Sinice
Sinice (Cyanobacteria, ale také Cyanophyta či Cyanoprokaryota[2]) je kmen nebo oddělení (záleží, zda se jedná o bakteriologické či botanické pojetí) gramnegativních bakterií. Vyznačují se schopností fotosyntézy, při níž vzniká kyslík (tzv. oxygenní typ). Český název této skupiny pochází ze slova siný, tedy tmavě modrý.
 | |
| Vědecká klasifikace | |
| Doména | bakterie (Bacteria) |
| Kmen | sinice (Cyanobacteria) Stanier ex Cavalier-Smith, 2002 |
| Třída | Cyanophyceae Schaffner, 1909 |
| Řády[1] | |
| Některá data mohou pocházet z datové položky. | |
_bei_Gy%C3%B6rs%C3%B6v%C3%A9nyh%C3%A1z.jpg.webp)
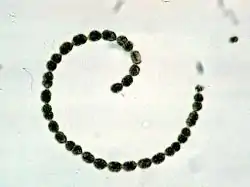
Sinice jsou jednobuněčné, zpravidla však žijí v koloniích tvaru kokálních shluků či vláknitých stélek (hormogonií). Nejčastěji jsou modrozeleně zbarvené. Jejich buňky jsou v mnohých ohledech typicky prokaryotické: obsahují kruhovou molekulu DNA, bakteriální typ ribozomů a chybí u nich složitější membránové struktury. Fotosyntetická barviva se nachází ve speciálních útvarech, fykobilizomech nebo thylakoidech. K hlavním pigmentům účastnícím se fotosyntézy patří chlorofyl (zpravidla typu a, někdy též b, c nebo d) a dále allofykocyanin, fykocyanin, fykoerythrin a další. Sinice se rozmnožují nepohlavně, a to buněčným dělením či fragmentací vláken (vegetativně).
Vyskytují se velmi hojně ve vodním prostředí, ale i v půdě a mnohdy také v extrémních podmínkách, jako jsou pouště či polární oblasti. Velmi často také vstupují do symbiotických vztahů. Vyjma endosymbioticky vzniklých plastidů je možné se setkat s mnoha případy, kdy sinice pomáhají svému hostiteli fixovat dusík či uhlík.
Vznik a vývoj

Sinice se pravděpodobně vyvinuly z anaerobních fotosyntetizujících bakterií, jako jsou dnešní purpurové bakterie či chlorobakterie (Chloroflexi).[3]
Nejstarší známé důkazy o existenci sinic na Zemi v podobě fosílií jsou staré 3,5 miliardy let a pochází především z formace Apex Chert v Austrálii.[4] Tyto prekambrijské nálezy tzv. stromatolitů možná představují vůbec nejstarší nálezy buněčných organismů.[5] Některé studie však tvrdí, že tyto nálezy jsou abiotického původu a představují jen složité struktury vzniklé devitrifikací horniny na bázi uhlíku.[6] Přibližně před 2,5 miliardami let se zintenzivnil vulkanismus a uvolněný oxid uhličitý způsobil rozmach sinic.[7] Lépe zachované fosilní nálezy sinic pochází z období mezi 2–0,9 miliardami let. Vzhledem jsou identické s dnešními sinicemi a pokud je nějaká fosílie zařazena do určitého současného rodu, před daný rodový název se často dává předpona „paleo“. Udává se, že tyto sinice patří do řádů Chroococcales a Oscillatoriales.[4] Až do počátku kambria (před cca 600 miliony lety) byly sinice dominantními organismy na Zemi, a tento dlouhý úsek geologické historie Země je proto označován jako „věk sinic“. V tomto období se také díky sinicím postupně zvyšoval obsah kyslíku v atmosféře.[4] Přes dlouhou evoluční historii sinic se zdá, že se vzhled jejich buněk téměř nezměnil, i když ke genetickým změnám dochází. Tato teorie může být označena jako stagnující evoluce.[4]
Plastidy, organely mnohých eukaryotických organismů (především různé řasy a rostliny), v mnohých ohledech připomínají sinice. Podle tzv. endosymbiotické teorie, která je podpořena různými strukturálními a genetickými podobnostmi, se plastidy vyvinuly z sinic pohlcených eukaryotními buňkami[8] asi před 1,5 miliardami lety.[9][10]
Stavba buněk
Sinice se vyskytují jako jednotlivé buňky, shluky buněk v koloniích nebo jako vlákna. Jejich stélka se proto obvykle označuje buď jako kokální či trichální.[4] Některé vláknité formy mívají specializované buňky heterocyty (či též heterocysty), ve kterých probíhá fixace vzdušného dusíku, případně akinety, což jsou klidové buňky určené k přetrvání nepříznivých období. Buňka sinic je prokaryotického typu a obvykle dosahuje velikosti 1–10 mikrometrů,[3] jen vzácně více (např. buňka Chroococcus giganteus má až 60 mikrometrů).[11] Pouhým okem je možné spatřit kolonie sinic. Sinice a jejich kolonie jsou zbarveny nejčastěji modrozeleně, ale dosahují barev od blankytně modré přes malachitově zelenou, žlutou, červenou až po černou.[11]
Struktury na povrchu buňky
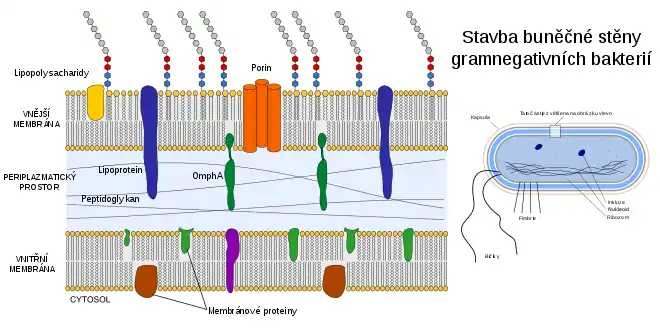
Sinice jsou do jisté míry gramnegativní bakterie a jako takové Gramovou metodou získávají růžové zabarvení, protože tyto bakterie mají specifický typ buněčné stěny. Sinice jako gramnegativní bakterie mají totiž na povrchu dvě plazmatické membrány (vnitřní a vnější) a mezi nimi poměrně tenkou vrstvu peptidoglykanu (murein), jenž tvoří pevnou složku celé buněčné stěny.[4] Na druhou stranu některé specifické vlastnosti buněčné stěny sinic sinice odlišují od ostatních gramnegativních bakterií, a sinice se tak zdají být směsí vlastností gramnegativní a grampozitivní buňky. Zatímco běžné gramnegativní bakterie mají buněčnou stěnu o šířce jen asi 2–6 nanometrů, u jednobuněčných sinic je to asi 10 nanometrů, u vláknitých sinic 15–35 nanometrů a vzácně až 700 nanometrů (u sinice Oscillatoria princeps). Po chemické stránce jsou ale peptidy vytvářející zmíněná příčná spojení spíše charakteristická pro gramnegativní bakterie. Rovněž kyselina teichoová, typická složka stěn grampozitivních bakterií, u sinic chybí.[12] Membrána obsahuje mnohé transportní kanály umožňující pasivní či aktivní transport látek přes stěnu. Příkladem jsou pasivní kanály z proteinu porinu či různé specifické kanály určené pro pomalu difundující látky. U některých vláknitých sinic byly nalezeny v příčných přehrádkách mezi jednotlivými buňkami zvláštní kanály připomínající plazmodezmy rostlin.[4]
Směrem ven od vnější plazmatické membrány navíc mají sinice zpravidla slizovou vrstvu (glykokalyx) složenou z lipopolysacharidů. Obecně mívá fibrilární (vláknitou) strukturu. Někdy bývá vyvinuta více a jindy méně a v určitých případech může tvořit silnou homogenní či vrstevnatě uspořádanou pochvu kolem celé buňky (a tento obal je někdy ještě navíc zbarvený).[4]
U sinic nikdy nebyly nalezeny bičíky,[12] přesto se ale mnohé z nich (zejména zástupci řádu Oscillatoriales) dokáží po povrchu aktivně pohybovat. Při pohybu je produkováno velké množství slizu, skutečný hnací motor však představují svazky stažitelných bílkovinných vláken umístěných na povrchu buněk. Obvykle se klouzají, někdy však projevují i zvláštní typ rotačního pohybu (který dal jméno například českému rodu Oscillatoria, drkalka).[4]
Protoplast
V protoplastu (vnitřním prostoru) sinic není v porovnání s eukaryotickými buňkami takové množství organel. Obvykle se rozlišují dva typy cytoplazmy: na povrchu buněk bývá výrazně barevná chromatoplazma, obsahující velké množství fotosyntetických barviv, zatímco uvnitř je centroplazma neboli centroplazmatická oblast, kde převládá sinicová DNA, ribozomy a podobně. Ribozomy sinic jsou prokaryotního typu: jejich malá podjednotka obsahuje 16S RNA, zatímco velká podjednotka 23S RNA.[4]
DNA vytváří jedinou kruhovitou molekulu DNA, nazývanou nukleoid, která neobsahuje histony.[pozn 1] Velikost genomu se pohybuje od asi 1,7 milionu párů bází (Prochlorococcus[14][15]) až po asi 8,9 milionu párů bází (Nostoc punctiforme[16]). Zajímavá je skutečnost, že sinice mohou mít více kopií své genetické informace, tzn. nemusí být haploidní. Dodatečný genetický materiál mohou také představovat plazmidy, malé kruhové molekuly DNA. Sinice jsou schopné předávání částí své genetické informace z jedné buňky do druhé (tzv. horizontální genový transfer).[4]
Další buněčné struktury se vyvinuly u sinic v souvislosti s jejich fotosyntetickým způsobem výživy. Významné jsou zejména thylakoidy, membránou obalené měchýřky uložené podél cytoplazmatické membrány či prorůstající skrz naskrz celou buňkou. Fotosyntéze mimoto pomáhají i tzv. fykobilizomy. Některé vodní sinice tvoří i válcovité měchýřky o délce až 1000 nanometrů a naplněné vzduchem, které umožňují vznášení buněk ve vodním sloupci.
Klasifikace


Je známo velké množství taxonů sinic, ale přesný počet nelze zjistit. Kalina a Váňa[4] například uvádí 150 rodů s 2000 druhy s tím, že je tento počet pravděpodobně silně podhodnocený. Někdy se díky značné proměnlivosti (fenoplasticitě) sinic vůbec nepřistupuje k popisování druhů, ale užívají se jen rody.[4] Pro jednotlivé morfologické odchylky v rámci rodu se pak používá termín morfotyp.[11] V historii byly sinice považovány za nižší rostliny. V angličtině na to dosud upomíná termín blue-green algae, český historický název sinné řasy[3] a rovněž latinský název Cyanophyceae. Na druhou stranu dnešní název „sinice“ je neutrální, pochází ze slova sinný, tedy modrý.[17][18]
Při určování sinic se nejprve hledí na typ stélky, tvar a velikost buněk, případně navíc uspořádání buněk v koloniích a přítomnost slizu. U vláknitých kolonií se navíc zjišťuje přítomnost slizové pochvy, způsob propojení buněk, tvar vlákna a případně typ jeho větvení. Důležitá je také přítomnost heterocytů a akinet.[11]
Když se díky mikroskopickým a biochemických studiím ukázalo, že jsou sinice jednou z podskupin bakterií, získaly nejen botanické, ale i druhé, bakteriologické názvosloví. Botanické názvosloví ustanovuje tzv. Mezinárodní kód botanické nomenklatury (ICBN), zatímco klasifikaci bakterií se zabývá Mezinárodní bakteriologický kód (ICNB).[19] Na přelomu sedmdesátých a osmdesátých let dvacátého století se přešlo z botanického na bakteriologické názvosloví, ale dosud bylo pod platným bakteriologickým názvoslovím publikováno jen velmi málo druhů sinic.[20] Další významnou změnou je skutečnost, že jsou dřívější prochlorofyty považovány za sinice, přestože mají některé specifické vlastnosti.[21]
Klasifikace sinic je stále v pohybu.[20] Thomas Cavalier-Smith zařadil v roce 2002 oddělení sinic do velké skupiny Glycobacteria a naznačil jejich příbuznost zejména s kmeny Firmicutes a Actinobacteria. Dále dělí sinice na skupiny Gloeobacteria (pro rod Gloeobacter, neobsahující thylakoidy) a Phycobacteria (ostatní sinice včetně prochlorofytů). Druhou jmenovanou skupinu Cavalier-Smith rozdělil na pět obvyklých řádů, jež jsou uznávány i botaniky:[1]
- Chroobacteria
- Chroococcales (včetně prochlorofytů)
- Pleurocapsales (někdy řazena do Chroococcales)
- Oscillatoriales
- Hormogoneae
Systém NCBI prakticky přejímá Cavalier-Smithovo uspořádání, ale s jedním rozdílem. Zde figuruje skupina Prochlorales jako samostatný řád.[22] Naopak velmi odlišné je pojetí systému sinic v publikaci Bergey's Manual of Systematic Bacteriology. V této knize je rozdělen kmen sinice na pět sekcí (I–V) bez taxonomického označení,[23][24] které v určitých ohledech korespondují se systémem Cavalier-Smithe.[25] Zmíněný systém pěti sekcí vyjmenovává jen rody sinic, druhy podle autora u sinic nelze rozlišit.[4] V literatuře se přiklání k tomuto typu klasifikace i další autoři.[26] Tak či onak, molekulárně biologické studie některé navrhované taxony nepotvrdily a označují je za nepřirozené z hlediska vývoje života na Zemi.[25]
Metabolismus
Fotosyntéza
Z metabolického hlediska jsou sinice fototrofní a autotrofní (podobně jako většina rostlin), tedy zkráceně fotoautotrofové. Vyznačují se především fotosyntézou oxygenního typu, při níž je voda donorem elektronů, oxid uhličitý je fixován na organické sloučeniny a jako vedlejší produkt se uvolňuje kyslík. Tato reakce se dá zjednodušeně vyjádřit rovnicí:
- 6 CO2 + 12 H2O → C6H12O6 + 6 O2 + 6 H2O
Centrem fotosyntetických reakcí jsou u sinic tzv. thylakoidy (vyjma primitivní sinice Gloeobacter, která je nemá), tyto struktury totiž obsahují vlastní fotosyntetická barviva, která jsou nutná pro přeměnu světelné energie na chemickou. Přehled fotosyntetických barviv u sinic je skutečně pestrý. U sinic se vyskytují všechny čtyři známé druhy chlorofylu, tedy nejen a (který má zpravidla roli hlavního fotosyntetického pigmentu), ale u některých zástupců navíc i b, c, nebo d. Chlorofyl b byl nalezen u prochlorofytů (Prochloron, Prochlorococcus, Prochlorothrix), chlorofyl c u některých zástupců rodu Prochlorococcus a chlorofyl d se zdá být hlavním fotosyntetickým pigmentem u sinice Acaryochloris marina. Ve fykobilizomech bývají obsaženy tři fykobiliproteiny: modrý allofykocyanin a fykocyanin a červený fykoerytrin. V některých případech byla nalezena i barviva β-karoten, zeaxantin, echinenon, kantaxantin či myxoxantofyl (typ xanthofylu), mnohé z nich nemusí zastávat roli v procesu fotosyntézy.[4]

Dále jsou v tylakoidech přítomna reakční centra – tzv. fotosystémy I a II. Ve fotosystému II probíhá fotolýza vody za vzniku kyslíku a vodíkového kationtu, který spolu s uvolněnými elektrony vstupuje do dalších reakcí. Elektrony jsou transportovány řetězcem proteinových komplexů a nakonec vedou k vytvoření protonového gradientu napříč membránou, který je podstatou vzniku adenosintrifosfátu (ATP). Mimo tylakoidy se uplatňují při fotosyntéze ještě další struktury, jako například fykobilizomy, které umožňují využít co nejširší spektrum světla. Karboxyzomy jsou tělíska obsahující enzym Rubisco, který umožňuje syntézu cukrů v Calvinově cyklu. Zásobní látkou je u sinic především tzv. sinicový škrob, ale dusík se obvykle skladuje ve formě cyanofycinu a fosfor ve formě zrn volutinu.[4]
Fotosyntetická schopnost sinic je v mnoha ohledech poměrně flexibilní. Konkrétním příkladem je chromatická adaptace probíhající ve ztížených světelných podmínkách. Jedná se o jev, při němž se na fykobilizomech upravuje počet fykocyaninových a fykoerytrinových jednotek a tím se posouvá spektrum využitelného světla. Pokusy bylo dokázáno, že buňka je schopna změnit svou barvu od ocelově šedé přes různé stupně zelené po červenavou. Druhým příkladem přizpůsobivosti je schopnost přejít v anaerobním prostředí s množstvím sirovodíku na anaerobní fotosyntézu, při níž je jako donor elektronů užíván místo vody právě sirovodík.[4]
Evoluční původ oxygenní fotosyntézy je nadále předmětem zkoumání. Nové teorie ukazují, že společný předek sinic jí nebyl schopen; geny pro ni se pravděpodobně vyvinuly až poměrně pozdě v evoluční historii sinic (přibližně před 2,3 miliardami let) v linii vedoucí k recentní třídě Oxyphotobacteria a odtud se dále šířily horizontálním genovým přenosem a fúzemi fotosystémů.[27][28]
Úloha v koloběhu dusíku
Sinice jsou také významnými fixátory vzdušného dusíku v celosvětovém měřítku a významně tak ovlivňují koloběh dusíku v přírodě. Zpravidla se reakce odehrává ve speciálních buňkách bez fotosyntetické funkce, zvaných heterocyty či heterocysty. V striktně anaerobních podmínkách uvnitř heterocytů se pomocí enzymu nitrogenázy ze vzdušného dusíku (N2) vytváří za spotřeby adenosintrifosfátu (ATP) amonné sloučeniny. Některé sinice ani nemají heterocyty (např. rod Lyngbya), ty pak fixují dusík v noci, kdy v buňkách neprobíhá fotosyntéza a množství kyslíku uvnitř buňky je tedy minimální.[4]
Sekundární metabolismus
Sinice produkují množství dalších sekundárních metabolitů a podpůrných látek, například různé oligosacharidy, karboxylové kyseliny, vitamíny, peptidy, atraktanty, hormony, enzymy, antibiotika, polysacharidy a toxiny.[3] Na ochranu proti UV záření např. produkují pigment scytonemin, případně i gloeocapsin, β-karoten, kantaxantin či myxoxantofyl. Všechny tyto látky odfiltrovávají záření o velmi nízké vlnové délce. Sinice rovněž produkují široké spektrum jedů, souhrnně tzv. cyanotoxinů. Způsobují kožní alergie, zánět spojivek, bronchitidu, u dobytka napájeného znečištěnou vodou i otravu. Nebezpečné koncentrace dosahují sinicové jedy především v době, kdy je ve vodě rozvinutý tzv. „vodní květ“.[4]
Rozmnožování
U sinic může být pozorováno pouze nepohlavní rozmnožování, žádné rozmnožovací buňky u sinic neexistují.[11] Buňky jednobuněčných (kokálních) sinic se množí pouze prostým dělením. Toto dělení začíná tvorbou příčné přehrádky, která vzniká od krajů buňky a postupně se uzavírá (podobně jako clona fotoaparátu). Pokud dělení probíhá ve více rovinách, vznikají kolonie s různou vzájemnou orientací buněk. Některé sinice se však tomuto klasickému scénáři vyhýbají: Chamaesiphon se rozmnožuje pomocí exocytů, tedy spor vznikajících na volném konci buňky, a Chroococcidiopsis se rozmnožuje mnohonásobným dělením tzv. baeocytu. Vláknité sinice se mohou štěpit na dílčí pohyblivá vlákna, tzv. hormogonie.[4] K dlouhodobému přežívání slouží u některých sinic akinety, zvláštní druh tlustostěnných spor.[11]
Ekologie
Sinice za svůj evoluční úspěch a velmi dlouhou geologickou historii vděčí kombinací několika pro ně charakteristických vlastností. V prekambrickém období pravděpodobně byly schopny sinice tolerovat nízký obsah kyslíku, vysokou míru UV záření a zvýšené koncentrace sirovodíku.[29] I dnes jsou sinice téměř všudypřítomné a mnohdy také vstupují do významných symbiotických svazků. Pokud se přemnoží ve vodním prostředí, jsou nazývány „vodní květ“.
Výskyt

Sinice se vyskytují v širokém spektru různých prostředí: v sladkých i slaných vodách (plankton), na povrchu půdy a v tenké vrstvě pod ní, na skalách (například vápencových) i uvnitř jeskyní, ale nápadná je schopnost sinic osídlovat různá extrémní prostředí, která jsou pro jiné skupiny organismů nehostinná, a z tohoto důvodu v nich sinice také často zcela dominují.[29] Sinice jsou například velmi často primárními kolonizátory dosud neosídlených skal či nově vzniklých půd.[30] Byly zaznamenány termofilní sinice rodu Synechococcus, žijící v termálních pramenech v teplotě až 73 °C. Ve špatných světelných a nutričních podmínkách je schopna žít řada sinic podílejících se na tvorbě mořského pikoplanktonu. Tzv. halofilní druhy jsou schopné odolávat vysokým koncentracím soli v okolní vodě, například v Mrtvém moři. Alkalofilní zástupci dokáží žít i ve velmi zásaditém prostředí, při pH 13,5 žije sinice rodu Leptolyngbya (zřejmě nejvyšší zjištěná hodnota pH, v níž byl objeven život). V suchozemském prostředí úspěšně vzdorují vyschnutí tzv. xerofilní sinice: v poušti Negev se vyskytují sinice v drobných dutinkách v půdě, kde se nachází stopové množství vody. V polárních oblastech jsou sinice velmi důležitou složkou zdejšího koloběhu živin, kolonizují například dna antarktických jezer a ledovou krustu v Grónsku.[4][29] Některé další sinice osídlují póry uvnitř kamenů.[11]
Sinice se podílí na vzniku povlaků inkrustovaných uhličitanem vápenatým, z nichž mohou vznikat různé travertiny, onkolity a stromatolity.[31]
Vodní květ
Sinice jsou častým původcem jevu známého jako vodní květ, při němž dochází k přemnožení některých vodních fotosyntetizujících organismů nad určitou míru. Problém představuje především vodní květ v sladkovodních nádržích, které jsou využívány jako zdroj pitné vody a k rekreaci. Při rozvoji vodního květu jsou ve větším množství do vody uvolňovány cyanotoxiny. Rizikové pro rozvoj sinicového vodního květu jsou zejména vodní plochy s vyšší hodnotou pH, vyšší teplotou vody a vysokým obsahem živin (především fosforu). Mnohdy vodní květ způsobuje nadměrné vypouštění živin do vody, tedy tzv. eutrofizace vod.[32] Existují různé způsoby boje proti vodnímu květu.
Symbiotické interakce

Sinice vstupují velmi často do symbiotických svazků s jinými organismy, a to zejména do mutualistických (oboustranně prospěšných) vztahů, kde sinice zpravidla hraje roli fotobionta. V některých případech vstupují sinice přímo do cizích eukaryotních buněk. Pozůstatkem minimálně jedné takové události jsou plastidy – organely mnoha eukaryot připomínající v mnohém své prapředky, sinice. Jinou, ale podobnou, událostí vznikly primitivní plastidy u prvoků rodu Paulinella, které mají dodnes patrnou peptidoglykanovou stěnu. Tyto mezičlánky na cestě mezi endosymbiontem a organelou se často nazývají cyanely. Poněkud známějším příkladem symbiózy sinic jsou lišejníky (konkrétně cyanolišejníky), které asi v 8–15 % obsahují sinice, například rodu Nostoc či Calothrix. Nostoc žije v symbiotickém svazku také s houbou Geosiphon pyriforme, sinice Richelia uvnitř některých rozsivek (Diatomeae) a jiný rod zase uvnitř buněk obrněnek (Dinoflagellata). Také rostliny mohou hostit sinice: Nostoc žije v krytosemenných rostlinách rodu Gunnera, ale i v kořenech mnohých cykasů.[4] Jiné sinice žijí uvnitř těl kapradinek rodu Azola.[33] I uvnitř těl mořských hub (Porifera) a u pláštěnců (Tunicata) mohou být přítomny fotosyntetizující sinice, ty pak jsou nazývány zoocyanely.[34]
Viry sinic
U sinic jsou známy i jejich virové patogeny, tzv. cyanofágy, jejichž genetickým materiálem je zásadně dvouvláknová DNA. Známými viry jsou např. sladkovodní as-1 a lpp-1. Viry významně ovlivňují početnost sinic, některé průzkumy například objevily viry v téměř všech studovaných populací sinic.[4] Ekologické výzkumy v oceánu také ukazují, že u pobřeží mají sinice rodu Synechococcus daleko vyšší rezistenci vůči svým virům, protože se s nimi na mělčině častěji setkávají a přírodní výběr zde tedy působí daleko silněji.[29]
Význam pro člověka
Sinice mají do jisté míry nezanedbatelný hospodářský význam. Nepočítaje negativní vliv vodního květu na ekonomiku a lidské zdraví, mají sinice mnoho kladných stránek. Sinice především obsahují v sušině vysoké koncentrace proteinů (až 70 %), mnohem více než například zelené řasy.[11] Sinice rodu Arthrospira (známá pod komerčním názvem Spirulina) se pěstuje v mnoha zemích na výrobu vitamínových tablet. Obsahuje mimo jiné ve velké míře vitamíny (vitamín B12) a karoteny.[4] Především v nezápadních civilizacích byly sinice dříve nedílnou součástí jídelníčku[35] a v některých případech (Čad) se k přípravě pokrmů užívají dosud.[36]
Některé pigmenty sinic (fykobiliny), zejména fykocyanin, se používají jako netoxická barviva, díky nimž se pozorují metabolické procesy probíhající v buňkách různých organismů.[4] Jiné látky obsažené v sinicích by se mohly do budoucna stát výchozí surovinou pro výrobu protirakovinných a protizánětlivých léků, antibiotik a antivirotik.[35] Kosmické agentury NASA a ESA uvažují o zapojení sinic do stravy kosmonautů na budoucích vesmírných misích na velkou vzdálenost,[37][38] některé sinice jsou schopné růst i na měsíční půdě.[18] Uvažuje se o užití sinic pro výrobu biopaliv.[39]
Odkazy
Poznámky
Reference
- CAVALIER-SMITH, T. The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification. International Journal of Systematic and Evolutionary Microbiology. 2002-01, roč. 52, čís. Pt 1, s. 7–76. Dostupné v archivu pořízeném dne 2008-07-24. ISSN 1466-5026. DOI 11837318. Archivováno 24. 7. 2008 na Wayback Machine
- Biolib – Cyanobacteria (sinice) [online]. Dostupné online.
- MARŠÁLEK, Blahoslav. Sinice jako producenti biologicky aktivních látek. Živa. 2002, čís. 5.
- KALINA, Tomáš; VÁŇA, Jiří. Sinice, řasy, houby, mechorosty a podobné organismy v současné biologii. Praha: Karolinum, 2005. ISBN 80-246-1036-1. S. 606.
- Schopf JW, Kudryavtsev AB, Agresti DG, Wdowiak TJ, Czaja AD. Laser-Raman imagery of Earth's earliest fossils. Nature. March 2002, roč. 416, čís. 6876, s. 73–6. DOI 10.1038/416073a. PMID 11882894.
- BRASIER, Martin, Owen Green, John Lindsay, Andrew Steele. Earth's oldest (approximately 3.5 Ga) fossils and the 'Early Eden hypothesis': questioning the evidence. Origins of Life and Evolution of the Biosphere: The Journal of the International Society for the Study of the Origin of Life. 2004-02, roč. 34, čís. 1–2, s. 257–69. Dostupné online [cit. 2008-10-21]. ISSN 0169-6149. DOI 14979661.
- https://phys.org/news/2019-12-mysteries-linked-breathable-oxygen.html - New study points to one cause for several mysteries linked to breathable oxygen
- McFadden G. Endosymbiosis and evolution of the plant cell. Curr Opin Plant Biol. 1999, roč. 2, čís. 6, s. 513–9. (anglicky)
- HEDGES, S Blair, Jaime E Blair, Maria L Venturi, Jason L Shoe. A molecular timescale of eukaryote evolution and the rise of complex multicellular life. BMC Evolutionary Biology. 2004-01-28, roč. 4, s. 2. Dostupné online [cit. 2008-10-21]. ISSN 1471-2148. DOI PMC341452.
- YOON, Hwan Su, Jeremiah D Hackett, Claudia Ciniglia, Gabriele Pinto, Debashish Bhattacharya. A molecular timeline for the origin of photosynthetic eukaryotes. Molecular Biology and Evolution. 2004-05, roč. 21, čís. 5, s. 809–18. Dostupné online [cit. 2008-10-21]. ISSN 0737-4038. DOI 14963099.
- ŠEJNOHOVÁ, Lenka; MARŠÁLEK, Blahoslav. Pohled do mikroskopického světa sinic. Živa. 2005, čís. 3.
- HOICZYK, Egbert, Alfred Hansel. Cyanobacterial Cell Walls: News from an Unusual Prokaryotic Envelope. Journal of Bacteriology. 2000-03, roč. 182, čís. 5, s. 1191–1199. Dostupné online [cit. 2008-10-24].
- WELSH, Eric A., Michelle Liberton, Jana Stöckel, Thomas Loh, Thanura Elvitigala, Chunyan Wang, Aye Wollam, Robert S. Fulton, Sandra W. Clifton, Jon M. Jacobs, Rajeev Aurora, Bijoy K. Ghosh, Louis A. Sherman, Richard D. Smith, Richard K. Wilson, Himadri B. Pakrasi. The genome of Cyanothece 51142, a unicellular diazotrophic cyanobacterium important in the marine nitrogen cycle. Proceedings of the National Academy of Sciences of the United States of America. 2008-09-30, roč. 105, čís. 34, s. 15094–15099. Dostupné online. DOI 10.1073/PNAS.0805418105.
- G. Rocap, et al.. Genome divergence in two Prochlorococcus ecotypes reflects oceanic niche differentiation. Nature. 2003, roč. 424, s. 1042–1047. DOI 10.1038/nature01947.
- A. Dufresne, et al.. Genome sequence of the cyanobacterium Prochlorococcus marinus SS120, a nearly minimal oxyphototrophic genome.. Proc. Natl Acad. Sci. USA. 2003, roč. 100, s. 10020–10025. DOI 10.1073/pnas.1733211100. PMID 12917486.
- J.C. Meeks, et al.. An overview of the genome of Nostoc punctiforme, a multicellular, symbiotic cyanobacterium. Photosynth. Res.. 2001, roč. 70, s. 85–106.
- Sinice. Toxicita sinic vodního květu [online].
- HROMADOVÁ, Miroslava. Pozemské sinice mohou růst v měsíční půdě [online]. Dostupné online.
- KOMÁREK, Jiří; GOLUBIĆ, Stjepko. Guide to the nomenclature and formal taxonomic treatment of oxyphototroph prokaryotes (Cyanoprokaryotes) (PROPOSAL) [online]. 2004 [cit. 2008-10-26]. Dostupné v archivu pořízeném dne 2010-08-21.
- OREN, Aharon. A proposal for further integration of the cyanobacteria under the Bacteriological Code. Int J Syst Evol Microbiol. 2004-09-01, roč. 54, čís. 5, s. 1895–1902. Dostupné online [cit. 2008-10-26]. DOI 10.1099/ijs.0.03008-0.
- LEWIN, Ralph A. Prochlorophyta – a matter of class distinctions. Photosynthesis Research. 2002, roč. 73, čís. 1–3, s. 59–61. Dostupné online [cit. 2008-10-20]. ISSN 0166-8595. DOI 10.1023/A:1020400327040.
- Taxonomy Browser (Cyanobacteria) [online]. NCBI. Dostupné online.
- G. Garrity, D. R. Boone, and R. W. Castenholz. Bergey's Manual of Systematic Bacteriology. 2. vyd. New York: [s.n.], 2001. Kapitola Phylum BX. Cyanobacteria. Oxygenic Photosynthetic Bacteria: The Archaea and the Deeply Branching and Phototropic Bacteria.
- Cyanobacteria – Tree of Life Web Project [online]. Dostupné online.
- FLORES, Enrique; HERRERO, Antonia. The Cyanobacteria: Molecular Biology, Genomics and Evolution. [s.l.]: Horizon, 2008. Dostupné online. ISBN 1904455158. (anglicky)
- The Cyanobacteria and Their Taxonomy [online]. Dostupné online.
- SOO, Rochelle M.; HEMP, James; PARKS, Donovan H.; FISCHER, Woodward W.; HUGENHOLTZ, Philip. On the origins of oxygenic photosynthesis and aerobic respiration in Cyanobacteria. S. 1436–1440. Science [online]. American Association for the Advancement of Science, 31. březen 2017. Svazek 355, čís. 6332, s. 1436–1440. Dostupné online. ISSN 1095-9203. DOI 10.1126/science.aal3794. (anglicky)
- How do plants make oxygen? Ask cyanobacteria
- WHITTON, Brian A.; POTTS, Malcolm. The Ecology of Cyanobacteria. Their Diversity in Time and Space. [s.l.]: Kluwer Academic Publ., 2000. Dostupné online. ISBN 0792347358 9780792347354. (anglicky)
- GARCIA-PICHEL, Ferran, Richard W Castenholz. Occurrence of UV-Absorbing, Mycosporine-Like Compounds among Cyanobacterial Isolates and an Estimate of Their Screening Capacity. Applied and Environmental Microbiology. 1993-01, roč. 59, čís. 1, s. 163–169. Dostupné online. ISSN 1098-5336. DOI PMC202072.
- PETRÁNEK, Jan. Řasy – geologická encyklopedie [online]. Dostupné online.
- ŘÍHOVÁ AMBROŽOVÁ, Jana. Encyklopedie hydrobiologie: Vodní květ [online]. Dostupné online.
- BAKER, Judith A, Barrie Entsch, David B McKay. The cyanobiont in an Azolla fern is neither Anabaena nor Nostoc. FEMS Microbiology Letters. 2003-12-05, roč. 229, čís. 1, s. 43–7. Dostupné online [cit. 2008-11-30]. ISSN 0378-1097. DOI 14659541.
- ČEPIČKA, Ivan; KOLÁŘ, Filip; SYNEK, Petr. Mutualismus, vzájemně prospěšná symbióza; Přípravný text – biologická olympiáda 2007–2008. Praha: NIDM ČR, 2007. S. 87.
- GANTAR, Miroslav, Zorica Svirčev. MICROALGAE AND CYANOBACTERIA: FOOD FOR THOUGHT. Journal of Phycology. 2008, roč. 44, čís. 2, s. 260–268. j.1529-8817.2008.00469.x Dostupné online. DOI 10.1111/j.1529-8817.2008.00469.x. (anglicky)
- ABDULQADER, Gatugel, Laura Barsanti, Mario R. Tredici. Harvest of Arthrospira platensis from Lake Kossorom (Chad) and its household usage among the Kanembu. Journal of Applied Phycology. 2000-10-01, roč. 12, čís. 3, s. 493–498. Dostupné online. DOI 10.1023/A:1008177925799. (anglicky)
- Normal, Al. Characterization of Spirulina biomass for CELSS diet potential. [s.l.]: Alabama A&M University, 1998.
- Cornet J.F., Dubertret G. The cyanobacterium Spirulina in the photosynthetic compartment of the MELISSA artificial ecosystem. Marseille, France: Workshop on artificial ecological systems, DARA-CNES, 1990.
- PAZDERA, Jozef. Mikrob stvořený pro produkci biopaliv [online]. Dostupné online.
Literatura
česky:
- KALINA, Tomáš; VÁŇA, Jiří. Sinice, řasy, houby, mechorosty a podobné organismy v současné biologii. Praha: Karolinum, 2005. 606 s. ISBN 80-246-1036-1.
- HINDÁK, František. Fotografický atlas mikroskopických siníc. 1. vyd. Bratislava: Veda, 2001. ISBN 80-224-0658-9. S. 127.
- MARŠÁLEK, Blahoslav; KERŠNER, V; MARVAN, V. Vodní květy sinic. Brno: Nadatio flos-aquae, 1996. S. 142.
- ZNACHOR, Petr. Vodní květy řas a sinic. Scientific American české vydání. Červenec 2005, s. 42–51. Dostupné online.
- ŠEJNOHOVÁ, Lenka; MARŠÁLEK, Blahoslav. Pohled do mikroskopického světa sinic. Živa. 2005, čís. 3.
- BABICA, Pavel; MARŠÁLEK, Blahoslav, Bláha, L.,Drábková,M. Toxiny sinic – zbraň proti konkurenci?. Živa. 2004, čís. 4.
- MARŠÁLEK, Blahoslav. Sinice jako producenti biologicky aktivních látek. Živa. 2002, čís. 5.
anglicky:
- FLORES, Enrique; HERRERO, Antonia. The Cyanobacteria: Molecular Biology, Genomics and Evolution. [s.l.]: Horizon, 2008. Dostupné online. ISBN 1904455158. (anglicky)
- WHITTON, Brian A.; POTTS, Malcolm. The Ecology of Cyanobacteria. Their Diversity in Time and Space. [s.l.]: Kluwer Academic Publ., 2000. Dostupné online. ISBN 0792347358. (anglicky)
Externí odkazy
 Obrázky, zvuky či videa k tématu sinice na Wikimedia Commons
Obrázky, zvuky či videa k tématu sinice na Wikimedia Commons  Slovníkové heslo sinice ve Wikislovníku
Slovníkové heslo sinice ve Wikislovníku Taxon Cyanobacteria ve Wikidruzích
Taxon Cyanobacteria ve Wikidruzích- Seznam děl v Souborném katalogu ČR, jejichž tématem jsou sinice
- (česky) Studijní materiály o sinicích na katedře botaniky PřF UK
- (česky) Stránky věnované studiu řas a sinic – Jihočeská univerzita v Českých Budějovicích; podrobné informace o sinicích, určovací klíče, návody, galerie Archivováno 9. 4. 2009 na Wayback Machine
- (česky) Centrum pro cyanobakterie a jejich toxiny – studijní a popularizační materiály o sinicích
- (česky) Centrum pro cyanobakterie a jejich toxiny – sinice a koupání v české přírodě
- (česky) Laboratoř ekologie fykoplanktonu AV ČR: Galerie mikrofotografií mnoha druhů sinic Archivováno 11. 1. 2012 na Wayback Machine
- (česky) Encyklopedie hydrobiologie – sinice; přehled základních informací o sinicích
- (slovensky) Informácie o siniciach – rodové rozdelenie, obrázky
- (anglicky) Cyanosite – rozsáhlá galerie fotografií, rozcestník stránek o sinicích na internetu
- (anglicky) The on-line database of cyanobacterial genera; stránky věnované klasifikaci sinic