Strunatci
Strunatci (Chordata) jsou kmen druhoústých živočichů, který zahrnuje obratlovce a několik blízce příbuzných skupin bezobratlých. Jsou dvoustranně souměrní, v některém období svého života mají strunu hřbetní, nervovou trubici, žaberní štěrbiny, endostyl a svalnatou ocasní část až za řitním otvorem. Přitom platí, že nervová trubice je na hřbetní straně, struna hřbetní probíhá centrálně a srdce je umístěno na břišní straně.
Stratigrafický výskyt: spodní kambrium–současnost[1] | |
|---|---|
 Tetra průsvitná (Pristella maxillaris) | |
| Vědecká klasifikace | |
| (nekategorizováno) | Opisthokonta |
| Říše | živočichové (Animalia) |
| (nekategorizováno) | druhoústí (Deuterostoma) |
| Kmen | strunatci (Chordata) Bateson, 1885 |
| Podkmeny a třídy | |
| |
| Některá data mohou pocházet z datové položky. | |
Strunatci se vyvinuli pravděpodobně na začátku kambria[1] (asi před 530 miliony lety). Zatím známe asi 45 000 druhů.[2] Kmen strunatců se dělí na tři podkmeny: pláštěnce, bezlebečné a obratlovce, přičemž se zdá, že blíže obratlovcům mají pláštěnci. Ti mají v larválním stádiu hřbetní strunu a nervový provazec, které se pak v dospělosti ztrácí. Zato bezlebeční mají jak hřbetní strunu, tak i nervový provazec, ale bez páteře. U všech obratlovců, s výjimkou sliznatek, je nervový provazec obemknut chrupavčitými nebo kostěnými obratli a hřbetní struna je zásadně redukovaná. Mezi strunatce patří vedle samotného člověka také mnoho obecně známých a hospodářsky významných živočichů řazených mezi savce, ptáky, paprskoploutvé ryby apod.).
Popis
Strunatci jsou mnohobuněční živočichové s dokonale vyvinutým tělem (členěným na orgány), které je dvoustranně souměrné (bilaterálně symetrické). Tělo je tvořeno třemi zárodečnými listy (ektoderm, mezoderm, entoderm). Velikost těla je velmi různorodá a sahá od méně než 1 centimetru (někteří pláštěnci a hlaváčovité ryby) až po desítky metrů (plejtvák obrovský, někteří dinosauři).[3] S ostatními živočichy mají strunatci některé společné rysy, a to zejména v embryonálním stadiu: například střední část trávicí soustavy vzniká z entodermu, zatímco začátek a konec z ektodermu.[1]
Největšími známými zástupci této skupiny jsou obří kytovci (zejména plejtvák obrovský, dosahující hmotnosti přes 170 tun) a druhohorní sauropodní dinosauři, jejichž délka se mohla pohybovat až kolem 40 metrů.[4]
Anatomie
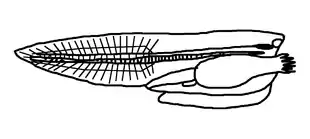
Jako definující se běžně uvádí čtyři nebo pět anatomických znaků přítomných vždy alespoň v embryonálním stadiu:[5][6][7][8]
- Struna hřbetní (chorda dorsalis, též notochord) – opora těla tvořená silně vakuolizovanými[9] buňkami, vzniká z entodernu. Nachází se nad trávicí soustavou a pod nervovou trubicí. Představuje podélnou osu těla směřující od hlavy až k ocasu. U pokročilejších strunatců v dospělosti zaniká a její funkci přejímá chrupavka či kost (páteř).
- Nervová trubice (neurocoel) – trubicovitá nervová soustava na hřbetní straně těla, ektodermálního původu. U mnohých strunatců se v hlavové části z nervové trubice vyvíjí mozek a zbytek je mícha; centrální nervový systém umožňuje poměrně složité smyslové vnímání či dokonalý pohyb.
- Žaberní štěrbiny či alespoň váčky – v hltanu, alespoň během embryonálního vývoje. Procházejí skrze stěnu trávicí trubice a směřují ven z těla. U nižších strunatců slouží mimo výměny plynů i k filtrování potravy. U těch suchozemských jsou silně redukovány a zřejmé jsou spíše jen v embryonální fázi.
- Pravý ocas – nezasahuje do něj trávicí trubice (nachází se celý až za řitním otvorem, je tedy postanální). Je vyztužen chordou či páteří.
- Endostyl (též podhltanová žláza), případně štítná žláza[6] – vyvíjí se jako rýha v břišní stěně hltanu, někdy produkuje hlen umožňující lepší posun potravy dále,[10] navíc představuje úložiště jódu.[5]
Mimo to se uvádí i další rysy, které obvykle platí pro strunatce. Vakovité srdce (tedy stahu schopná céva) se nachází na břišní straně těla[8] a pumpuje krev nejprve směrem k hlavě. U většiny strunatců je cévní soustava spjata s žábrami: odkysličená krev přichází do srdce či do kontraktilních cév, odkud je pumpována do žaber, načež se odvádí aortou do těla. Srdce tedy bylo původně venózní, ale po vzniku plic u suchozemských obratlovců se i toto změnilo a např. u člověka je samozřejmě do něj přiváděna i krev okysličená.[1] Trávicí trubice je trubicovitá a zakládá se v podstatě podobně jako u ostatních živočichů.[6] Světločivné buňky oka (sítnice) se vyvíjí z ektodermu centrální nervové soustavy, nikoliv z epidermis.[11]
Co se týče nejbližších příbuzných strunatců, některé ze znaků uvedených výše jsou v určitých obměnách nalézány i např. u polostrunatců. Mají například něco podobného jako je struna hřbetní – u nich se tomu říká stomochord, vypadá poměrně podobně, ale zřejmě vznikl nezávisle, na základě konvergentní evoluce[10] (jiné zdroje ho však naopak považují za homologii[11]). Podobná je zřejmě situace s neurochordem žaludovců. Naopak jednoduché „žaberní štěrbiny“ polostrunatců jsou zřejmě do určité míry homologické (vývojově spjaté) s pravými žaberními štěrbinami strunatců. Endostyl, další charakteristický orgán strunatců, se zřejmě v pravém slova smyslu u polostrunatců nevyskytuje.[10]
Embryonální vývoj
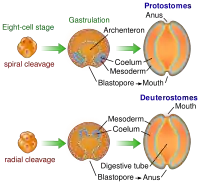
Strunatci patří mezi druhoústé, tedy skupinu živočichů, která je pojmenována právě na základě typu embryonálního vývoje. Konkrétně se pro druhoústé ve většině embryologických učebnic uvádí, že ústa se tvoří až poté, co vznikl řitní otvor, a nevznikají z blastoporu, ale spíše na místě protilehlém. Navíc většina druhoústých tvoří tělní dutinu pomocí váčků, jež se vychlipují z střeva (vzniká tzv. enterocoel). Konečně se uvádí, že druhoústí mají radiální typ rýhování.[1][12][13] Jenže výjimek z těchto pravidel je u strunatců velké množství.[13] Například u obratlovců se nevyskytuje enterocoel, ale naopak, tělní dutina se zakládá jako schizocoel[14][15] (enterocoelie je typický pro část těla kopinatců a mihule[16]). Mezoderm tedy často vzniká z velkých odštěpených bloků mezodermu či ze zakladatelských buněk, jež vcestovaly do prostoru budoucího mezodermu.[16] Navíc mnozí strunatci nevytváří dříve ústa ani řitní otvor – namísto toho v blastoporu vzniká žloutková stopka,[15] pak se uzavírá a řitní otvor se prolamuje nezávisle.[16]
Pro strunatce je typické zakládání některých orgánových soustav v raných fázích embryonálního vývoje. Z dorzální (zádové) stěny střeva se odškrcuje pás mezodermu (chordamezoderm), z něhož vzniká struna hřbetní. Z jiné části mezodermu (stěny coelomu) se vyvíjí somity, jakési články, z nichž vzniká segmentované svalstvo. Z ektodermu se zase vchlipuje neuroektodermální pás buněk, který tvoří nervovou trubici. Žaberní štěrbiny vznikají obvykle z entodermu vychlípením, ale u obratlovců se na jejich vzniku podílí i vchlipující se ektoderm. (protože dochází k perforaci tělní stěny). Co se týče kosterní soustavy, pro strunatce je poměrně typické, že kostra (pokud se vyvíjí) má svůj původ v mezodermu a nikoliv v ektodermu, jak tomu je u mnohých jiných živočichů. Nakonec tak vzniká tělo strunatců složené z charakteristicky vzájemně uspořádaných orgánů: toto uspořádání vyplývá právě z průběhu embryonálního vývoje.[1]
Evoluce
| druhoústí |
| ||||||||||||||||||
Moderní přístupy, které zkoumají příbuznost jednotlivých živočišných kmenů, obvykle považují za nejbližší příbuzné strunatců polostrunatce (Hemichordata) a ostnokožce (Echinodermata). Tyto dva kmeny tvoří společně sesterskou skupinu ke strunatcům. Tato sesterská skupina se označuje Ambulacraria.[17][18][19]
Stáří kmene strunatci se odhaduje s odstupem stovek milionů let poměrně těžko. Jednou z metod, jež to alespoň přibližně umožňují, jsou molekulární hodiny. Podle jedné studie založené na molekulárních hodinách se linie strunatců oddělila od skupiny Ambulacraria zhruba před 896 miliony lety (konkrétně před 832–1022 miliony lety s důvěryhodností 95 %).[20] Jiná, ač starší studie udává výrazně jiná čísla, klade tento mezník do doby před 600 miliony lety, což by více sedělo na paleontologické nálezy.[21] Dawkins zase odhadl stáří společného předka tří současných podkmenů na 565 milionů let.[22]
Evoluční hypotézy
Podle moderních dat o příbuznosti pláštěnců, kopinatců a obratlovců si zřejmě můžeme udělat obrázek, jak vypadal primitivní (rozuměj „evolučně původní“) strunatec. Tento společný předek strunatců měl zřejmě zřetelnou tělní segmentaci, měl ploutevní lem, chordu po celé délce těla a také přímý vývoj. Pláštěnci jsou v mnohých ohledech zřejmě druhotně odvozená skupina, u níž např. vznikl nepřímý vývoj (přes larvu).[16]
Vzniklo několik hypotéz, které se pokouší osvětlit vznik strunatců z druhoústých předků:[23]
- Aurikuláriová (též dipleurová hypotéza): předkem strunatců byla pohyblivá obrvená larva živočicha podobného křídložábrým (Pterobranchia). Z řady brv na zádové straně se postupně vyvinula nervová trubice, od dospělého křídložábrého byly „okoukány“ zase žaberní štěrbiny a chorda. Díky procesu neotenie se z larev těchto živočichů podobných křídložábrým vyvinul jednoduchý pohlavně dospělý strunatec.[23] Jenže tato teorie má i trhliny: proč je například dospělým strunatcům podobnější dospělec polostrunatců více, než larva, z níž se podle této hypotézy strunatci vyvinuli? A proč se u strunatců tak výrazně změnilo dorzoventrální (hřbetobřišní) uspořádání těla?[16]

- Hypotéza inverzní (dorzoventrální inverze): předek strunatců měl nejprve podobné uspořádání orgánů, jako prvoústí – nervová soustava a svalovina na břišní straně (ventrálně), srdce na zádové straně (dorzálně). Tento předek zřejmě již měl nervovou trubici. Mutací tohoto tělního plánu došlo k radikální změně a celé tělo se invertovalo („obrátilo“ dorzoventrálně). Zřejmě k této inverzi nedošlo u polostrunatců, což znamená, že by to byl exkluzivní znak strunatců. Důkazem pro inverzní hypotézu je např. exprese genu bmp – u prvoústých se uplatňuje na zádech, u druhoústých na břišní straně.[23]
- Hypotéza „hemichordate“ (z angl. výrazu pro polostrunatce): strunatci se vyvinuli z živočicha podobného polostrunatcům (již s nervovou trubicí), během evoluce došlo ke zdůraznění a zveličení znaků typických pro polostrunatce. Přispět k tomu podle některých autorů mohla přestavba coelomových váčků a somitů na hlavové i zádové části.[23]
- Hypotéza bilaterálního předka: v podstatě jde o tvrzení, že v evoluci strunatců k ničemu významnému nedošlo. Pohnutkou k tomuto názoru bylo zjištění v devadesátých letech 20. století, že se genom strunatců v zásadních genech vlastně shoduje s genomy velmi odlišných prvoústých živočichů, jako je např. octomilka (Drosophila). Příkladem podobnosti jsou různé Hox geny určující vývoj hlavy a hrudi, pax6 a six geny v očích či např. různé geny určující vývoj nervové tkáně, srdce, střeva a útrobního mezodermu.[23]
Fosilní záznam
Fosilních záznamů prvních strunatců je poskrovnu, je navíc velmi těžké přiřadit více než půl miliardy let starou fosílii k určitému kmeni. Jejich těla postrádají kosti a jiné tvrdé části, a proto špatně fosilizují.[24] K údajným strunatcům patří fosilní živočichové Yunnanozoon (spodní kambrium, Čcheng-ťiang, provincie Jün-nan), Cathaymyrus (totožné naleziště),[1] Haikouella (spodní kambrium, Kchun-ming v Číně[25]) a Pikaia (střední kambrium, burgesské břidlice, Britská Kolumbie).[1]
Systematika


Popis taxonu Chordata (česky strunatci) je přisuzován Williamu Batesonovi (1885),[26] nicméně tohoto názvu se evidentně používalo již před rokem 1885.[27] Klasifikace strunatců procházela složitým vývojem, nicméně již poměrně brzy se ukázalo, že mimo obratlovce se podobnou tělní stavbou vyznačují také pláštěnci a bezlebeční. Do dnešní doby se uznávají tyto tři skupiny (nejčastěji podkmeny) strunatců:[1][26]
- podkmen pláštěnci (Tunicata)
- podkmen bezlebeční (Cephalochordata)
- podkmen obratlovci (Craniata)
Taxon strunatci je s největší pravděpodobností monofyletický, ačkoliv s důkazy pro tuto skutečnost byly poměrně značné problémy.[19] Vzájemné vztahy tří hlavních podkmenů nejsou zcela jisté. Některé (zejména starší) zdroje považují za sesterskou skupinu obratlovců bezlebečné:[3][17][28]
| strunatci |
| ||||||||||||
Mnohé nedávné studie však přichází se zcela novým konceptem, který považuje pláštěnce za příbuznější obratlovcům, než jsou kopinatci.[20][19][29][30] Tyto studie pracují obvykle s rozsáhlejšími daty získaných čtením protein–kódujících genů.[19] Tato novější teorie se tak postupně dostává i do popularizačních knih (Zrzavý 2006[16] ji v podstatě uvádí jako hotovou věc). Rozhodně se však stále objevují i kritické hlasy[31] proti této teorii, která je zobrazena na níže uvedeném fylogenetickém stromu:
| strunatci |
| ||||||||||||
Snaha porozumět vývoji strunatců pomocí morfologických informací (tedy především na základě srovnávací anatomie pláštěnců, kopinatců a obratlovců) je poměrně neúspěšná a nedává odpověď na otázku, zda jsou obratlovcům evolučně blíže pláštěnci nebo kopinatci. Obratlovce a kopinatce spojuje chorda dorsalis po celé délce těla, podobná stavba neurální trubice a smyslových orgánů, ale i třeba jistý ploutevní lem na hřbetě a ocasu či rozdělení těla do velkého počtu segmentů, metamer nebo somitů. Obratlovci a pláštěnci však mají, jak se ukazuje, také mnoho společných rysů. Spojuje je nový typ buněčných spojů (tzv. těsné spoje), podobná stavba chordy, ztráta některých myoepitelů, neuromastové smyslové buňky (základy budoucí postranní čáry obratlovců) a podobně.[16]
Odkazy
Reference
- ROČEK, Zbyněk. Historie obratlovců : evoluce, fylogeneze, systém. Praha: Academia, 2002. 512 s. ISBN 80-200-0858-6.
- T.A. Jefferson, S. Leatherwood, M.A. Webbe. Marine Mammals of the World > Species: Phylum Chordata (Chordates) [online]. eti.uva.nl [cit. 2010-05-14]. Dostupné v archivu pořízeném dne 2007-09-13.
- LUNDBERG, John G. Chordata - Tree of Life Web Project [online]. [cit. 2010-03-16]. Dostupné v archivu pořízeném dne 2007-02-24.
- SOCHA, Vladimír. Nejdelší obratlovec všech dob?. OSEL.cz [online]. 15. listopadu 2018. Dostupné online. (česky)
- RYCHEL, Amanda L., Shannon E. Smith, Heather T. Shimamoto, Billie J. Swalla. Evolution and Development of the Chordates: Collagen and Pharyngeal Cartilage. Mol Biol Evol. 2006-03-01, roč. 23, čís. 3, s. 541–549. Dostupné online [cit. 2010-03-16]. DOI 10.1093/molbev/msj055.
- MILLER, Stephen A; HARLEY, John P. Zoology. 5. vyd. [s.l.]: The McGraw−Hill Companies, 2001.
- KIMBALL, John W. The Vertebrates [online]. [cit. 2010-03-16]. Dostupné v archivu pořízeném dne 2010-03-17.
- ROSYPAL, Stanislav. Nový přehled biologie. [s.l.]: Scientia, 2003. S. 797.
- ROČEK, Zbyněk. Obecná morfologie živočichů; kapitola Pokryv těla a opěrná soustava [online]. Dostupné online.
- Ruppert, E. Key characters uniting hemichordates and chordates: homologies or homoplasies?. Canadian Journal of Zoology. 2005, roč. 83, s. 8–23. Dostupné v archivu pořízeném dne 2012-12-09. DOI 10.1139/Z04-158.
- HAJER, Jaromír. Fylogeneze a systém strunatců. Ústí nad Labem: Univerzita JAP, přírodovědecká fakulta, katedra biologie, 2006. Dostupné v archivu pořízeném dne 2012-01-10. Archivováno 10. 1. 2012 na Wayback Machine
- GILBERT, Scott F. Developmental Biology. 6. vyd. Swarthmore College: Sinauer Associates Dostupné online. ISBN 0-87893-243-7.
- VALENTINE, James W. Cleavage patterns and the topology of the metazoan tree of life. Proceedings of the National Academy of Sciences of the United States of America. 1997-07-22, roč. 94, čís. 15, s. 8001–8005. Dostupné online [cit. 2010-03-22]. DOI VL - 94.
- EAKIN, Richard Marshall. Vertebrate Embryology. [s.l.]: [s.n.], 1949. 144 s. Dostupné online.
- ONKEN, Michael. What are the 'advantages' of developing a deuterostome pattern of embryonic [online]. MadSci Network, 1999. Dostupné online.
- ZRZAVÝ, Jan. Fylogeneze živočišné říše. Praha: Scientia, 2006. 255 s. ISBN 80-86960-08-0.
- CAMERON, C B, J R Garey, B J Swalla. Evolution of the chordate body plan: new insights from phylogenetic analyses of deuterostome phyla. Proceedings of the National Academy of Sciences of the United States of America. 2000-04-25, roč. 97, čís. 9, s. 4469–4474. Dostupné online. ISSN 0027-8424.
- ADOUTTE, A, G Balavoine, N Lartillot, O Lespinet, B Prud'homme, R de Rosa. The new animal phylogeny: reliability and implications. Proceedings of the National Academy of Sciences of the United States of America. 2000-04-25, roč. 97, čís. 9, s. 4453–4456. Dostupné online [cit. 2010-03-18]. ISSN 0027-8424.
- SWALLA, Billie J, Andrew B Smith. Deciphering deuterostome phylogeny: molecular, morphological and palaeontological perspectives. Philosophical Transactions of the Royal Society B: Biological Sciences. 2008-04-27, roč. 363, čís. 1496, s. 1557–1568. ISSN 0962-8436. DOI 10.1098/rstb.2007.2246.
- BLAIR, Jaime E., S. Blair Hedges. Molecular Phylogeny and Divergence Times of Deuterostome Animals. Mol Biol Evol. 2005-11-01, roč. 22, čís. 11, s. 2275–2284. Dostupné online. DOI 10.1093/molbev/msi225.
- AYALA, Francisco José, Andrey Rzhetsky, Francisco J. Ayala. Origin of the metazoan phyla: Molecular clocks confirm paleontological estimates. Proceedings of the National Academy of Sciences of the United States of America. 1998-01-20, roč. 95, čís. 2, s. 606–611. Dostupné online [cit. 2010-03-21]. DOI VL - 95.
- DAWKINS, Richard. Příběh předka. Praha: Academia, 2008.
- GERHART, John, Christopher Lowe, Marc Kirschner. Hemichordates and the origin of chordates. Current Opinion in Genetics & Development. 2005-08, roč. 15, čís. 4, s. 461–467. Dostupné online. ISSN 0959-437X. DOI 10.1016/j.gde.2005.06.004.
- Problematic Cambrian Fossil, Possibly a Primitive Chordate Fossil [online]. The Virtual Fossil Museum, Fossils of the Cambrian Explosion from Utah. Dostupné online.
- Chen, J-Y., Hang, D-Y., and Li, C.W. An early Cambrian craniate-like chordate. Nature. December 1999, roč. 402, s. 518–522. Dostupné online [cit. 2008-09-23]. DOI 10.1038/990080.
- Taxonomicon: Taxon: Phylum Chordata Bateson, 1885 - chordates [online]. Dostupné online.
- ZENG, Liyun, Billie J. Swalla. Molecular phylogeny of the protochordates: chordate evolution. Canadian Journal of Zoology. 2005-01-01, roč. 83, s. 24–33. Dostupné v archivu pořízeném dne 2012-01-11.
- WINCHELL, Christopher J, Jack Sullivan, Christopher B Cameron, Billie J Swalla, Jon Mallatt. Evaluating hypotheses of deuterostome phylogeny and chordate evolution with new LSU and SSU ribosomal DNA data. Molecular Biology and Evolution. 2002-05, roč. 19, čís. 5, s. 762–776. Dostupné online [cit. 2010-03-30]. ISSN 0737-4038.
- PHILIPPE, Hervé, Nicolas Lartillot, Henner Brinkmann. Multigene analyses of bilaterian animals corroborate the monophyly of Ecdysozoa, Lophotrochozoa, and Protostomia. Molecular Biology and Evolution. 2005-05, roč. 22, čís. 5, s. 1246–1253. Dostupné online [cit. 2010-03-30]. ISSN 0737-4038. DOI 10.1093/molbev/msi111.
- DELSUC, Frédéric, Henner Brinkmann, Daniel Chourrout, Hervé Philippe. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature. 2006-02-23, roč. 439, čís. 7079, s. 965–968. Dostupné online [cit. 2010-03-30]. ISSN 1476-4687. DOI 10.1038/nature04336.
- Is everything OK with Olfactores? A call to properly assess morphological implications [online]. 2008. Dostupné online.
Literatura
- ROČEK, Zbyněk. Historie obratlovců : evoluce, fylogeneze, systém. Praha: Academia, 2002. 512 s. ISBN 80-200-0858-6.
- ZRZAVÝ, Jan. Fylogeneze živočišné říše. Praha: Scientia, 2006. 255 s. ISBN 80-86960-08-0.
- HAJER, Jaromír. Fylogeneze a systém strunatců. Ústí nad Labem: Univerzita JAP, přírodovědecká fakulta, katedra biologie, 2006. Dostupné v archivu pořízeném dne 2012-01-10. Archivováno 10. 1. 2012 na Wayback Machine
Externí odkazy
 Obrázky, zvuky či videa k tématu strunatci na Wikimedia Commons
Obrázky, zvuky či videa k tématu strunatci na Wikimedia Commons  Taxon Chordata ve Wikidruzích
Taxon Chordata ve Wikidruzích- (anglicky) Kmen strunatci na projektu „Tree of Life“ Archivováno 24. 2. 2007 na Wayback Machine
- (česky) Kurzy morfologie živočichů na rocek.gli.cas.cz