Mitochondria
Mitochondria alebo chondriozóm je organela, ktorá je obalená dvojitou membránou, ktorá má vlastnú genetickú sústavu (tzv. semiautonómna organela), a ktorá sa vo veľkom množstve (stovky až tisíce) vyskytuje v cytoplazme eukaryotických buniek. Slúži prevažne na získavanie energie tzv. bunkovým dýchaním (aeróbnou oxidáciou). Mitochondrie sa však podieľajú aj na ďalších procesoch, ako je bunková diferenciácia, bunková smrť a kontrola bunkového cyklu a rastu. Na druhej strane poruchy ich funkcie môžu mať za následok rôzne mitochondriálne choroby.

1. Vnútorná membrána
2. Vonkajšia membrána
3. Kristy
4. Matrix (základná hmota)
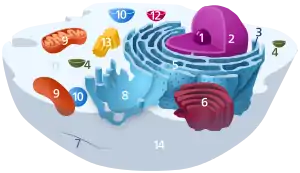
1. Jadierko
2. Jadro
3. Ribozóm (malé bodky)
4. Vezikula
5. Drsné endoplazmatické retikulum
6. Golgiho aparát
7. Cytoskelet
8. Hladké endoplazmatické retikulum
9. Mitochondria
10. Vakuola
11. Cytosól (spolu s ostatnými organelami tvorí cytoplazmu)
12. Lyzozóm
13. Centrozóm spolu s centriolami
14. Cytoplazmatická membrána
V prokaryotických bunkách sa mitochondrie nenachádzajú a enzýmy potrebné na získavanie energie sa v nich nachádzajú v plazmatickej membráne alebo jej záhyboch. Vo veľa baktériách ich funkciu vykonávajú mezozómy, ktorých funkcia nie je celkom objasnená.
Zo živočíšnych somatických (telových) eukaryotických buniek chýbajú iba u extrémne špecializovaných parazitov, napr. Henneguya[1], Haemosporidia a Microspora. U Pelomyxozoa ich funkciu vykonávajú symbiotické baktérie.
Zloženie
Na povrchu sú mitochondrie obalené dvojitou membránou. Vonkajší list tejto membrány je hladký, vnútorný list vbieha vo forme úzkych záhybov do mitochondrie a vytvára priehradky – kristy. Niekedy tieto priehradky nadobúdajú tvar rúrok – tubulárny typ mitochondrií. Kristy zväčšujú vnútorný povrch mitochondrií. Ich počet je priamo úmerný metabolickej aktivite mitochondrie i bunky ako takej. Bunky s vysokou metabolickou aktivitou majú mitochondrie s vyšším počtom kríst, ako bunky s nižšou metabolickou aktivitou. Rovnaký vzťah platí aj pre počet mitochondrií v bunke. Priestor medzi kristami vypĺňa základná hmota – matrix. V nej prebieha Krebsov cyklus, preto sú tu lokalizované všetky potrebné enzýmy. Na kristách dochádza k uvoľňovaniu elektrónov a vzniku H+. Takto získaná energia sa postupne uskladňuje do ATP.
Funkcie
Bunkové dýchanie a syntéza ATP
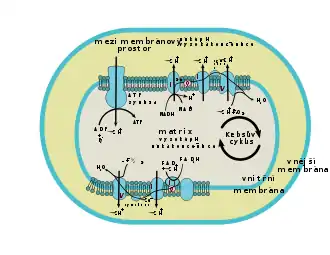
V mitochondriách prebieha prevažná časť bunkového dýchania (výnimku tvorí hlavne glykolýza, ktorá prebieha v cytosole). Tento sled reakcií predstavuje rozklad rôznych organických látok, pričom sa získava energia potrebná pre syntézu adenozíntrifosfátu (ATP). Východiskovou látkou sú hlavne pyruvát a mastné kyseliny. Obe látky v mitochondriálnej matrix prechádzajú reakciami, pri ktorých vzniká acetylkoenzým A (pyruvát prechádza dekarboxyláciou, mastné kyseliny beta-oxidáciou). Acetylkoenzým A následne vstupuje do Krebsovho cyklu, čo je séria reakcií, ktoré umožňujú redukciu koenzýmov NAD+ na NADH a FAD na FADH2. Elektróny z týchto koenzýmov postupujú do dýchacieho reťazca, ktorý je umiestnený na vnútornej membráne mitochondrie. Dýchací reťazec prenáša do mezimembránového priestoru vodíkové katióny (H+, čím vzniká v priestore medzi membránami kyslé pH. Toto pH má sklony sa vyrovnávať, a tak prechádza otvorom v enzýme ATP syntáza späť dovnútra bunky. Priechodom H+ však tento enzým vytvára ATP, ktorý je výsledným produktom všetkých reakcií.
Ďalšie funkcie
Napriek tomu, že mitochondria je známa prevažne ako energetická jednotka bunky, zistilo sa, že má i ďalšie dôležité funkcie. S bunkovým dýchaním súvisí skutočnosť, mitochondrie sú schopné v hnedom tukovom tkanive produkovať teplo. Mechanizmus je elegantný: namiesto toho, aby vodíkové katióny prechádzali ATP syntázou, prechádzajú protónovým kanálom termogenínom (UCP1), ktorý je práve zodpovedný za vznik tepelnej energie.[2][3]
Matrix mitochondrie môže fungovať aj ako zásobáreň vápnika (podobne ako endoplazmatické retikulum), čím udržuje vnútornú homeostázu bunky. Dovnútra sa dostávajú ióny vápnika špeciálnym prenášačom, ktorý je poháňaný membránovým potenciálom,[4][5] pri masívnom uvoľňovaní vápníku von sa môže týmto spôsobom navodiť v bunke určitá odpoveď (produkcia hormónov, neurotransmiterov, a pod.). Navyše sa v mitochondrii nachádza veľa proteínov spúšťajúcich apoptózu, programovanú smrť bunky[6]. Prebiehajú v nich aj niektoré časti syntézy hemu[7] a steroidov.[8]
Ako vedľajší produkt bunkového dýchania mitochondrie produkujú kyslíkové radikály, ktoré môžu spôsobiť početné mutácie v mitochondriálnej DNA.[9] Čím viac sa táto DNA poškodzuje, tým defektnejšie molekuly vznikajú, a tým viac sa môžu degeneratívne zmeny prehlbovať. Množstvo zmien je v mitochondriách pozorovaných v priebehu ľudského starnutia.[10] Niektoré zmeny sa dávajú do súvislosti s Parkinsonovou chorobou.[11]
Vznik
Mitochondrie vznikajú len delením už existujúcich mitochondrií. Majú vlastnú DNA a mitochondriálne ribozómy, čo im umožňuje syntézu potrebných enzýmov. Preto sa o nich hovorí, ako o semiautonómnych organelách.
Evolučný pôvod
.jpg.webp)
V súčasnosti sa všeobecne akceptuje, že mitochondrie predstavujú potomkov endosymbiotickej baktérie, ktorá sa v procese vzniku eukaryotickej bunky určitým spôsobom transformovala na semiautonómnu organelu.[12] Predpokládá sa, že touto baktériou, ktorá vstúpila do primitívnej eukaryotickej bunky, bol zástupca alfaproteobakterií z príbuzenského okruhu rodu Rickettsia.[13] Tieto názory však odborná verejnosť pomerne dlho odmietala ako „nehodiace sa do slušnej prírodovedeckej spoločnosti“. Prvé predstavy o takomto pôvode sa však paradoxne začali objavovať už na konci 19. storočia, kedy ešte nebol ani ustanovený termín mitochondria (napr. Richard Altmann, 1890). Známymi zástupcami tejto tzv. endosymbiotickej teórie bol na začiatku 20. storočia Konstantin Merežkovskij, a v druhej polovici 20. storočia Lynn Margulisová.
Presný scenár „endosymbiotickej udalosti“, teda procesu, pri ktorom sa baktéria zmenila na mitochondriu, je stále relatívne zahalený tajomstvom. Podľa tzv. molekulárnych hodín (metóda založená na pozorovaní mutácií v DNA) sa zdá, že k tomu došlo približne pred 2 miliardami rokov, čiže zrejme nie veľmi dávno po vzniku samotnej eukaryotickej bunky.[14] Dnes sa už považuje za dokázané, že všetky známe eukaryotické organizmy buď mitochondriu majú, alebo aspoň mali za predka eukaryonta, ktorý mitochondriu mal. Preto sa dnes ustupuje od termínu Archezoa, ktorý označoval skupiny eukaryotov, ktorí vznikli ešte pred „endosymbiotickou udalosťou“: žiadne také neboli objavené.[13] Takmer isté je aj to, že mitochondria vznikla v histórii len jedenkrát, a všetky súčasné mitochondrie sú potomkami tejto udalosti (tzv. monofyletický pôvod).
Konkrétny scenár vzniku poskytujú napríklad vodíkové hypotézy, ktoré sú v podstate všetky založené na predpoklade, že hostiteľskou bunkou bola anaeróbna, vodík metabolizujúca autotrofná archebaktéria, ktorá istým spôsobom pohltila symbiotickú baktériu schopnú v rámci svojej respirácie produkovať vodík. Tieto baktérie následne prešli evolúciou a zmenili sa na mitochondrie.[15][16] Bez ohľadu na pôvod po endosymbiotickej udalosti muselo celkom určite dôjsť k čiastočnej redukcii genómu symbiotickej baktérie a pravdepodobne aj k tzv. horizontálnemu transferu, čiže prechodu časti génov z mitochondrie do jadra.
Redukcia mitochondrií
Hoci všetky v súčasnosti známe eukaryotické organizmy pochádzajú zo spoločného predka ktorý mitochondriu mal, u mnohých eukaryotov boli namiesto mitochondrií popísané len akési redukované organely neschopné oxidatívnej fosforylácie, ako sú hydrogenozómy a mitozómy. Tieto organely sú často schopné syntézy ATP bez prítomnosti kyslíka, a hodia sa teda do anaeróbneho prostredia. Hydrogenozómy a mitozómy boli popísané hlavne (nie však výhradne) v taxónoch Excavata a Amoebozoa. Známe sú napríklad Entamoeba histolytica, Giardia intestinalis, rôzne mikrosporídie (Microsporidia) a chytrídiomycety (Chytridiomycota), prípadne rôzne anaeróbne nálevníky (Ciliophora). U niektorých eukaryotov však doteraz neboli popísané ani mitochondrie, ani žiadne iné organely tohto typu.[17]
Referencie
- Radovan Malina: Všeobecná zoológia – zdroj, z ktorého (pôvodne) čerpal tento článok.
- Tento článok je čiastočný alebo úplný preklad článku Mitochondrie na českej Wikipédii (číslo revízie nebolo určené).
- Dayana Yahalomi, Stephen D. Atkinson, Moran Neuhof, E. Sally Chang, Hervé Philippe, Paulyn Cartwright, Jerri L. Bartholomew, Dorothée Huchon. A cnidarian parasite of salmon (Myxozoa: Henneguya) lacks a mitochondrial genome [online]. PNAS, 2020, [cit. 2020-02-28]. Dostupné online. (po anglicky)
- Mozo J, Emre Y, Bouillaud F, Ricquier D, Criscuolo F. Thermoregulation: What Role for UCPs in Mammals and Birds?. Bioscience Reports., november 2005, roč. 25, s. 227–249. DOI: 10.1007/s10540-005-2887-4.
- Nicholls DG, Lindberg O. Brown-adipose-tissue mitochondria. The influence of albumin and nucleotides on passive ion permeabilities. Eur. J. Biochem., 1973, roč. 37, s. R551. DOI: 10.1111/j.1432-1033.1973.tb03014.x. PMID 4777251.
- Miller RJ. Mitochondria – the kraken wakes!. Trends in Neurosci., 1998, roč. 21, čís. 3, s. 95–97 doi=10.1016/S0166–2236(97)01206–X.
- Siegel GJ, Agranoff BW, Fisher SK, Albers RW, Uhler MD. Basic Neurochemistry. [s.l.] : [s.n.], 1999. ISBN 0-397-51820-X.
- Green DR. Apoptotic pathways: the roads to ruin. Cell., september 1998, roč. 94, čís. 6, s. 695–8. DOI: 10.1016/S0092-8674(00)81728-6. PMID 9753316.
- OH-HAMA, T. Evolutionary consideration on 5-aminolevulinate synthase in nature. Orig Life Evol Biosph., august 1997, roč. 27, čís. 4, s. 405–12. DOI: 10.1023/A:1006583601341. PMID 9249985.
- ROSSIER, M. F. T channels and steroid biosynthesis: in search of a link with mitochondria. Cell Calcium., 2006, roč. 40, čís. 2, s. 155–64. DOI: 10.1016/j.ceca.2006.04.020. PMID 16759697.
- Richter C, Park J, Ames BN. Normal Oxidative Damage to Mitochondrial and Nuclear DNA is Extensive. PNAS, september 1988, roč. 85, čís. 17, s. 6465–6467. DOI: 10.1073/pnas.85.17.6465. PMID 3413108.
- http://www.circuitblue.com/biogerontology/mito.shtml
- Bender A, Krishnan KJ, Morris CM, Taylor GA, Reeve AK, Perry RH, Jaros E, Hersheson JS, Betts J, Klopstock T, Taylor RW, Turnbull DM. High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat Gen., 2006, roč. 38, s. 515–517. DOI: 10.1038/ng1769. PMID 16604074.
- GRAY, M. W.. The endosymbiont hypothesis revisited. International Review of Cytology, 1992, roč. 141, s. 233–357. ISSN 0074-7696. PMID 1452433. (po anglicky)
- EMELYANOV, Victor V.. Mitochondrial connection to the origin of the eukaryotic cell. European Journal of Biochemistry, apríl 2003, roč. 270, čís. 8, s. 1599–1618. DOI: 10.1046/j.1432-1033.2003.03499.x. PMID 12694174. (anglicky)
- HAMPL, Vladimír. Eukaryogeneze (PROTISTOLOGIE 2008) [online]. . Dostupné online.
- MARTIN, William; MÜLLER, Miklós. The hydrogen hypothesis for the first eukaryote. Nature, marec 1998, roč. 392, čís. 6671, s. 37–41. DOI: 10.1038/32096. PMID 9510246. (po anglicky)
- MOREIRA1, David; LÓPEZ-GARCÍA, Purificación. Symbiosis between methanogenic archaea and delta-proteobacteria as the origin of eukaryotes: the syntrophic hypothesis. Journal of Molecular Evolution, november 1998, roč. 47, čís. 5, s. 517–30. ISSN 0022-2844. DOI: 10.1007/PL00006408. PMID 9797402.
- Hydrogenosomes and Mitosomes: Mitochondria of Anaerobic Eukaryotes. Redakcia Jan Tachezy. Berlín : Springer, 2008. ISBN 978-3-540-76732-9. (po anglicky)