Deoxyribonukleová kyselina
Deoxyribonukleová kyselina alebo dezoxyribonukleová kyselina alebo kyselina de (z) oxyribonukleová, skr. DNK alebo najčastejšie DNA (angl. deoxyribonucleic acid) je prírodný polymér zložený z deoxyribonukleotidov. Spolu s kyselinou ribonoukleovou (RNA) patrí medzi nukleové (jadrové) kyseliny. Nukleové preto, lebo boli objavené v bunkovom jadre (lat. nucleus).

DNA je nositeľkou genetickej informácie bunky, riadi rast, delenie a regeneráciu bunky. Väčšinou je DNA v bunke uložená ako dvojzávitnicová špirála, ktorej vlákna majú navzájom opačnú orientáciu fosfodiesterových väzieb (sú antiparalelné). Dvojzávitnica DNA je veľmi tenká, ale zároveň dosahuje veľkú dĺžku, ktorá mnohonásobne presahuje dĺžku celej bunky. Preto je v bunke veľmi poohýbaná a zvinutá. Prokaryotické organizmy, mikroorganizmy bez bunkového jadra, majú iba jednu pre život nevyhnutnú molekulu DNA v bunke. Je umiestnená voľne v cytoplazme a jej konce sú väčšinou spojené. Eukaryotické organizmy, organizmy s jadrom, majú hlavnú časť DNA v jadre, kde tvorí niekoľko oddelených molekúl, spravidla s voľnými koncami. Okrem hlavnej DNA, označovanej ako chromozomálna DNA, majú organizmy menšie molekuly DNA uložené v bunkových organelách,[1] alebo v krátkych do kruhu uzavretých úsekoch umiestnených v cytoplazme, tzv. plazmidoch. Genetická informácia zapísaná v DNA sa realizuje prostredníctvom dvoch základných krokov, a to transkripcie a translácie.
Základné zložky DNA, jej monoméry, sa nazývajú deoxyribonukleotidy alebo všeobecne nukleotidy. Každý nukleotid pozostáva z troch zložiek:
- fosfátového zvyšku kyseliny fosforečnej (PO43−),
- molekuly deoxyribózy,
- dusíkatej bázy. Dusíkaté bázy tvoriace štruktúru DNA sú štyri: adenín (A), guanín (G), cytozín (C) a tymín (T). Spájajú obe vlákna dvojzávitnice DNA pomocou vodíkových väzieb, pričom platí princíp tzv. komplementarity báz. Komplementarita znamená, že adenín sa v DNA prednostne páruje s tymínom z druhého vlákna dvojzávitnice a guanín s cytozínom.[2]
Poradie jednotlivých dusíkových báz je kľúčové v prenose genetickej informácie. Funkčné úseky DNA, ktoré dokážu určité enzýmy prepísať do RNA, sa nazývajú gény.
Význam
Všetky v súčasnosti žijúce organizmy okrem sporných útvarov na hranici medzi živým a neživým potrebujú na svoje množenie DNA. V DNA je totiž obsiahnutá informácia na tvorbu základných stavebných komponentov bunky – proteínov a prostredníctvom DNA je táto informácia dlhodobo skladovateľná a replikovateľná. DNA má každý jedinec spomedzi bunkových foriem života a niektoré vírusy. Ďalšie vírusy – retrovírusy, v sebe neobsahujú DNA iba RNA, ale aj táto RNA musí byť na zmnoženie vírusovej častice najprv prepísaná v hostiteľskej bunke do DNA enzýmom zvaným reverzná transkriptáza.[3] Niektoré vírusy ale uskutočňujú kopírovanie svojej genetickej informácie len podľa RNA a DNA forma sa v ich životnom cykle nevyskytuje.
DNA je potrebná na množenie buniek, ich rast, špecializáciu v určitej funkcii (diferenciáciu), signalizáciu, ale aj na základné životné pochody v bunkách, ktoré nemusia vykonávať nijakú z predtým spomenutých aktivít. V každej bunke totiž pod vplyvom nepriaznivých vonkajších faktorov, ale aj samovoľnými procesmi časom jej stavebné a funkčné makromolekuly zanikajú. Preto musia byť neustále tvorené nové molekuly podľa vzoru zapísaného v DNA. Bunky, ktoré neobsahujú DNA, alebo podľa ich DNA nie je možné vytvárať proteíny (DNA je v kondenzovanom stave), majú obmedzenú životnosť. Takéto bunky sa nemôžu replikovať a tvorba ďalších takýchto buniek je závislá na bunkách, ktoré majú funkčnú – transkripčne aktívnu DNA. Neplatí, že každá bunka, ktorá má transkripčne aktívnu DNA, je schopná sa deliť, no je to jeden z nevyhnutných predpokladov. Keďže DNA je pre život buniek a organizmov z dlhodobého hľadiska nevyhnutná, v priebehu evolúcie sa v organizmoch vyvinuli mechanizmy, ako v prípade poškodenia DNA toto poškodenie opraviť.
Genetickú informáciu obsiahnutú v DNA, ani množenie a opravy poškodenej DNA, však nie je možné realizovať bez už existujúcich proteínov. Proteíny a DNA sú od seba vo svojom vzniku a šírení vzájomne závislé.
Vedci však uvažujú, že prvé živé organizmy na Zemi obsahovali ako základnú genetickú informáciu RNA[4] a len prostredníctvom RNA sa dokázali rozmnožovať. Neskôr boli vytlačené efektívnejšími a odolnejšími DNA formami života.
História výskumu

Nukleové kyseliny objavil v roku 1869 švajčiarsky lekár Friedrich Miescher. Jemu sa ako prvému podarilo z jadier bielych krviniek hnisu izolovať hmotu, ktorú nazval nukleín.[5] Vzorka však nebola dosť čistá na to, aby sa mohla ďalej skúmať. Phoebus Levene začiatkom 20. storočia zistil, že DNA sa skladá z cukrov, fosfátov a báz.[6] Jej funkcia v prenose genetickej informácie však dlho nebola známa. Jedna teória totiž hovorila, že genetická informácia sa prenáša vo forme špecializovaných bielkovín a nie DNA.[7] Až v roku 1943 priniesol slávny Averyho-MacLeodov-McCartyho experiment prvý dôkaz o úlohe DNA v dedičnosti. Oswald Avery, Colin Munro MacLeod a Maclyn McCarty sériou pokusov s transformáciou pneumokokov zistili, že DNA je genetickým materiálom buniek.[8] Ďalším dôkazom bol Hersheyho-Chaseovej experiment v roku 1952.
Erwin Chargaff so svojimi študentmi na základe analýz rôznych vzoriek DNA zistil, že pomer adenínu k tymínu a pomer guanínu a cytozínu je v nich rovnaký. Vzájomný pomer dvojíc adenín-tymín k dvojici guanín-cytozín bol rozdielny. Chargaff považoval tieto výsledky za významné, ale nevedel pre ne nájsť vysvetlenie.[7]
Odhalenie trojrozmernej štruktúry DNA bolo v jej výskume veľmi významným míľnikom. Objav dvojzávitnicovej štruktúry sa spája s menami Jamesa Watsona a Francisa Cricka. Pred vypracovaním Watsonovho-Crickovho modelu už bolo známe, že DNA je tvorená monomérmi – nukleotidmi, ktoré sa líšia svojimi dusíkatými bázami. Dlho však nebolo jasné, ako sú nukleotidy usporiadané za sebou do polynukleotidového reťazca, či v ich usporiadaní existujú nejaké zákonitosti, koľko polynukleotidových reťazcov tvorí jednu molekulu DNA a aká je priestorová orientácia reťazca. Dôležitý pokrok v poznaní štruktúry DNA urobil S. Furberg, ktorý v roku 1949 objavil, že rovina dusíkatej bázy v nukleotide je takmer kolmá na rovinu, v ktorej leží väčšina atómov cukornatej zložky. Výskumná skupina Alexandra Todda v roku 1951 zistila, že nukleotidy sú spojené fosfodiesterovou väzbou spájajúcou 5' atóm cukru s 3' atómom cukru nasledujúceho nukleotidu. Problémom priestorového usporiadania atómov v DNA sa však nezaoberali.[7] Správny priestorový model DNA predstavili až Watson a Crick v roku 1953 v časopise Nature. Pri tomto objave vychádzali z röntgenovej difrakčnej analýzy, ktorú rok pred ich publikáciou vykonali Rosalind Elsie Franklinová a Raymond Gosling a tiež publikovali v rovnakom čísle časopisu Nature. Crick predložil v roku 1957 sériu pravidiel označovaných ako centrálna dogma molekulárnej biológie. Tieto pravidlá popisujú vzťahy medzi DNA, RNA a proteínmi.[9] V nasledujúcom roku Meselsonov–Stahlovv experiment predstavil spôsob, akým je DNA v bunkách množená.[10] Na začiatku 60. rokov Har Gobind Khorana, Robert W. Holley a Marshall Warren Nirenberg rozlúštili genetický kód.[11] V roku 1980 dostali Walter Gilbert a Frederick Sanger Nobelovu cenu za objav metódy, ktorou možno určiť poradie báz v DNA.[12]
Štruktúra DNA
Primárna štruktúra

DNA je polymér, čiže molekula zložená z mnohých jednotiek zvaných monoméry. Monoméry DNA sa nazývajú deoxyribonukleotidy, niekedy sa požíva aj názov nukleotidy, čo je súhrnný názov pre monoméry DNA – deoxyribonukleotidy a monoméry RNA – ribonukleotidy. Do polynukleotidového reťazca sú deoxyribonukleotidy spojené esterovou (fosfodiesterovou) väzbou. Deoxyribonukleotid tvorí
- zvyšok kyseliny trihydrogenfosforečnej
- deoxyribonukleozid
- cukor deoxyribóza
- dusíkatá báza
Dusíkatých báz je známych niekoľko, v DNA sa však uplatňujú len štyri: adenín (skratka A), tymín (T), guanín (G) a cytozín (C). Spojením adenínu s deoxyribózou vzniká deoxyadenozín (skratka dA), spojením tymínu s deoxyribózou deoxytimidín (dT), atď. Keďže kyselina fosforečná a deoxyribóza sú spoločné zložky pre všetky nukleotidy, jednotlivé nukleotidy sa od seba odlišujú len bázou. Práve bázy sú zodpovedné za kľúčovú schopnosť DNA zaznamenávať a prenášať genetickú informáciu. Deoxyribózová a kyselinová zložka slúžia na to, aby držali bázy vo vhodných polohách a vzdialenostiach. Tieto dve zložky tvoria takzvanú pentózafosfátovú kostru DNA.[13]
Väzba sa vytvára medzi zvyškom kyseliny trihydrogenfosforečnej na 5. uhlíkovom atóme pentózy jedného nukleotidu a hydroxylovou skupinou viazanou na 3. uhlíkovom atóme pentózy susedného nukleotidu. Sú teda viazané 3',5'-fosfodiesterovou väzbou[13] (čítaj tri s čiarou päť s čiarou fosfodiesterovou väzbou),[2] zapisovanou aj ako 5'-3'-fosfodiesterová väzba.[12] Na vlákne DNA s nespojenými koncami (lineárnej molekule) rozlišujeme dva konce: na 5' konci je vlákno ukončené fosfátom a na 3' konci je vlákno ukončené hydroxylovou -OH skupinou. Na rozdiel od primárnej štruktúry bielkovín, na ktorej sa podieľa 21 aminokyselín, primárna štruktúra NK obsahuje iba štyri nukleotidy, pričom pomer adenínu s tymínom (A : T) a guanínu s cytozínom (G : C) je rovnaký 1 : 1.[2] Niektoré z báz (u človeka odhadom asi 1 %) môžu byť metylované. U zvierat je toto percento i niekoľkonásobne vyššie (napríklad u makakov až 6 %).
Poradie nukleotidov – sekvencia DNA – sa udáva vždy pre jedno z dvoch vlákien vypísaním jednotlivých nukleotidov od 5' konca k 3' koncu. Každý nukleotid sa označuje jednopísmenovou skratkou dusíkatej bázy, ktorú obsahuje. Veľkosť genómu jednotlivých organizmov alebo organel sa udáva počtom bázových párov (skratka bp), ktoré obsahuje ich DNA.
Komplementárnosť


Dôležitá vlastnosť štruktúry DNA je komplementárnosť jednotlivých nukleotidov. To znamená, že dve špecifické dusíkaté bázy dvoch vlákien alebo jedného dostatočne ohnutého vlákna sú, pokiaľ sa nachádzajú oproti sebe, schopné utvoriť chemické väzby. Podľa pravidla, ktoré sa nazýva Watsonovo-Crickovo podľa jeho objaviteľov, sa páruje vždy purínová s pyrimidínová báza. V prípade DNA sa teda adenín páruje s tymínom a guanín s cytozínom.[13] Guanín s cytozínom sa viažu tromi vodíkovými väzbami, a adenín s tymínom zase dvomi vodíkovými väzbami.[2] Keďže väzba adenín-tymín je kvôli menšiemu počtu vodíkových mostíkov slabšia, veľké množstvo adeníno-tymínovych párov sa nachádza v miestach, kde je potrebné, aby sa dvojzávitnica ľahko rozdelila na dve jednotlivé vlákna (napr. v tzv. replikačných počiatkoch). Spárovaným komplementárnych báz sa utvorí typická dvojreťazová štruktúra DNA. Takto sa uchováva v každom z dvoch vlákien vlákne rovnaká informácia, iba s rozdielom, že ide o vzájomný negatív. Výhoda takejto štruktúry je v tom, že ak sa jedno vlákno dvojzávitnice DNA poškodí, informácia v ňom obsiahnutá ešte nie je úplne stratená a možno ju obnoviť podľa druhého vlákna.
Reťazce sú vzhľadom na seba postavené v opačnom smere. Kým jeden reťazec je orientovaný v smere 5'→ 3', druhý bude orientovaný v smere 3'→ 5'. Oddeliť od seba tieto vlákna možno zahriatím na vysokú teplotu, zmenou pH, zmenou iónovej sily roztoku a prítomnosťou niektorých organických látok, napríklad močoviny. Tento jav, rozpadnutie dvojvlákna na jednotlivé vlákna, sa nazýva denaturácia DNA. Pri teplotnej denaturácii je možné postupným ochladzovaním roztoku dosiahnuť opätovné spárovanie báz v oboch reťazcoch a obnovenie pôvodnej štruktúry.[14]
Watson-Crickovo pravidlo o párovaní báz je základný spôsob párovania, na ktorom je postavený celý prenos genetickej informácie. Existujú však aj iné spôsoby párovania, ktoré vedú k vzniku trojreťazových a štvorreťazových molekúl DNA. Taktiež existuje dvojreťazová DNA, ktorej reťazce nie sú antiparalelné, ale paralelné. V takomto prípade sa bázy párujú tzv. obráteným Watsonovo-Crickovým párovaním, ktorého zvláštnosťou je, že dovoľuje aj párovanie dvoch rovnakých báz, napríklad guanínu s guanínom.[13]
Hoogestenovo párovanie báz je párovanie za nezvyčajných fyzikálnochemických podmienok, pri ktorom sa vytvára spojenie medzi troma bázami súčasne. Pri neutrálnom pH však takéto párovanie nie je stabilné. Existuje aj párovanie štyroch báz – tetrád.[13]
Sekundárna štruktúra

Sekundárnou štruktúrou označujeme priestorové usporiadanie polynukleotidového reťazca. Dvojvlákno spojené vodíkovými väzbami komplementárnych nukleových báz má najčastejšie tvar pravotočivej dvojzávitnice, α-helixu.[2] Forma stočení nie je vždy za každých podmienok rovnaká. Pri vysokom obsahu vody zaujíma DNA štruktúru B formy. Je to forma pravotočivej dvojzávitnice, v ktorej sú roviny báz kolmé ku kostre dvojzávitnice. V ideálnej B-forme má molekula 10 párov báz na jednu otáčku dvojzávitnice[15] a dva susediace bázové páry pri kolmom pohľade zhora zvierajú uhol 36°. Roviny prechádzajúce bázovými pármi sú navzájom rovnobežné.[14] V B forme je DNA v skúmavke (in vitro), ale v živých organizmoch (in vivo) za určitých podmienok prirodzene prechádza aj na inú formu. B forma je svojim tvarom často prirovnávaná k točitému schodisku.
A forma je pravotočivá. Obsahuje 11,6 komplementárnych párov na jeden závit, čo naznačuje, že stúpanie helixu je pozvoľnejšie ako v B forme.[14] Priemer vlákna je 2,3 nm. Rovina prechádzúca párom báz je asi o 20° vychýlená z polohy kolmej na os vlákna. Ľavotočivá Z forma má 12 komplementárnych párov na jeden závit; priemer vlákna je 1,8 nm.
Povrch dvojzávitnice sa vyznačuje dvoma žliabkami odlišujúcimi sa navzájom šírkou aj hĺbkou. Rozpoznávame menší žliabok so šírkou 0,6 nm a väčší žliabok široký 1,2 nm. V oboch žliabkoch sa nachádzajú atómy schopné vytvárať vodíkové väzby s atómami proteínov. Vo väčšom žliabku je ich viac než v menšom. Konformácie A, B a Z sa od seba odlišujú aj tvarom týchto žliabkov – kým u konformácie A je veľký žliabok veľmi úzky a hlboký, v konformácii B je širší a menej hlboký a v konformácii Z sploštený na povrchu.[13]
Terciárna štruktúra
Pod terciárnou štruktúru rozumieme priestorové usporiadanie dvojzávitnice. Tá sa môže stočiť do superhelixu spôsobom tzv. nadzávitnicového vinutia. Takto zvinutá DNA sa nazýva superšpiralizovaná DNA.[2] Superšpiralizovaná môže byť dvojvláknová DNA, ktorá je uzavretá do kruhu, alebo lineárna dvojvláknová DNA, ktorej konce sú pripevnené o podklad. Terciárnu štruktúru zabezpečujú enzýmy topoizomerázy. Dvojzávitnica bez terciárnej štruktúry sa nazýva relaxovaná DNA.[13]
Druhy DNA
- jednovláknová DNA (skratka ssDNA z anglického single-stranded DNA) – len jedno vlákno DNA, ktorému chýba komplementárne vlákno. V živých bunkách sa bežne nevyskytuje, no môžu ju ako svoju genetickú informáciu obsahovať niektoré druhy vírusov.
- dvojvláknová DNA (skratka dsDNA z anglického double-stranded DNA) – DNA zložená z obidvoch vlákien spojených navzájom vodíkovými mostíkmi. V tejto forme je DNA uložená v živých bunkách a túto formu opúšťa len na krátkych úsekoch a na krátku dobu, napríklad počas replikácie či transkripcie.
- komplementárna DNA (skratka cDNA z anglického complementary DNA) – DNA, korá vznikla prepísaním mRNA enzýmom reverzná transkriptáza do DNA.
- genómická DNA (skratka gDNA z anglického genomic DNA) – chromozomálna DNA
- chloroplastová DNA – (skratka cpDNA z anglického chloroplast DNA) – DNA v chloroplastoch, organelách v bunkách rias a rastlín.
- mitochondriálna DNA – (skratka mtDNA z anglického mitochondrial DNA) – DNA v mitochondriách, organelách väčšiny eukaryotických organizmov.
DNA viažúce proteíny
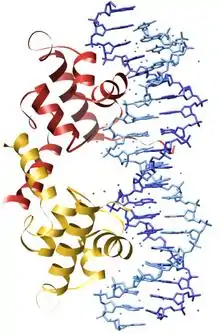
DNA by nebola schopná sa replikovať a správne vykonávať svoje funkcie bez proteínov, ktoré sa na ňu v živých organizmoch viažu. Tieto proteíny sa na DNA viažu tzv. nekovalentnými väzbami – vodíkovými mostíkmi, iónovými väzbami alebo hydrofóbnymi interakciami väčšinou v oblasti väčšieho žliabku. Každé z týchto spojení je slabé, preto proteín musí vytvárať takýchto väzieb viacero, aby sa dokázal na DNA udržať po dlhšiu dobu. Pri svojom nadviazaní proteíny nemenia jej chemické zloženie, ale napriek tomu ovplyvňujú jej vlastnosti.[14] Môžu napríklad spôsobiť ohyb molekuly DNA, čo sa uplatňuje pri zbalení molekuly DNA do kompaktnejších štruktúr, ale aj pri plnení funkcie DNA (napríklad proteín TBF ohnutím molekuly DNA umožňuje, aby na ňu nasadli ďalšie proteíny slúžiace na spustenie transkripcie). Obvykle sa na DNA neviaže celý proteín, len jeho takzvaná DNA-väzobná doména. Rozpoznávací význam má väzba jednotlivých aminokyselín proteínu na bázy DNA, väzba na fosfodiesterovú kostru len upravuje pozíciu proteínu vzhľadom na bázy. DNA-väzobné proteíny môžu rozoznávať rôzne bázové páry bez toho, aby rozrušili vodíkové mostíky medzi nimi a tým spôsobili "rozpletenie" molekuly. Schopnosť proteínu viazať sa len na bázy zoradené v správnom poradí (špecifickom pre daný proteín), nazývame sekvenčná špecifita proteínu. Inými slovami, DNA poradím svojich báz v jednom úseku dovoľuje na tento úsek nasadnúť len proteínu z určitej skupiny. Niektoré DNA-viažúce proteíny sa môžu na DNA nadviazať, len ak sú na nej už prítomné iné proteíny.
Proteíny nadviazané na DNA môžu umožňovať transkripciu daného úseku, zosilňovať ju, alebo naopak tlmiť až úplne zastaviť. Sú preto kľúčové pre reguláciu génov a sú zodpovedné za to, aby sa správny gén prejavoval v bunke v správnom čase. DNA väzobné proteíny, ktoré sa viažu na DNA len dočasne a špecificky zapínajú a vypínajú prepisovanie určitého génu, sú napojené na signálne dráhy v bunke.
Spôsob výskytu DNA
Miesto výskytu a spôsob uloženia DNA má veľké odlišnosti u jednotlivých domén živých organizmov.
Prokaryotická DNA
V prokaryotickej bunke nie je genetická informácia oddelená od zvyšku cytoplazmy jadrovou membránou. Spravidla, nie však vždy, je uzavretá do kruhu a označuje sa ako nukleoid alebo kruhový chromozóm. Okrem chromozómu vlastní takmer každá prokaryotická bunka niekoľko plazmidov, ktoré sa definujú ako doplnkové genetické elementy. Obsahujú gény, ktoré nie sú potrebné na prežitie bunky, ale obsahuje napr. gény nesúce informácie o rezistencii voči antibiotikám alebo pre tvorbu toxínov.
Eukaryotická DNA

Jadrová DNA eukaryotov na rozdiel od prokaryotického chromozómu je chránená od metabolických procesov prebiehajúcich v cytoplazme dvojitou jadrovou membránou. Členená je spravidla do viacerých oddelených molekúl. V interfáze, vo fáze medzi deleniami bunky, je komplex DNA a bielkovín označovaný ako chromatín. Chromatín sa skladá z nukleozómov, čo sú asociácie 8 molekúl špeciálnych bielkovín – histónov, ktoré sú 2,5-krát obtočené molekulou DNA. V čase bunkového delenia sa chromatín preorganizuje na chromozómy, ktoré sa počas delenia správajú ako samostatné elementy a sú viditeľné aj svetelným mikroskopom. Eukaryotická DNA je dvojvláknová a lineárna, len výnimočne kruhová. Proteíny, s ktorými je DNA asociovaná, napomáhajú jej špiralizácii a majú aj regulačnú úlohu pri realizácii genetickej informácie.
Mimojadrová DNA
Dve membránové organely eukaryotických buniek, mitochondrie a chloroplasty, obsahujú vlastné molekuly DNA. Tieto molekuly sú zväčša kruhové a neobsahujú históny. Preto sa predpokladá že tieto organely vznikli pohltením baktérií na základe symbiózy. Okrem mitochondrií a chloroplastov možno v eukaryotických organizmoch nájsť DNA tiež v nukleomorfe, čo je pozostatok jadra iného eukaryotického organizmu, ktoré bunka v priebehu evolúcie pohltila. Jednoduché eukaryotické organizmy, napríklad kvasinky, majú tiež plazmidy, podobne ako baktérie.
Funkcie DNA
Najznámejšou úlohou DNA je uchovávať a odovzdávať genetickú informáciu o poradí aminokyselín v bielkovinách, ktorá je zapísaná v jej primárnej štruktúre. Na základe primárnej štruktúry sa podľa oboch vlákien dvojzávitnice DNA vytvoria dve nové dvojzávitnice DNA (replikácia), alebo sa podľa jedného z nich vytvorí jedna jednovláknová RNA (transkripcia). Vzniknuté vlákno RNA môže niesť informáciu o tvorbe peptidového reťazca a podľa tejto informácie sa vytvorí nový peptid v ribozómoch. RNA, podľa ktorej sa peptid vytvára, sa nazýva mediátorová RNA. Existujú však aj ďalšie druhy RNA, ktoré sú pre život rovnako nevyhnutné a to takzvané funkčné RNA, čo sú RNA, ktoré sa do bielkovín neprepisujú a plnia určitú úlohu. Medzi funkčné RNA patrí ribozómová RNA a transferová RNA[16] – bez účasti oboch týchto RNA by translácia nemohla prebiehať. Matrice pre funkčnú RNA sú tiež dané primárnou štruktúrou DNA.
Pri transkripcii DNA do RNA vznikajú náhodné chyby, ktoré môžu byť neškodné, ale tiež môžu úplne zmeniť funkciu produktu. Okrem toho však dochádza u eukaryotických organizmov k zámerným zmenám RNA (RNA processing a RNA editing), ktorej primárna štruktúra už potom nezodpovedá matrici v DNA a preto jej nemusí zodpovedať ani výsledný produkt. Odvodiť preto všetky produkty v organizme, u ktorého je úplne známa sekvencia jeho DNA, je oveľa zložitejšie, než sa na prvý pohľad zdá.
Genetický kód
Ako informácia pre zaradenie určitej aminokyseliny do vznikajúceho peptidového reťazca slúžia v DNA (a takisto v jej transkripte, RNA) tri vedľa seba ležiace nukleotidy. Táto trojica nukleotidov – tzv. triplet sa nazýva kodón. Keďže bázy sú štyri a každá z nich sa môže nachádzať na jednej z troch pozícií, malo by existovať 64 rôznych kodónov. Z nich však tri nekódujú nijakú aminokyselinu a ich prítomnosť v RNA naznačuje, že v tomto mieste je potrebné syntézu proteínu ukončiť. Aj tak však zostáva 61 rôznych kodónov, pričom poznáme len 21 aminokyselín, ktoré organizmy používajú pre proteosyntézu. Rozličné kodóny však môžu kódovať tie isté aminokyseliny. Len metionín, tryptofán a selenocysteín majú svoje kodóny jedinečné. Ostatné aminokyseliny sú kódované dvoma, štyrmi až šiestimi rozličnými kodónmi. Kvôli tejto vlastnosti sa genetický kód nazýva degenerovaný.[12]
Topológia DNA
Najvýznamnejšími úsekmi molekuly DNA sú gény. Ako gén možno označiť sekvenciu, ktorá je prepisovateľná do RNA a plní v organizme určitú úlohu. Aby gény mohli byť prepisované a aby sa to odohrávalo v správny čas, obsahujú gény takzvané regulačné sekvencie. Tie nie sú súčasťou polypeptidového reťazca (neprekladajú sa), ale umožňujú regulačným proteínom a proteínom sprostredkujúcim priamo proces transkripcie nájsť kódujúce sekvencie v DNA. Využívajú sa pritom DNA-väzobné proteíny viazané na špecifické sekvencie. Gény sú zoradené lineárne za sebou, ale vzdialenosti medzi jednotlivými génmi bývajú rôzne, u prokaryotov všeobecne menšie než u eukaryotov. V medzerách mimo regulačných sekvencií sa nachádzajú úseky DNA, ktorých funkcia väčšinou nie je celkom alebo vôbec známa. Niektoré úseky tvoria mnohopočetné opakovania (repetície), iné sú jedinečné. Každá molekula DNA musí obsahovať tiež minimálne jeden replikačný počiatok, čiže miesto, z ktorého sa zahajuje jej replikácia.
Replikácia DNA
| Priebeh replikácie DNA u eukaryotov s názvami príslušných enzýmov |
|---|
DNA polymeráza α
Topoizomeráza
Primáza
Helikáza
SSB proteíny
DNA polymeráza δ
|
Na to aby mohla DNA plniť svoju funkciu musí byť schopná zdvojenia sa, čo sa uskutočňuje replikáciou – kopírovaním.
Pri vlastnej syntéze DNA sa obidve vlákna pôvodnej molekuly začnú od seba vzďaľovať a rozpletať, čo zabezpečí enzým helikáza. Rozpletením dvojzávitnice vzniká „replikačná bublina“. Voľné nukleotidy začne enzým DNA-polymeráza podľa princípu komplentarity prikladať k pôvodnému (matricovému) vláknu DNA (3’ → 5’). Oproti druhému vláknu (5’→ 3’) syntetizuje DNA-polymeráza iba krátke fragmenty (tzv. Okazakiho fragmenty), ktoré následne spája enzým DNA ligáza. Aby nový nukleotid mohol byť zaradený, musí byť v tzv. aktivovanej forme, musí mať na 5' uhlíku deoxyribózy nadviazané tri fosfátové skupiny. Navzájom reagujú deoxynukleotidtrifosfáty, všeobecne dNTP (dATP, dTTP, dGTP, dCTP), pričom sa pri reakcii odštepuje PPi pyrofosfát (spojené dve fosfátové skupiny). Týmto spôsobom z jednej pôvodnej DNA vzniknú dve molekuly, z toho každá má jedno vlákno pôvodné a jedno vlákno nové. Je to tzv. semikonzervatívny spôsob.
Podmienky pre aktivitu DNA-polymerázy:
- V reakčnom prostredí musia byť ako substrát prítomné všetky štyri dNTP.
- Syntéza DNA nemôže vznikať de novo, DNA polymerázy vyžadujú prítomnosť voľnej 3’-OH na reťazci, ktorý sa má predlžovať. Molekula poskytujúca túto skupinu sa označuje ako primer. Na úplnom začiatku syntézy je to krátka jednovláknová RNA, dlhá približne 15 nukleotidov, ktorá sa komplementárne napojí na matricovú DNA.
- Pre funkčnosť a presnosť DNA polymerázy sú potrebné dvojmocné katióny, najčastejšie Mg2+ ako konfaktor. Aktívne miesto DNA polymerázy obsahuje dva katióny Mg2+ a každý je komplexne viazaný s šiestimi atómami kyslíka, ktoré poskytuje voda, COO- skupina aminokyseliny asparágovej zaradenej v peptidovom reťazci enzýmu, fosfátové skupiny dNTP a -OH skupina primeru. Takto nadviazané a stabilizované nukleotidy sa naviažu a odbúra sa pritom už spomínaný pyrofosfát.
- Potrebný je aj templát (vzor). Ide o jednovláknový úsek DNA podľa ktorého sa na základe komplementarity zaraďujú nukleotidy v syntetizovanom vlákne.
Zmeny v DNA
Pri replikácii DNA občas (asi v 1 prípade z 107) dochádza k zapojeniu chybného nukleotidu. Väčšina týchto chýb je opravená buď samotnou DNA-polymerázou, alebo systémom opravy chybného párovania báz, ale niektoré pretrvávajú. Tieto chyby, ktoré môžu byť pri ďalšej replikácii prenášané na novovzniknuté DNA a s nimi do dcérskych organizmov, sa nazývajú mutácie. Mutácie vznikajú aj v období mimo replikácie DNA, napríklad pod vplyvom ionizujúceho žiarenia, či pôsobením voľných radikálov. Ale aj DNA umiestnená v ideálnych podmienkach mimo všetkých bežných škodlivých vplyvov, podlieha kontinuálnemu poškodzovaniu. Medzi tieto druhy poškodenia patrí depurinácia (strata purínových báz), deaminácia – zmena cytozínu na uracil, a ďalšie. Mutácie môžu mať mnohé podoby. Môžu postihnúť naraz aj dlhé sekvencie či dokonca celú molekulu DNA alebo genóm. Môže napríklad dôjsť k zlomu v jednom alebo naraz v oboch vláknach DNA, pričom druhý prípad je oveľa závažnejší.
Keby neexistoval opravný mechanizmus mutácií, nahromadené mutácie by vo väčšine prípadov rýchlo spôsobili zánik organizmu. Našťastie je väčšina mutácií rozpoznávaná a opravovaná reparačnými mechanizmami. Tieto zahŕňajú enzýmy zo skupiny endonukleáz, ktoré (poškodenú) DNA štiepia a ligáz, ktoré zlepujú rozštiepenú DNA dokopy. DNA je preto dynamická štruktúra, v ktorej neustále prebiehajú poškodenia a opravy.
Občas ale niektorá mutácia opravnému mechanizmu unikne. Pokiaľ daná mutácia spôsobí zmenu zmyslu životne dôležitého génu, bunka zahynie. Mutácia však môže byť aj synonymná, tzn. jeden nukleotid sa zmení, ale zhodou okolností aj so zmeneným kodónom priradí translačný aparát k príslušnému kodónu tú istú aminokyselinu ako pôvodne (pretože pre niektoré aminokyseliny existuje až do 6 rôznych kodónov[12]). Menej škodlivé, neškodné a priaznivé mutácie sú odovzdávané potomstvu bunky. DNA je chemicky všade rovnaká a neopravené mutácie sa v nej vyskytujú úplne náhodne. No keďže mutácie meniace zmysel životne dôležitých génov vedú k smrti bunky, potomstvu sa budú odovzdávať najčastejšie mutácie v miestach DNA, ktoré nebudú mať pre bunku veľký význam.
Realizácia genetickej informácie
Realizácia genetickej informácie na molekulovej úrovni prebieha ako dvojstupňový proces:
- Transkripcia: prepis genetickej informácie z DNA do mRNA → syntéza mRNA
- Translácia: preklad genetickej informácie z poradia nukleotidov na mRNA do poradia aminokysleín v polypeptidovom reťazci → syntéza bielkovín
Transkripcia
Je to syntéza molekúl RNA podľa matrice DNA. Tento proces využíva komplementaritu báz medzi DNA a RNA. Pri transkripcii sa reťazce molekuly DNA čiastočne oddelia narušením vodíkových mostíkov, pričom iba jedno z vlákien potom slúži ako matrica pre syntézu RNA. Proces syntézy katalyzuje enzým RNA-polymeráza, ktorý na základe komplementarity báz pripája ribonukleotidy za vzniku RNA. Jednou z úloh RNA-polymerázy je aj urýchliť oddelenie novovznikajúcu RNA od jej matrice v podobe DNA. Začiatok transkripcie, miesto, kde sa RNA polymeráza viaže, ako aj koniec, v ktorom sa oddeľuje, tvoria špecifické sekvencie báz. Existuje viacero druhov RNA, ktoré takýmto spôsobom môžu vznikať. Proteíny sa môžu vytvárať len podľa takzvanej mediátorovej RNA – mRNA.
Translácia
Je zavŕšením realizácie genetickej informácie, ide o vlastnú syntézu bielkovín (proteosyntéza). Tento proces sa uskutočňuje na ribozómoch, časticiach nachádzajúcich sa v cytoplazme bunky. Translácie sa zúčastňuje jedna molekula mRNA, ktorá nesie vlastnú informáciu vo forme kodónov a veľa molekúl tRNA, na ktoré sa viaže aminokyselina a ktoré zároveň obsahujú antikodón, sekvenciu komplementárnu s kodónom na mRNA. Molekuly tRNA sa dočasne pripájajú antikodónmi ku kodónom na mRNA a tak prenášajú jednotlivé aminokyseliny zo zásoby voľných aminokyselín v cytoplazme bunky. Medzi jednotlivými aminokyselinami potom vznikajú peptidické väzby, čím vzniká polypeptidový reťazec – bielkovina.
Rekombinácia DNA
DNA všetkých organizmov na Zemi je po chemickej stránke rovnaká. Hociktorý voľný 5' koniec sa môže spojiť s hocijakým voľným 3' koncom. Preto je možné spojiť fragmenty DNA aj vysoko nepríbuzných druhov a takisto je možné aj spojenie DNA chromozomálnej a extrachromozomálnej. Rekombináciu DNA si netreba zamienať s pohlavným rozmnožovaním. Pri spojení dvoch pohlavných buniek nedochádza k vzájomnej rekombinácii ich DNA. DNA v každej pohlavnej bunke tvorí samostatné oddelené molekuly, ktoré sa síce v oplodnenom vajíčku dostanú spolu do jedného jadra, no fyzicky sa nespájajú ani nerekombinujú. Ale v oveľa neskoršej fáze ontogenézy, pri tvorbe vlastných pohlavných buniek jedinca, rekombinovať môžu – tento spôsob prirodzenej rekombinácie sa nazýva crossing over.
Umelou rekombináciou sú vedci schopní vyrobiť také sekvencie DNA, ktoré sa v prírode nevyskytujú. Takáto DNA sa nazýva rekombinantná. Samotné spojenie dvoch koncov molekuly však ešte nezaručuje, že daný úsek DNA bude možné transkribovať a že sa bude podľa neho bude v inom organizme tvoriť rovnaký produkt ako v pôvodnom organizme. Osud RNA po transkripcii je napríklad u prokaryotických organizmov iný než u eukaryotických. Takisto proteíny po skončení syntézy podliehajú u eukaryotov často ešte zložitým úpravám. Dosiahnuť preto, aby napríklad baktéria produkovala nejaký cicavčí proteín, je preto veľmi zložité a nestačí jednoducho vyštiepiť príslušný génu z DNA cicavcov a vložiť ho do DNA baktérie. Keďže baktérie napríklad nedokážu zostrihať z pre-mRNA intróny, pre expresiu génu v baktérii sa do nej musí vniesť cDNA daného proteínu.
Rekombinácia DNA má význam napríklad v medicíne. Vložením génov pre produkciu ľudských proteínov do mikroorganizmov je možné získavať vo veľkých množstvách čistý proteín. Ten sa potom umelo podá pacientovi, ktorého telo kvôli genetickej chorobe daný proteín neprodukuje. Tvorba rekombinantných organizmov však naráža na etické otázky, ako aj obavy z nepredvídateľných následkov rekombinácii či z úniku rekombinantných organizmov do prírody a ich nepriaznivých efektov na ekosystémy.
Zdroje
- Nukleové kyseliny [online]. Biopedia.sk, [cit. 2011-01-13]. Dostupné online.
- ALBERTS, Bruce; BRAY, Dennis; JOHNSON, Alexander, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter Základy buněčné biologie. Redakcia Miranda Brownová, Eleanor Lawrenceová, Valerie Nealová, Anne Vinnicombeová; preklad Arnošt Kotyk, Bohumil Bouzek, Pavel Hozák; ilustrácie Nigel Orme. 2.. vyd. Ústí nad Labem : Espero Publishing, © 1998. ISBN 80-902906-2-0. (česky)
Referencie
- DNA deoxyribonukleová kyselina [online]. Encyklopédia Vševěd, [cit. 2011-01-13]. Dostupné online. (česky)
- Nukleové kyseliny [online]. Biopedia.sk, [cit. 2011-01-13]. Dostupné online.
- FERENČÍK, Miroslav; ROVENSKÝ, Jozef; MAŤHA, Vladimír, Erika Jensen-Jarolim Imunológia a alergológia v základných heslách a termínoch. prvé. vyd. Bratislava : SLOVAK ACADEMIC PRESS s.r.o., 2006. ISBN 80-89104-82-7. S. 357.
- HOUSER, Pavel. RNA svět: Existoval vůbec někdy? [online]. Science world, 12.03.2002, [cit. 2011-01-25]. Dostupné online. (česky)
- Dahm R. Discovering DNA: Friedrich Miescher and the early years of nucleic acid research [online]. . S. 565–81. DOI:10.1007/s00439-007-0433-0
- Levene P,. The structure of yeast nucleic acid [online]. 1 December 1919. S. 415–24. Dostupné online.
- WATSON, James D.. Tajemství DNA. Preklad František Sládeček. prvé. vyd. Praha : Academia, 1995. 150 s. ISBN 80-200-0556-0. (česky)
- Avery O, MacLeod C, McCarty M. Studies on the chemical nature of the substance inducing transformation of pneumococcal types. Inductions of transformation by a desoxyribonucleic acid fraction isolated from pneumococcus type III [online]. . S. 137–158. Dostupné online. DOI:10.1084/jem.79.2.137
- Crick, F.H.C. On degenerate templates and the adaptor hypothesis (PDF). genome.wellcome.ac.uk (Lecture, 1955). Accessed 22 December 2006
- Oxford dictionary of biochemistry and molecular biology; revised edition. Ed. R. Cammack et al. New York : [s.n.], 2006. ISBN 0-19-852917-1.
- The Nobel Prize in Physiology or Medicine 1968 Nobelprize.org Accessed 22 December 06
- THOMPSON, James S.; THOMPSONOVÁ, Margaret W.. Klinická genetika. Preklad Viliam Izakovič. štvrté. vyd. Martin : Osveta, 1988. 440 s.
- ROSYPAL, Stanislav. Úvod do molekulární biologie. tretie. vyd. Brno : [s.n.], 1998. (česky)
- http://is.muni.cz/th/151360/prif_b/bc_MB.txt
- http://orion.chemi.muni.cz/zakladni_pojmy_z_biochemie/page0431.htm
- Nukleové kyseliny a proteosyntéza [online]. [Cit. 2011-02-09]. Dostupné online.