Lišejník
Lišejník (latinsky Lichen), někdy lichenizovaná houba,[1] je symbiotické společenství houby (mykobionta) a řasy či sinice (tedy fotobionta neboli fykobionta). Vědecký obor studující lišejníky se nazývá lichenologie.
.jpg.webp)



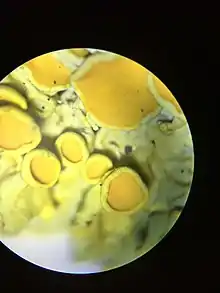
Fotobiontem je řasa nebo sinice, mykobiont je vřeckovýtrusná nebo vzácně i stopkovýtrusná houba. Vzájemné vztahy mezi fotobiontem a mykobiontem nejsou ještě přesně zjištěny, mohou přecházet od oboustranně výhodného svazku až k negativnímu (parazitismu).[2] Specifický vztah v lišejníku se označuje jako lichenismus.
Je známo přibližně 13 500 – 17 000 druhů lišejníků[3] a každoročně jsou popisovány další. Lišejníky mají své vědecké i české taxonomické názvy, přičemž jméno lišejníku je zároveň jménem houby, příkladem může být terčovka bublinatá (Hypogymnia physodes). České názvy však byly většinou vytvořeny v 19. století a jsou zastaralé, neodpovídají dnešnímu zařazení lišejníků v systému hub. Řasy či sinice mají svá zvláštní jména.[1] Z celkového počtu druhů hub tvoří lichenizované houby asi 21 %.[1]
Etymologie slova
Lišejník je latinsky, francouzsky i anglicky lichen, německy však atypicky Flechte. Základem pro románské i další názvy je pravděpodobně řecké leichen, které pochází ze slovesa leichein, lízat. Původně se anglické lichen používalo pro játrovky, první užití v dnešním slova smyslu bylo zaznamenáno roku 1715.[4]
Symbiotický vztah
Symbióza lišejníků je poměrně svérázný způsob soužití dvou i více odlišných biologických druhů. Tato symbióza se nazývá lichenismus. Symbiotickou povahu lišejníků objevil roku 1867 Švýcar Simon Schwendener[5] a trvalo téměř sto let, než ve vědecké obci definitivně vymizely námitky proti tomuto faktu. Poslední práce popírající lišejníky jako symbiotické asociace byla uveřejněna roku 1953.[1]
Typický lišejník se skládá z mykobionta, jímž může být vřeckovýtrusná (Ascomycota) nebo vzácně i stopkovýtrusná (Basidiomycota) houba, a z fotobionta. Fotobiont, tedy fotosyntetizující část lišejníku, může být jednobuněčná nebo vláknitá řasa, ale asi v 8 procentech i prokaryotická sinice (Cyanobacteria, tvoří tzv. cyanolišejníky[6]). Z řas jsou fotobionty nejčastěji zelené řasy (Chlorophyta). Řasa rodu Trebouxia je pravděpodobně nejčastějším fotobiontem lišejníků, je součástí asi 2/3 všech lišejníků[1] (podle jiných zdrojů ale jen 20 %,[3] uváděna jsou i jiná čísla). Dále jsou známy řasy z rodů Coccomyxa či Trentepohlia.
Někdy však lišejníky obsahují i zcela odlišné skupiny řas. Je takto známo několik různobrvek (Xanthophyceae)[7] a v jednom případě dokonce i hnědá řasa.[8] Během dlouhé doby fylogeneze společného svazku se vyvinula vzájemná závislost houby a řasy v lišejníku, takže dnes jen ojediněle mohou žít některé řasy a cyanobakterie tvořící lišejníky samostatně. Tyto výjimky patří např. do rodů Trentepohlia a Nostoc.
Rozlišujeme dva základní typy vzájemných vztahů mezi mykobiontem a fotobiontem, mutualistický a poloparazitický.[1]
Mutualistický vztah
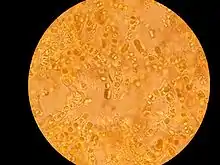
Oba partneři v mutualistickém vztahu si navzájem poskytují výhodné služby. Fotobiont poskytuje organické látky, které vyrobil fotosyntézou, houba anorganické látky, vodu, vhodné prostředí pro růst a výměnu plynů.[9] Situace je však velmi komplikovaná – pravděpodobně existují různé přechody mezi mutualismem a dalšími typy vztahů, jako je komenzálismus a parazitismus.
Přesto se zdá, že mykobiont má ze symbiotického svazku větší prospěch.[1] V tomto smyslu se někdy uvažuje, že je fotobiont jen „zajatcem“ houby, která jej vysává, ale přitom se stará, aby fotobionta neusmrtila. Takový vztah se nazývá kontrolovaný parazitismus.[9] Někdy také bývá toto soužití popisováno spíš jako vzájemné získávání než jako spolupráce.[10] Obecně se dá říci, že nejmírumilovnější je houba v těch lišejnících, které žijí na extrémních stanovištích (např. holé skály). Na těchto místech totiž na sobě fotobiont a mykobiont závisí daleko více. Takový vztah se někdy popisuje jako hladová symbióza.[11]
Poloparazitický vztah
V poloparazitickém vztahu působí mykobiont fotobiontovi újmu, může způsobit částečné nebo i úplné odumření. Neexistuje však přesná hranice mezi poloparazitickým vztahem a stavem, kdy houba zcela parazituje na řase (jako např. algikolní Epigloea) nebo se na jejích odumřelých tkáních vyživuje saprofyticky. U některých korovitých lišejníků dokonce houba svými chemickými látkami ředí stěnu řasy a vstupuje dovnitř svými haustorii.[11]
Zvláštní případy
Existuje mnoho druhů lišejníků, které se odchylují od klasického vzoru, popsaného výše.
Někdy je poměr biomasy mykobionta a fotobionta obrácený – řasa výrazně převládá nad houbou. Například chaluha druhu Ascophyllum nodosum je posetá drobnými plodnicemi symbiotické vřeckovýtrusné houby Mycosphaerella ascophylli. Houba žije v mezibuněčných prostorech chaluhy. Část vědecké obce se domnívá, že tato houba pomáhá překonávat nepříznivé podmínky za odlivu.[9] Podobně je na tom hnědá řasa Pelvetia canaliculata. Tento specifický vztah mezi houbou a řasou se nazývá fotomykobióza. Podobný vztah tvoří řasa Desmococcus vulgaris, která roste na kůře stromů a bývá často asociována s podhoubím hub.[1] S arbuskulární mykorhizou hraničí zvláštní symbiotický svazek houby Geosiphon pyriformis z oddělení Glomeromycota se sinicemi rodu Nostoc. Někdy je považován za jediný známý lišejník z tohoto oddělení,[12] jindy se Geosiphon popisuje spíše jako houba, která ztratila schopnost klasické mykorhizy a vytvořila náhradní symbiotický vztah se sinicemi.[9] Existují také houby, které mohou tvořit lišejník s různými druhy fotobiontů. Dva lišejníky se stejným mykobiontem a různým fotobiontem se nazývají fototypy (fotomorfy). Vznikají tak lišejníky, které někdy dokonce mají různý vzhled. Některé druhy laločníku (Sticta) tvoří s určitým fotobiontem keříčkovitou stélku, s jiným zase stélku lupenitou.[1]
Někdy bývá partnerů více než dva. Asi 500 druhů lišejníků přibírá do asociace druhého fotobionta, jeden však z nich většinou dominuje. Druhý, méně početný fotobiont pak většinou sídlí v ostrůvcích nebo tzv. cefalodiích na povrchu. Někdy mohou být fotobionti i tři.[1] Jsou i lišejníky s více než jedním mykobiontem. Druhá houba se pak označuje jako parasymbiont či jako lichenikolní houba. Někdy tyto houby působí destruktivně na celou stélku, někdy se jen přiživují.[1] Objevují se stále nové lichenikolní houby, známe přibližně 1000 druhů lichenikolních hub.[1] (seznam českých druhů viz[13]) Zásluhou vícečlenných vztahů může vzniknout lišejník, který obsahuje například dva mykobionty a dva fotobionty.[9]
Studie z r. 2016 pomocí genetického rozboru ukázala, že běžné lišejníky s vřeckovýtrusou lichenizovanou houbou obsahují jako druhého mykobionta jednobuněčné kvasinky z linie stopkovýtrusých. Ty jsou usídleny v kůře stélky a mohou mít podstatný vliv na její morfologii i metabolismus. Rozdílné druhy lišejníků se stejnou lichenizovanou houbou tak mohou být způsobeny nejen odlišným fotobiontem, ale i pouhým rozdílem v symbiotické kvasince.[pozn. 1] Lišejníky se třetím, kvasinkovým symbiontem byly nalezeny na všech šesti kontinentech. Prokáže-li se totéž u většiny druhů, může objev zvrátit současné paradigma jediného hlavního mykobionta a za standardní lišejník bude nutno považovat symbiózu tří organismů – lichenizované mnohobuněčné houby, jednobuněčné kvasinky a fotosymbionta.[14][15][16]
Stélka

Tělo lišejníku je stélka, složená (většinou z jednoho jedince) houby (tzv. mykobiont) a z milionů jedinců fotosyntetických mikroorganismů (tedy tzv. fotobionta). Stélka lišejníku je ve většině případů uspořádaná a řasa se v ní vyskytuje v gonidiové vrstvě (běžně „řasová vrstva“) pod korovou houbovou vrstvou na povrchu lišejníku.
Vnější vzhled stélky
Existuje několik druhů stélky lišejníků:
- korovitá – pevně srůstají s podkladem, často obrůstají skaliska i borku stromů, mnohé z nich patří mezi mikrolišejníky; např. mapovník zeměpisný (Rhizocarpon geographicum), který přirůstá celou plochou ke skalnímu podkladu
- lupenitá – stélky lnou k podkladu jen částečně, jsou od podkladu oddělitelné; např. terčovka bublinatá (Hypogymnia physodes);
- keříčkovitá – z jednoho místa vyrůstají dlouhé (až metrové) stélky; např. dutohlávka sobí (Cladonia rangiferina), která bývá větvená, odstává od podkladu a tvoří „keřík“;
- vláknitá – stélka je tvořena dlouhými vlákny; např. provazovka (Usnea).
Stélka některých druhů lišejníků je dvoutvárná (dimorfní), kombinuje tedy v sobě více druhů stélek. Příkladem je rod dutohlávka (Cladonia), který se sice považuje za keříčkovitý lišejník, ale ve skutečnosti výrazná keříčkovitá stélka vyrůstá z druhé, lupenité části na povrchu půdy.[17]
Anatomie lišejníku


Anatomie lišejníků je značně odvislá od mykobionta, který utváří většinu stélky, ale i od fotobionta. Pokud mykobiont může spolupracovat s více druhy fotobiontů, vznikají pak často i velmi rozdílné stélky, rozlišitelné pouhým okem.[9]
Některé lišejníky mají zcela primitivní, téměř nepravidelnou stélku. Označuje se jako homeomerická stélka, má ji např. rod huspeník (Collema).[9] Taková stélka mívá díky slizu sinic slizovitou konzistenci, někdy je ale naopak práškovitá a rozpadavá.[12]
Běžné lišejníky jsou však rozlišené na několik vrstev a vytváří tzv. heteromerickou stélku. Na jejím povrchu se nachází ochranná vrstva kůry, tvořená houbovými vlákny mykobionta se ztluštělými buněčnými stěnami,[1] tedy houbovým pletivem zvaným pseudoparenchym.[18] Z této vrstvy někdy také vyrůstají plodnice houby, sloužící k pohlavnímu rozmnožování mykobionta (viz obrázek). Pod ní leží úzká vrstva zelených fotosyntetizujících buněk obalených houbovými vlákny, tedy jediná vrstva fotobionta. Toto pletivo je rozvolněné, plektenchymální.[18] Pod ní se nachází dřeň, tvořená mykobiontem a sloužící jako zásobárna vody. Pod dřeňovou vrstvou se někdy nachází další vrstva spodní kůry, nebo navíc rhiziny (příchytné kořínky, někdy též rhizoidy[18]) Spodní korová vrstva mívá v sobě trhliny, určené k výměně plynů.
Kulovité útvary na povrchu některých lišejníků (např. u některých hávnatek rodu Peltigera) se nazývají cefalodium. Tyto útvary se objevují zejména u lišejníků s vícečlennými vztahy. Hostí totiž druhého fotobionta, sinici (velmi často rodu Nostoc), který má za úkol fixovat ze vzduchu dusík.
Houbová vlákna tvoří haustoria, jimiž houby vysávají z fotobiontů živiny. Haustoria však do buněk řas pronikají jen u lišejníků s korovitou stélkou. U keříčkovitých a lupenitých stélek se vlákna jen omotávají kolem buněk fotobionta.[8]
Růst a věk
Lišejníky jsou považovány za nejpomaleji rostoucí organismy,[18] přestože v českých podmínkách rostou lišejníky téměř celoročně (i pod bodem mrazu[19]). Nejpomaleji rostou druhy s korovitou stélkou, například lišejník zeměpisný (Rhizocarpon geographicum) roste rychlostí 0,06 až 1 mm za rok. Dutohlávka roste rychleji, asi 2,5 mm za rok, některé lupenité a keříčkovité lišejníky rostou až 15 mm/rok[18] To je důležité například pro správné vyhodnocení bioindikace pomocí lišejníků (viz níže).
Na druhou stranu se dožívají lišejníky velmi vysokého věku. Epifytické druhy žijí několik desítek let, saxikolní druhy (také epilitické, rostoucí na kamenech) se dožívají i několika set let.[19] Stáří stélek lišejníku zeměpisného v nejvyšších českých pohořích se odhaduje na 200 – 400 let, některé arktické lišejníky však dosahují stáří až 4000 let (srovnatelné s tisy a sekvojemi).[18]
Rozmnožování

Lišejníky se rozmnožují především vegetativně. Důvodem je zřejmě to, že houby se ve většině případů neobejdou bez svého fotobionta. Někdy vzniká nový lišejník prostou fragmentací stélky na několik částí.[18] Hlavními mechanismy vegetativního rozmnožování jsou ale tzv. izidie a soredie, které vznikají fragmentací stélky. Izidie jsou dobře viditelné výrůstky stélky, které se odlupují od mateřské stélky a dorůstají v nového jedince. Soredie jsou malé (pod 1 mm) a vznikají jako prášek na tzv. sorálech, místech, kde není stélka lišejníku přikrytá kůrou.
Někdy dochází k pohlavnímu rozmnožování samotné houby pomocí plodnic s výtrusy. Plodnice sice někdy obsahují fotobionta, ale ten se většinou s výtrusy nešíří. Výjimkou jsou rody Endocarpon a Staurothele, které jsou v symbiotickém svazku s tzv. hymeniálními řasami. Buňky těchto řas jsou v hymeniu (výtrusorodém roušku) společně s výtrusy hub a společně se také šíří větrem nebo vodou.[1]
Je známo více druhů plodnic, z nichž jsou tři nejběžnější: apothecium, perithecium a pyknidium. Apotheciové plodnice bývají vyduté a otevřené nahoru, i když to nemusí být pravidlem. Mezi vřecky jsou zvláštní sterilní vlákna, tzv. parafýzy, které často obsahují pigment. Barevnost apothecií může být důležitá pro determinaci.[20]
Perithecium a pycnidium vypadají na první pohled velmi podobně, jedná se v obou případech o dutinky vrostlé do stélky lišejníku. Zatímco však perithecia obsahují askospory (podobně jako apothecium), pyknidium obsahuje nepohlavně vznikající konidie (konidiospory).[20]
Paleontologické nálezy lišejníků
Na místech, kde obyčejně rostou lišejníky, vznikají fosilie jen vzácně. Přestože známe z Jižní Afriky nálezy již ze starších starohor, které připomínají primitivní lišejníky, za nejstarší nález skutečného lišejníku se považuje lišejník z mladšího devonu z lokality ve Skotsku, je tedy asi 400 milionů let starý.[21] Kandidátem na lišejník jsou i podobně staré, záhadné fosílie Spongiophyton a Nematothallus.
Klasifikace lišejníků

Zatímco dříve se lišejníky řadily do zvláštního rostlinného oddělení, po poznání jejich podstaty jakožto hub s endosymbiotickým fotobiontem se dnes zásadně klasifikují podle mykobionta, podle pravidla „jedna houba – jeden lišejník“.[1] Jedná se tedy o zástupce říše houby (Fungi), podříše Dikarya, kteří však jakožto lišejníky netvoří společně žádnou přirozenou skupinu. Známe 13 500 – 17 000 lichenizovaných hub. Mezi ně patří zejména vřeckovýtrusné houby (Ascomycota), ale vzácně (méně než 2 %[22]) i stopkovýtrusné houby (Basidiomycota). U některých lichenizovaných hub se nepodařilo pozorovat pohlavní rozmnožování, které by umožnilo klasifikaci do jedné z hlavních skupin Dikarya; v některých systémech jsou proto prozatím podřazeny umělé skupině Deuteromycota[3] a o jejich zařazení mohou rozhodnout v budoucnosti až molekulární analýzy jejich genomu.
Má se za to, že se lichenismus nezávisle vyvinul mnohokrát, a to nejen jednou u vřeckovýtrusých a podruhé u stopkovýtrusých hub. Molekulární analýzy ukazují, že se tak stalo i uvnitř některých tříd hub vícekrát nezávisle na sobě, zároveň jsou poznatky o druhotné ztrátě této schopnosti u některých podřazených taxonů.[23]
Klasifikace na úrovni druhu a nižších taxonů podle principu „jedna houba – jeden lišejník“ dosud budí kontroverze, a to zejména proto, že se nelichenizované a lichenizované houby stejného druhu, jakož i stejný druh s odlišnými fotosymbionty mohou výrazně odlišovat morfologicky a v návaznosti na to i stylem života, a mohou mít i odlišnou a nezávislou evoluční historii.[23][24]
Askolišejníky
Lišejníků tvořených vřeckovýtrusnou houbou (askolišejníků) je 98 %.[22] Asi polovina všech známých vřeckovýtrusných hub je lichenizována.[3] Nejvíce lišejníků patří do třídy Lecanoromycetes a v jejím rámci především do podtřídy Lecanoromycetidae.[25] Podle dostupných studií, založených na fylogenetické příbuznosti, se rozlišují tři základní řády:[26][27]
- řád misničkotvaré (Lecanorales), např. terčovka bublinatá (Hypogymnia physodes)
- řád hávnatkotvaré (Peltigerales), např. hávnatka psí (Peltigera canina)
- řád krásníkotvaré (Teloschistales), např. terčovník zední (Xanthoria parietina)
Známe však i další skupiny askolišejníků, řazené buď rovněž do třídy Lecanoromycetes (podtřídy Ostropomycetidae, Acarosporomycetidae), nebo do zcela jiných taxonomických skupin (např. Lichinomycetes, Arthoniomycetes, Dothideomycetes).[28]
Bazidiolišejníky
Některé lišejníky, tzv. bazidiolišejníky, patří do řádů lupenotvaré (Agaricales) a chorošotvaré (Aphyllophorales)[3] z oddělení stopkovýtrusných hub. Sem patří zejména rod Lichenomphalia. Tyto houby vytvářejí stélky, z nichž vyrůstají velmi nápadné plodnice v klasickém slova smyslu. Poměrně známá je i tužnatka slizká (Multiclavula mucida).
Výskyt a význam


Lišejníky najdeme téměř všude[29] s výjimkou volného oceánu. Lišejníky byly například objeveny pouhých 300 kilometrů od pólu. Terčovník pohledný (Xanthoria elegans) byl nalezen i v nadmořské výšce 7000 m n. m. Lišejník misnička jedlá (Lecanora esculenta) se vyskytuje na poušti Sahara. Někdy rostou lišejníky i ve vodě, jako například nitroplodka potoční (Dermatocarpon rivulorum).[19] I když jsou schopné porůstat všechny substráty, nejčastěji je nalézáme na kamenech a skalách, na půdě a jako epifyty na stromech a keřích. To jsou místa, kde je malá konkurence vyšších rostlin, proti nimž se lišejníky například v hustém lesním podrostu neprosadí.[9] Mnoho druhů také vyžaduje stabilní substrát (horniny), protože roste pomalu. Na druhou stranu existují i mnohé efemery, které osídlují také rychle tlející dřevo, listy dřevin v tropech či kamenité suti.
Konkurenční výhodou lišejníků je, že jsou poikilohydrické, snášejí velké výkyvy obsahu vody ve stélce. Když obsah vody klesne, vstupují lišejníky do stavu, známého jako kryptobióza. V tom dokáží vzdorovat i překvapivě nehostinným podmínkám. Pokusy ve vesmíru ukázaly, že lišejníky přežijí i pobyt ve studeném a mrtvém prostoru na oběžné dráze Země,[30] kde je především velmi silné UV záření. Důležité je i to, že lišejníky nemají kořeny, a proto nepotřebují stálý přísun vody. Vodu pohlcují celým povrchem těla.
Lišejníky často osídlují nově vzniklá nebo ještě neosídlená stanoviště. Proto patří k pionýrským druhům. Na poušti zpevňují půdu před erozí asi milimetr silnou kryptobiotickou krustou, která se postupně rozkládá a do půdy dodává živiny. Tak se tvoří místo, které později mohou osídlit i cévnaté rostliny.[31] Podobně lišejníky kolonizují též povrchy skal, kde naopak erozi podporují. Na skalních substrátech, kde houbová vlákna lišejníků pronikají do substrátu a navětrávají ho, vznikají primární půdy a cévnaté rostliny mohou substrát osidlovat. To může vadit na povrchu soch, například v americkém Mount Rushmore.[32] Lišejníky rostoucí na dřevinách nejsou nikdy parazity, nekonzumují žádnou část rostliny ani ji netráví. Vyživují se samostatně, jak bylo výše uvedeno. Řasa fotosyntetizuje, houba čerpá vodu ze srážek s anorganickými látkami v ní obsaženými. Zejména na půdním substrátu však produkují např. lišejníky rodu Cladonia chemikálie, které zastavují klíčení semen a růst mladých rostlin.[33]
Lišejníky jsou důležitým zdrojem potravy pro některé živočichy, zejména pro soby polární (karibu),[31] ale také pro mnoho druhů housenek motýlů.[34] Lišejníky však obsahují málo cukrů, a tak pro některé živočichy nestačí.
Mnoho lišejníků má velmi specifické požadavky na chemismus substrátu. Některé epifytické druhy vyhledávají zásaditou borku, jiné neutrální či kyselou. Některé lišejníky preferují prostředí bohaté na dusík. Mezi ně patří rody Physcia, Xanthoria či druh Amandinea punctata, které proto odolávají i znečištění oxidy dusíku. Nazývají se nitrofilní druhy.
Lišejníky a lidé
Využití člověkem
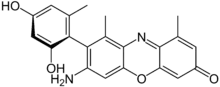
Lišejníky mají pro lidstvo překvapivě velký význam, kterým se dokonce zabývá samostatná věda, zvaná etnolichenologie.[35] Nejčastěji se lišejníky používaly a používají jako barvivo, ale známé jsou i lišejníky užívané k léčbě a jako potravina. Barviva i účinné látky v lécích jsou sekundární metabolity lišejníků.
Barviva se z lišejníků získávají tak, že se stélky uvaří ve vroucí vodě nebo se fermentují v moči. V Severní Americe je významný lišejník Letharia vulpina, z něhož se získávala žlutá barva. Také ve Skotsku se používala barviva z lišejníků – především hnědé a červené barvy. Z lišejníků rodu Roccella se na Kanárských ostrovech, Kapverdách, Madagaskaru a v Indii získávala fialová barviva, kterým se říká orceiny.
Využití lišejníků jako léčiv se dá vysvětlit tím, že lišejníky obsahují velké množství (často unikátních) sekundárních metabolitů, z nichž část má antibiotické účinky. Kyselina usnová z lišejníku provazovka (Usnea) patří k nejznámějším. Užívá se i pukléřka islandská (Cetraria islandica), a to jako prostředek proti kašli.[36] Lišejníky se také sbíraly jako potravina, a to téměř po celém světě. Sice obsahují některé toxiny nebo nestravitelné polysacharidy, ty se však některými metodami dají odstranit. V Evropě byl široce využíván lišejník pukléřka islandská, z níž se připravoval chléb, kaše, polévka i salát. V Severní Americe byl konzumován lišejník Bryoria fremontii. Dutohlávka sobí (Cladonia ranginifera) se tradičně vyjímala z předžaludků ulovených sobů a posléze byla konzumována.
Lišejníky se používají nebo používaly i k výrobě alkoholu, kosmetiky, parfémů, jako dekorace, na textilní vlákna, jako potrava pro dobytek, palivo, v průmyslu jako výchozí surovina, k vyčiňování kůží, k lákání zvěře při lovu, jako repelent proti hmyzu, jed, konzervant, k různým magickým rituálům a dokonce jako narkotikum a halucinogen.[37] Dnes se lišejníky užívají také k určování stáří usazenin a k bioindikaci čistoty ovzduší (viz níže).
Některé nitrofilní druhy na Aljašce původní obyvatelé využívali k lovu svišťů, protože tyto lišejníky rostly na místech, kde zmínění hlodavci zanechávali své (na dusík bohaté) exkrementy.[19]
Mytologie
Podle legendy severoamerického kmene Menomini jsou lišejníky strupy spadlé z hlavy boha jménem Må'nåpus, podle jiné verze strupy z jeho hýždí, které si způsobil, když se smekl po prudké skále.[38] Indiáni kmene Paiute z Nevady nazývali oranžové a žluté skalní lišejníky „semeno ještěrů“. Leguánci druhu Sceloporus occidentalis totiž dělají na skalách drobné pohyby těla dolů a nahoru.[37] Indiáni kmene Okanagan mají několik legend na téma, jak vznikl lišejník Bryoria fremontii. V jedné se kojot pokouší ulovit labuť. Ta však vzlétne a kojota vezme s sebou do vzduchu. Pouští ho až ve výšce a kojot padá do stromů a srst se mu zamotává ve větvích. Potom promění kojot své chlupy v lišejník slovy: „Má srst nesmí být zmařena. Lidé tě budou sbírat a připravovat z tebe jídlo“.[39]
Citlivost ke znečištění ovzduší
Pro lišejníky je typická schopnost tzv. bioindikace čistoty ovzduší, vzhledem k jejich citlivosti ke znečištění. Nejcitlivější jsou obecně druhy s keříčkovitou stélkou (provazovka, vousatec, a podobně), tolerantní jsou např. misničky (Lecanora)[18] a některé efemérní lišejníky s krátkou vegetační dobou. Cyanolišejníky (lišejníky s fotobiontem sinicí) jsou obecně velmi citlivé.[6]
Lišejníkům škodí zejména imise oxidu siřičitého (SO2). Ten jim narušuje vnitřní stabilitu buněk, znemožňuje fotosyntézu a má vliv na rozmnožování.[19] Do 35 μg/m3 se vyskytují provazovky (Usnea), do 70 μg/m3 terčovky (Parmelia), při vyšších koncentracích lišejníků rychle ubývá a při 170 μg/m3 rostou na kůře jen zelené řasy.[11] Z těchto znalostí můžeme zpětně určovat přibližné zatížení ovzduší SO2. S klesajícím znečištěním oxidem siřičitým v dnešní době se lišejníky pomalu vracejí na některá místa, kde byly vyhubeny. Dokazují to studie z Lucemburské zahrady v Paříži,[29] z údolí řeky Ohio v USA[40] či z Londýna.[41] Ještě v roce 1989 se však psalo, že do roku 2000 pravděpodobně u nás vyhynou všechny keříčkovité a lupenité lišejníky.[18]
Mezi další látky, které mají negativní vliv na růst lišejníků, patří sloučeniny těžkých kovů, fluor, brom a radioaktivní spad. Dokládají to studie provedené v okolí hliníkáren (např. Žiar nad Hronom), kde bývají vysoké koncentrace fluoru v ovzduší. V nejbližším okolí hliníkáren se dokonce tvoří tzv. lišejníková poušť.[19] Lišejníky doplácí na to, že jsou schopné dobře nasávat škodliviny. Po černobylské katastrofě vstřebaly lišejníky ve Skandinávii tolik radioaktivních částic, že maso sobů, kteří se jimi živí, bylo považováno za nevhodné ke konzumaci.[31][42]
Středoevropské druhy
Ve střední Evropě se vyskytuje asi 2000 druhů lišejníků. Mezi lišejníky známé v ČR patří například dutohlávka sobí (Cladonia rangiferina), terčovka bublinatá (Hypogymnia physodes), mapovník zeměpisný (též lišejník zeměpisný, Rhizocarpon geographicum), hávnatka psí (Peltigera canina), pukléřka islandská (Cetraria islandica), terčovník zední (Xanthoria parietina), rod provazovka (Usnea) a mnohé další.



.jpg.webp)
Odkazy
Poznámky
- Příkladem jsou druhy Bryoria tortuosa a Bryoria fremontii. Zatímco první z nich je žlutý díky vytvářené kyselině vulpinové, kterou je schopen syntetizovat jako ochrannou látku před spásáním i mikrobiálními infekcemi, druhý je tmavě hnědý a tento produkt netvoří. Přitom oba lišejníky tvoří stejná řasa (Trebouxia simplex) a houba (vousatec, Bryoria, Lecanoromycetes).
Reference
V tomto článku byly použity překlady textů z článků Lichen na anglické Wikipedii a Ethnolichenology na anglické Wikipedii.
- LIŠKA, Jiří. Vázaný a nevázaný život lišejníků. Vesmír. Listopad 2000, roč. 79 (130), čís. 11, s. 623–629. ISSN 1214-4029.
- SLAVÍKOVÁ, Jiřina. Ekologie rostlin. Praha: Státní pedagogické nakladatelství, 1986. S. 366.
- NASH, Thomas H. Lichen Biology. [s.l.]: Cambridge University Press, 1996. Dostupné online. ISBN 0521459745. S. 315. (anglicky)
- Online Etymology Dictionary. termín lichen [online]. Douglas Harper, 2001. Dostupné online. (anglicky)
- HONEGGER, Rosmarie. Simon Schwendener (1829–1919) and the Dual Hypothesis of Lichens. The Bryologist. 2000-06-01, roč. 103, čís. 2, s. 307–313. Dostupné online [cit. 2016-03-11]. ISSN 0007-2745. DOI 10.1639/0007-2745(2000)103[0307:SSATDH2.0.CO;2].
- PALICE, Zdeněk; HALDA, Josef P. Neviditelný svět mikrolišejníků. Živa. 2005, roč. LIII, čís. 2, s. 57–59. ISSN 0044-4812.
- THE DELWICHE LAB, autor neznámý. Photosynthetic Life, Heterokontophyta III — Xanthophyceae & Eustigmatophyceae [online]. 1998. Dostupné online. (anglicky)
- MONTEREY BAY AQUARIUM RESEARCH INSTITUTE, autor neznámý. Caloplaca coralloides lichen symbiosis [online]. 2005. Dostupné v archivu pořízeném dne 2008-04-15. (anglicky)
- ČEPIČKA, Ivan; KOLÁŘ, Filip; SYNEK, Petr. Mutualismus, vzájemně prospěšná symbióza; Přípravný text – biologická olympiáda 2007–2008. Praha: NIDM ČR, 2007. Dostupné v archivu pořízeném dne 20-09-2008. S. 87. Archivováno 20. 9. 2008 na Wayback Machine
- CAMPBELL, Neil A.; REECE, Jane B. Biologie. Praha: Computer press, 2006. S. 1332.
- RABŠTEINEK, Otomar; PORUBA, Miroslav; SKUHROVEC, Josef. Lišejníky, mechorosty a kapraďorosty ve fotografii. Praha: Státní zemědělské nakladatelství, 1987. S. 224.
- SEDLÁŘOVÁ, M; VAŠUTOVÁ, M. Lichenes [online]. 2004-2007. Dostupné online.
- KOCOURKOVÁ, Jana. Lichenicolous fungi of the Czech Republic (The first commented checklist). Acta Mus. Nat. Pragae. 2000, roč. 55, čís. 3–4, s. 59–169. ISSN 0044-4812.
- SPRIBILLE, Toby; TUOVINEN, Veera; RESL, Philipp; VANDERPOOL, Dan; WOLINSKI, Heimo; AIME, M. Catherine; SCHNEIDER, Kevin, STABENTHEINER, Edith; TOOME-HELLER, Merje; THOR, Göran; MAYRHOFER, Helmut; JOHANNESSON, Hanna; McCUTCHEON, John P. Basidiomycete yeasts in the cortex of ascomycete macrolichens. S. 488–492. Science [online]. 29. červenec 2016. Svazek 353, čís. 6298, s. 488–492. Dostupné online. Dostupné také na: . ISSN 1095-9203. DOI 10.1126/science.aaf8287. (anglicky)
- VAN HOOSE, Natalie, Purdue University. Yeast emerges as hidden third partner in lichen symbiosis. Agriculture News [online]. 21. červenec 2016. Dostupné online. (anglicky)
- HOUSER, Pavel. Třetí do party – lišejník tvoří i kvasinky. SCIENCEmag.cz [online]. 2. listopad 2016. Dostupné online.
- ROSYPAL, Stanislav. Nový přehled biologie. [s.l.]: Scientia, 2003. S. 797.
- KLÁN, Jaroslav. Co víme o houbách. Praha: Státní pedagogické nakladatelství, 1989. S. 310.
- SKALKA, Michal. Lišejníky jako bioindikátory. Živa. 2004, roč. 79 (130), čís. 3, s. 107–108. ISSN 0044-4812.
- RAMEL, Gordon. Lichen Reproductive Structures [online]. Rev. 2008. Dostupné online. (anglicky)
- Anonym. Fossil Record of Lichens [online]. University of California; Museum of Paleontology. Dostupné v archivu pořízeném dne 2010-01-25. (anglicky)
- LUTZONI, François, et al. Evolution of lichens; Consequences of mutualism (lichenization) on the phylogeny and diversification of Ascomycetes [online]. Department of Biology, Duke University, Durham: Francois Lutzoni's Lab, rev. 2001. Dostupné v archivu pořízeném z originálu dne 2008-10-11. (anglicky)
- Australian Lichens. Classification (and lichenization & de-lichenization). Australian National Herbarium & Australian National Botanic Gardens, 2012. Dostupné online (anglicky)
- Australian Lichens. Form and structure: Sticta and 'Dendriscocaulon'. Australian National Herbarium & Australian National Botanic Gardens, 2012. Dostupné online (anglicky)
- ZICHA, Ondřej. Biolib – Lichens (lišejníky), sekce taxonomie [online]. Dostupné online. (anglicky)
- HIBBETT, David S. A higher-level phylogenetic classification of the Fungi [online]. Mycological research, 2007. Dostupné online. (anglicky)
- [Field Museum]. Myconet [online]. 2007. Dostupné v archivu pořízeném dne 2009-03-18. (anglicky)
- LUTZONI, François, et al. Assembling the fungal tree of life: progress, classification, and evolution of subcellular traits. American Journal of Botany. 2004, roč. 91, s. 1446–1480. Dostupné online. (anglicky)
- ČERNOHORSKÝ, Zdeněk. Lišejníky rostou všude. Vesmír. Listopad 2000, roč. 79 (130), čís. 11, s. 629–630. ISSN 1214-4029.
- MIHULKA, Stanislav. Lišejníky na oběžné dráze [online]. OSEL.CZ, 2005-12-12 [cit. 2008-03-04]. Dostupné online.
- DEACON, Jim. Lichen biology (continued) [online]. Dostupné v archivu pořízeném z originálu dne 2012-02-29. (anglicky)
- THE ASSOCIATED PRESS. Mount Rushmore faces finally washed [online]. HighBeam encyclopedia, 30. července 2005. Dostupné v archivu pořízeném z originálu dne 2008-04-15. (anglicky)
- HOBBS, R. J. The persistence of Cladonia patches in closed heathland stands. Lichenologist. 1985, čís. 17, s. 103–109. Dostupné online. (anglicky)
- SIMONSON, Sara. Lichens and Lichen-Feeding Moths (Arctiidae: Lithosiinae) as Bioindicators of Air Pollution in the Rocky Mountain Front Range [online]. 1996. Dostupné v archivu pořízeném z originálu dne 2008-12-01. (anglicky)
- Ethnolichenology [online]. Dostupné online. (anglicky)
- Iceland moss remedies [online]. Gaia Garden Herbals. Dostupné v archivu pořízeném z originálu dne 2012-02-04. (anglicky)
- SHARNOFF, S. D. Lichens and people, Lichen use [online]. 1997. Dostupné online. (anglicky)
- SMITH, H. H. Ethnobotany of the Menomini Indian. Bulletin of the Public Museum of the City of Milwaukee. 1923, čís. 4(1), s. 1–174. (anglicky)
- TURNER, N. J.; BOUCHARD, R.; KENNEDY, D. I. D. Ethnobotany of the Okanagan-Colville Indians of British Columbia and Washington. Occasional Papers of the British Provincial Museum. 1980, čís. 21, s. 1–179. (anglicky)
- SHOWMAN, Ray E. Continuing Lichen Recolonization in the Upper Ohio River Valley. The Bryologist. 1997, roč. 100, čís. 4, s. 478–481. Dostupné online. (anglicky)
- HAWKSWORTH, David L.; MCMANUS, P. M. Lichen recolonization in London under conditions of rapidly falling sulphur dioxide levels, and the concept of zone skipping. Botanical journal of the Linnean Society. 1989, roč. 100, s. 99–109. Dostupné online. (anglicky)
- BRUCE, Ake; STORACH, Stuart A. Dietary implications of radioactive fallout in Sweden following the accident at Chernobyl. The American Journal of Clinical Nutrition. 1987, roč. 45, s. 1089–1093. Dostupné online. (anglicky)
Literatura
- RABŠTEINEK, Otomar; PORUBA, Miroslav; SKUHROVEC, Josef. Lišejníky, mechorosty a kapraďorosty ve fotografii. Praha: Státní zemědělské nakladatelství, 1987. S. 224.
- KALINA, Tomáš; VÁŇA, Jiří. Sinice, řasy, houby, mechorosty a podobné organismy v současné biologii. Praha: Karolinum, 2005. 606 s.
- KLÁN, Jaroslav. Co víme o houbách. Praha: Státní pedagogické nakladatelství, 1989. S. 310.
- ČEPIČKA, Ivan; KOLÁŘ, Filip; SYNEK, Petr. Mutualismus, vzájemně prospěšná symbióza; Přípravný text – biologická olympiáda 2007–2008. Praha: NIDM ČR, 2007. S. 87.
- LIŠKA, Jiří. Vázaný a nevázaný život lišejníků. Vesmír. Listopad 2000, roč. 79 (130), čís. 11, s. 623–629. ISSN 1214-4029.
- VĚZDA, Antomín; LIŠKA, Jiří. Katalog lišejníků České republiky. Průhonice: Botanický ústav AV ČR, 1999. S. 283.
- NASH, Thomas H. Lichen Biology. [s.l.]: Cambridge University Press, 1996. Dostupné online. ISBN 0521459745. S. 315. (anglicky)
Externí odkazy
 Obrázky, zvuky či videa k tématu lišejník na Wikimedia Commons
Obrázky, zvuky či videa k tématu lišejník na Wikimedia Commons  Slovníkové heslo lišejník ve Wikislovníku
Slovníkové heslo lišejník ve Wikislovníku- Atlas českých lišejníků
- (anglicky) Deacon, J.: Lichens – základní informace o lišejnících s kvalitními fotografiemi
- (anglicky) Deacon, J.: Lichens – fyziologie a anatomie lišejníků
- (anglicky) Deacon, J.: Lichens – ekologie, kryptobiotická půdní krusta
- (anglicky) Checklists of Lichens and Lichenicolous Fungi, seznamy lišejníků vyskytujících se v různých koutech světa
- (anglicky) Lichen Use – praktické možnosti využití lišejníků
- (anglicky) Slovníček termínů o lišejnících, typy stélky, stavba těla