického materiálu, tzv. konjugaci[2].
Popis E. coli
Morfologická charakteristika
E. coli je nesporotvorná tyčinka, jež se pohybuje pomocí bičíku. Bakterie dosahuje délky 2–3 μm a šířky 0,6 μm. Některé typy mohou tvořit slizovité obaly, složené z polysacharidů. Na svém povrchu nese dva typy fimbrií. První typ fimbrií se skládá z kyselého hydrofobního proteinu fimbrinu. Umožňuje bakterii přichytit se na epitel hostitele a následně jej kolonizovat. Kolonizaci usnadňuje velký počet fimbrií prvního typu na buňku (100-1000 ks/buňka). Tyto fimbrie jsou vysoce antigenní, neboť obsahují tzv. F antigeny. Druhým typem fimbrií jsou tzv. sex pili, jež hrají důležitou úlohu při konjugaci.
Bakterie se pohybuje pomocí bičíků. Ty jsou složeny z flagelinu (na lysin bohatý protein). Bičíky jsou stejně jako fimbrie vysoce antigenní, a to díky tzv. H antigenům. Na povrchu bakterie se při stresových podmínkách mohou také tvořit polysacharidové kapsule, jež obsahují tzv. K a M antigeny.
Vnější membrána je pokryta lipopolysacharidem a skládá se z lipidové dvojvrstvy, v níž je ukotveno množství membránových proteinů. K proteinům, které tvoří póry, patří poriny Omp C, Omp F a Pho E. Poriny slouží jako vstupní a výstupní kanály pro buněčné metabolity a pro příjem vitaminů z okolí.
Prostor mezi vnější membránou a buněčnou stěnou se nazývá periplazmatický. Vyskytují se zde např. proteiny vážící aminokyseliny či cukry a enzymy degradující antibiotika (beta-laktamasy).
Buněčná stěna E. coli (jakožto zástupce gramnegativních bakterií) se skládá z tenké vrstvy peptidoglykanu, jenž zajišťuje rigidní tvar buňky. Pod vrstvou peptidoglykanu se nalézá cytoplazmatická membrána. Ta se skládá především z proteinů (70 %), lipopolysacharidů a fosfolipidů. Je v ní lokalizováno mnoho biochemických pochodů, např. dýchací řetězec a syntéza ATP.
Cytoplazma bakteriální buňky je viskózní vodný roztok, jenž obsahuje rozpuštěné anorganické a organické látky. Nachází se zde množství ribozomů (cca 40 % hmotnosti celé buňky), díky nimž jsou proteosyntéza a dělení bakteriálních buněk velice rychlé. Při optimálních podmínkách (37 °C, dostatek živin) je doba generace zhruba 20 min. Bakteriální ribozomy jsou menší než eukaryotní. Mají sedimentační konstantu 70S.
Nachází se zde také molekula bakteriální DNA, v níž je uložena veškerá dědičná informace bakterie. Velikost DNA u E. coli K-12 je zhruba 4 700 kbp a kóduje cca 4 400 proteinů.[3] Cytoplazma bakterií na rozdíl od cytoplazmy eukaryot neobsahuje membránové organely.
Fyziologická charakteristika
E. coli je fakultativní anaerob, tzn. využívá respiračního i kvasného metabolismu (fermentace) pro dodávku energie. E. coli jako chemoheterotrof je schopna využívat množství cukrů i aminokyselin jako zdroj uhlíku, nejrychleji však roste na glukóze. Za anaerobních podmínek E. coli využívá glukózu za vzniku laktátu, sukcinátu, acetátu i ethanolu. Za aerobních podmínek je glukóza využita efektivněji a konečným produktem je především oxid uhličitý. E. coli produkuje indol, avšak neroste na citrátu a neprodukuje sirovodík. Je kataláza-pozitivní, oxidáza-negativní. Těchto vlastností se využívá při její identifikaci pomocí tzv. enterotestu[4]. E. coli je schopna růst za teploty 8 °C–48 °C, avšak optimální teplota je 37 °C. Rozsah pH pro růst je pH6–pH8[3].
Antigenní charakteristika
E. coli můžeme taxonomicky dělit dle antigenních struktur na sérotypy. K hlavním strukturám patří somatické O antigeny (lipopolysacharid), jichž je 170 typů, a kapsulární K antigeny (80 typů); dalšími strukturami jsou např. H antigeny (flagelární proteiny) a F antigeny (bílkoviny fimbrií)[3].
Patogenita E. coli
E. coli je jednou z nejčastěji se vyskytujících bakterií v klinických vzorcích. Její přítomnost je u člověka fyziologická pouze ve střevech jako součást střevní mikroflóry. Může se však patologicky vyskytovat i v krevních vzorcích a být tak původcem bakterémie. Často způsobuje nosokomiální infekce, tj. infekce získané v nemocnicích. Bakteriální infekce se léčí podáváním antibiotik. V dnešní době však nalézáme stále větší počet kmenů bakterií, které jsou k podávaným antibiotikům rezistentní.
Patogenní kmeny způsobují dva typy onemocnění. Prvním je extraintestiální onemocnění, při němž jsou napadeny především močové cesty, dochází k infekci ran a jejich hnisání. Pokud se bakterie dostane do intestinálního traktu člověka, vyvolává infekce provázené průjmy. Extraintestiální formy onemocnění jsou vyvolávány kmeny, jež mají polysacharidový kapsulární K antigen, příp. P fimbrie, jimiž adherují na povrch sliznic. Pokud se bakterie E. coli dostane do zažívacího traktu, mluví se o ní jako o enteropatogenním kmenu E. coli. Ty lze dále rozlišit:
Enteropatogenní (EPEC)
EPEC E. coli vyvolává vodnaté průjmy především u novorozenců. Může dojít k vysokému stupni dehydratace a následnému úmrtí. Toto onemocnění je stále problémem v rozvojových zemích. Frekvenci infekce způsobené EPEC E. coli u dospělých jedinců je složité stanovit, jelikož u pacientů starších tří let se po příčině nezávažného průjmu nepátrá.
Enterotoxigenní (ETEC)
_from_Escherichia_coli_O157-H7_PDB_1r4p.png.webp)
ETEC E. coli též vyvolává průjmové stavy jak u dětí, tak i u dospělých. Tento kmen se vyskytuje v teplých oblastech (Egypt, Bangladéš); do střední Evropy se může dostat s cestovateli. ETEC se vyznačuje tvorbou dvou typů enterotoxinů – termolabilního (TL) a termostabilního (TS). Informace pro tvorbu těchto toxinů je uložena na bakteriálních plazmidech.
TL enterotoxin se skládá z polypeptidové podjednotky A o molekulární hmotnosti 25 kDa a pěti podjednotek B o molekulární hmotnosti 11,5 kDa. Podjednotka B se váže na epitelové buňky a napomáhá translokaci podjednotky A do buňky. Ta katalyzuje aktivaci cAMP. Enzymatickou kaskádou dochází ke zvýšené sekreci sodíku z buňky, a tím i k úniku chloridů a následně i vody. Následkem jsou vodnaté průjmy. ST enterotoxin je slabě imunogenní. Aktivuje guanylátcyklázu a tím se zvyšuje koncentrace cGMP. Další mechanismus není přesně znám. Je možné, že určitou úlohu hraje vápník[5].
Enteroinvazivní (EIEC)
EIEC E. coli pronikají do buněk tlustého střeva, kde se množí. Toto onemocnění připomíná průběh bacilární dysenterie.
O epidemiologii EIEC není mnoho známo. Neexistuje její lidský ani zvířecí rezervoár. V oblastech s nedostatečnou hygienou způsobuje EIEC až 5 % všech průjmů. Nejčastější sérologickou skupinou je O124.
Enterohemoragické (EHEC)
Typ EHEC E. coli, stejně jako EPEC, je schopen adheze na stěny endotelu. Na rozdíl od EPEC E. coli se však EHEC váže na endotelie tlustého střeva a produkuje zde toxin, tzv. shigella toxin, též zvaný verotoxin. Ten způsobuje poškození sliznice tlustého střeva, což vede ke krvavým průjmům. Onemocnění, jež tento kmen způsobuje, se nazývá hemoragická kolitida. U některých pacientů může dojít k poškození ledvin a onemocnění přechází do hemolyticko-uremického syndromu (HUS), jenž bývá smrtelný. Zdrojem infekce je infikované hovězí maso. Identifikace EHEC E. coli se opírá o unikátní neschopnost utilizovat sorbitol[6]. Jedním ze zástupců kmene EHEC je sérotyp E. coli O157:H7.
Významné sérotypy E. coli
E. coli nissle
Poprvé jej izoloval v roce 1917 na východní frontě profesor Alfred Nissle. Už on usoudil, že přítomnost tohoto sérotypu ve střevní mikroflóře výrazně snižuje riziko vzniku ulcerózní kolitidy. V nedávné době byl tento objev znovu experimentálně potvrzován, přičemž se ukázalo, že ostatní sérotypy tuto schopnost nemají, a dokonce jejich přítomnost ve střevě může snižovat účinnost E. coli nissle. V současnosti se mikrobiologové snaží vyvinout léčbu ulcerózní kolitidy pomocí osazení střeva pacienta E. coli nissle.
E. coli O157:H7
Sérotyp E. coli O157:H7 je z hlediska veřejného zdraví nejdůležitějším zástupcem E. coli EHEC, neboť způsobuje HUS. Rezervoárem tohoto patogenu je především dobytek (přežvýkavci). Na člověka se přenáší kontaminovanými potravinami (tepelně neupravené maso a mléko), ale i při fekálním znečištění vody a křížovou kontaminací při přípravě pokrmů. Infekční dávka je velice nízká (jednotky bakterií na gram potravy). Poprvé byl tento sérotyp izolován r. 1982[7].
E. coli K-12
Tento sérotyp byl poprvé izolován r. 1922 na univerzitě ve Stanfordu (USA) z lidské stolice. E. coli K-12 je nejprobádanějším zástupcem E. coli vůbec. Příčinou je jednak přítomnost lysogenního bakteriofága lambda v bakterii, jednak široká vybavenost E. coli K-12 množstvím plazmidů, což dává široké možnosti pro využití v genovém inženýrství.
Sérotyp E. coli K-12 byl použit např. při výzkumu metabolismu dusíku u bakterií, biosyntéze L-tryptofanu z indolu a L-serinu a také při studiích konjugace bakterií.[8]
Léčba infekcí způsobených E. coli
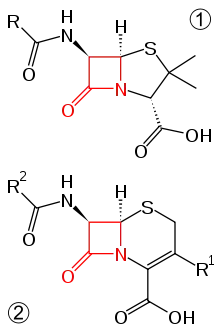
Bakteriální infekce jsou léčeny antibiotiky (beta-laktamová antibiotika, fluorochinolony a aminoglykosidy). Jelikož E. coli má krátkou generační dobu, může se u ní stejně jako u ostatních bakterií velice rychle vyvinout rezistence k podávanému antibiotiku.
Rezistence k beta-laktamovým antibiotikům
Mechanismus účinku beta-laktamových antibiotik (penicilin, ampicilin, cefalosporiny) je založen na skutečnosti, že beta-laktamový kruh antibiotika naruší syntézu buněčné stěny bakterie a ta, jelikož není chráněna, zahyne.
E. coli může produkovat enzym beta-laktamázu, jež hydrolyzuje beta-laktamový kruh antibiotika a tím se stává imunní k působení tohoto antibiotika. Geny pro tuto rezistenci bývají uloženy na plazmidu, a proto je genetická informace velice snadno přístupna k předávání jiným bakteriím (i mezidruhově). Nevýhodou informace uložené na plazmidu je skutečnost, že nemusí při dělení buňky dojít k jejímu předání oběma dceřiným buňkám. Rezistentní by byla pouze jedna z nově vzniklých buněk, druhá by rezistenci nenesla.
Dnes je známo, že v České republice je E. coli cca ze 60 % případů rezistentní k podávání aminopenicilinů a z 15 % k cefalosporinům[9].
Rezistence k fluorochinolonům
Fluorochinolony jsou antibiotika, jež inhibují replikaci bakteriální DNA. Pokud si bakterie na ně vyvine rezistenci, může se jednat o dva různé mechanismy. Nejčastějším je tzv. eflux – tj. poté, co se antibiotikum dostane do bakteriální buňky, je pomocí membránových pump vyčerpáno mimo buňku.
Druhým způsobem, jak se bakterie brání působení fluorochinolonů, je pozměnění (methylace, adenylace) jejich cílových struktur, tedy enzymů, jež jsou odpovědné za replikaci bakteriální DNA. Tyto enzymy se nazývají DNA gyráza a topoisomeráza IV. V České republice se rezistence kmenů E. coli k fluorochinolonům pohybuje okolo 25 % případů, což je zhruba evropský průměr.[9].
Rezistence k aminoglykosidům
Aminoglykosidy jsou antibiotika, která blokují proces translace, tj. přepis genetické informace z řeči nukleotidů do řeči aminokyselin. Enzymy, kde translace probíhá, se nazývají ribozomy. Rezistence k těmto antibiotikům spočívá v modifikaci ribozomů (methylace, adenylace) tak, že se stanou pro aminoglykosidová antibiotika biologicky inertní.
V České republice se výskyt E. coli rezistentní k aminoglykosidům pohybuje okolo 10 % případů, což je v rámci Evropy stále průměrné číslo[9].
Význam
Bakterie E. coli se vyskytuje jako součást přirozené mikroflóry teplokrevných živočichů v tlustém střevě a dolní části tenkého střeva. Člověk je touto bakterií kolonizován už od narození (kontaminace z potravy, přenos z již kolonizovaného jedince, nejčastěji matky). E. coli patří k nejlépe prostudovaným známým bakteriím, což je důvodem jejího využití v biotechnologiích a genovém inženýrství. Mezi její klady jakožto zástupce prokaryot patří rychlý růst a levná kultivační média, naopak nevýhodou použití E. coli může být její neschopnost posttranslačních modifikací, např. glykosylace. V takových případech se v průmyslu využívá kvasinka Saccharomyces cerevisiae[3].
Genové inženýrství
E. coli je nejběžněji používanou bakterií v metodách molekulární biologie. Její kultivace je úspěšná v médiích bohatých (kromě zdroje uhlíku, dusíku a dalších esenciálních látek tato média obsahují i aminokyseliny a vitaminy) i minimálních (obsahují pouze glukózu jako zdroj uhlíku a energie a soli jako zdroj síry, fosforu apod.). Pro klonování se používají speciálně upravené kmeny E. coli, které jsou zbaveny schopnosti syntézy restrikčních endonukleáz. Pro účely molekulární genetiky byla připravena řada mutantů E. coli:
- DH5 – kmen zbavený schopnosti rekombinace, tj. opravy DNA, používaný pro amplifikaci plazmidové DNA
- BL21 – kmen schopný vysoké exprese svých genů díky promotoru bakteriofága T7, jenž je integrován do chromozómu bakterie E. coli[10]
Lidský inzulin

Diabetes mellitus I. stupně je onemocnění, při němž slinivka břišní není schopna produkovat inzulin. Léčba je substituční, kdy se pacientovi exogenně podává inzulin. Dříve byl získáván z jatečných prasat. Na takto získaný inzulin však docházelo k alergickým reakcím. Příčinou byla různá lokace glykosylace řetězců inzulinu prasat a u člověka. R. 1982 byl na trh uveden inzulin získaný rekombinačně pomocí geneticky modifikované bakterie E. coli pod záštitou firmy Eli Lilly. Inzulin se také získává rekombinantními metodami i pomocí eukaryotních organismů, např. Saccharomyces cerevisiae (pod komerčním názvem Novokin od firmy NovoNordisk)[11].
Výroba indiga
Roku 1983 byla objevena schopnost bakterie E. coli syntetizovat textilní barvivo indigo. Jeho produkce byla však pro průmyslové účely malá a náklady vysoké (jako výchozí substrát byl používán indol, příp. aminokyselina L-tryptofan), proto byla pomocí rekombinace získána E. coli, jež by mohla jako výchozí substrát používat glukózu. Významným kladem této biotechnologické výroby s využitím modifikované E. coli oproti syntetické výrobě je ekologická nezávadnost, jelikož při syntetické výrobě mohou vznikat toxické meziprodukty[12].
Další produkty
K dalším produktům rekombinantní E. coli patří např. příprava lidského růstového hormonu, interferonu alfa, interferonu gama, interleukinu-2 či produkce aminokyselin (L-valin, L-threonin). Pod záštitou firmy Eli Lilly se na trh s léčivy dostaly rekombinantní hormony, jako jsou parathormon a kalcitonin, jež chrání před osteoporózou). Od r. 1999 je na trhu v USA vakcína proti lymské borelióze s názvem Lymerix. Opět se jedná o rekombinantní protein připravený z E. coli. Tato vakcína je účinná pouze na bakterii Borrelia burgdorferi, jež je majoritním původcem lymské boreliózy v USA (v Evropě je to nejčastěji Borrelia garinii)[13].
Odkazy
Reference
- Koliformní bakterie a E. coli (anglicky)
- Objev procesu konjugace u E. coli (anglicky)
- Escherichia coli (anglicky)
- Šilhánková L., Mikrobiologie pro potravináře a biotechnology, ACADEMIA, 2002. ISBN 80-200-1024-6.
- Greenwood D., Slack R., Peutherer J., Lékařská mikrobiologie, Grada, 1999.
- Bednář M., Fraňková V., Schindler J., Souček A., Vávra J., Lékařská mikrobiologie, Marvil, 1996.
- E. coli O157: H7, FDA (anglicky)
- E. coli K-12, J. Lederberg (anglicky). www.sgm.ac.uk [online]. [cit. 2011-10-01]. Dostupné v archivu pořízeném dne 2011-10-25.
- E. coli a resistence k antibiotikům (anglicky)[nedostupný zdroj]
- Ruml T., Rumlová M., Pačes V. Genové inženýrství, VŠCHT, 2002. ISBN 978-80-7080-499-5.
- E. coli jako producent insulinu (anglicky)
- E. coli jako producent textilního barviva (česky)
- Fusek M., Káš J, Ruml T. Bioléčiva, VŠCHT, 2008. ISBN 978-80-7080-678-4.
Literatura
- Lee, Sang Yup. Systems Biology and Biotechnology of Escherichia coli, Springer, 2009. ISBN 978-1-4020-9393-7.
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu Escherichia coli na Wikimedia Commons
Obrázky, zvuky či videa k tématu Escherichia coli na Wikimedia Commons - E. coli a její využití v terapeutické léčbě (anglicky)
- Infekční strategie E. coli (video)
- Epidemie E. coli EHEC O104: H4
- Epidemie E. coli EHEC-Státní zdravotní ústav
- Taxonomie E. coli O104: H4 (anglicky)
- Taxonomie E. coli O157: H7 (anglicky)