Apoptóza
Apoptóza je programovaná bunková smrť. Na bunku pôsobia signály (z vnútorného alebo vonkajšieho prostredia), ktoré postupne vedú k tomu, že bunka sama rozštiepi svoju DNA na fragmenty, rozloží vlastné proteíny a pomocou cytoskeletu sa rozpadne na viacero apoptotických teliesok, ktoré sú v extracelulárnom prostredí pohltené okolitými bunkami. V tom sa líši od nekrózy, kedy je bunka zničená „násilne“, jej obsah vytečie do okolia ostatných buniek a môže ich svojím obsahom neadekvátne ohroziť (výlevom mediátorov, ktoré mala vo vnútornom prostredí).
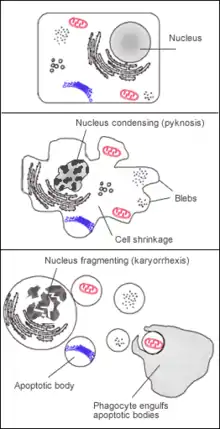
Apoptóza je veľmi dôležitá v embryonálnom vývoji (prsty na rukách sa zakladajú aj s blanami medzi prstami, ktoré neskôr odumrú apoptózou) i pri raste organizmu a jeho jednotlivých častí. Veľký význam má takisto pri rozpoznávaní nekontrolovateľne sa množiacich buniek. Organizmus má ochranné mechanizmy, ktoré dokážu takýmto bunkám príslušnou signalizáciou „vnútiť“ apoptózu.
Detekcia apoptózy
Apoptóza sa začína zmršťovaním bunky, pričom integrita cytoplazmatickej membrány ostáva zachovaná. Aktivácia kaspáz vedie k strate mitochondriálneho membránového potenciálu. Dochádza k zmenám intracelulárnej koncentrácie Ca2+. Lyzozomálne membránové pumpy strácajú svoju funkciu. Sú aktivované endogénne endonukleázy a bunka sa rozpadá na apoptotické telieska . Tieto bunkové zmeny sú merateľné metódou prietokovej cytometrie .
Rozptyl svetla bunkou
Apoptóza vo väčšine prípadov môže byť od nekrózy rozpoznaná na základe miery rozptylu svetla. Priamy rozptyl svetla (forward-angle light scatter, FSC) je spojený s priemerom bunky, bočný rozptyl (side-angle light scatter, SSC) je spojený so stavom vnútorných bunkových štruktúr . V prvých štádiách apoptózy sa bunka zmršťuje, zatiaľ čo membrána ostáva intaktná. Pri nekróze membrána bunky stráca integritu. V dôsledku týchto zmien pri prvých štádiách apoptózy sa znižuje FCS, a SSC sa zvyšuje, prípadne sa nemení. Pri nekróze sa FCS okamžite zvýši a SSC zníži. Výhodou tejto metódy je možnosť kombinácie signálov rozptylu svetla s analýzou markerov prítomných na bunkách pomocou fluorescenčne značených protilátok. Umožňuje to identifikáciu subpopulácií buniek, ktoré selektívne podstupujú apoptózu v experimentálnych podmienkach .
Absorpcia farbív
Prietoková cytometria môže byť využitá na kvantitatívnu analýzu počtu živých, apoptotických a nekrotických buniek prostredníctvom absorpcie a udržania konkrétnych farbív. Apoptotické bunky majú počas značnej časti procesu apoptózy intaktnú plazmatickú membránu, to znamená, že aj jej základné funkcie ako aktívny transport a nepriepustnosť pre makromolekulárne látky ostávajú zachované. Rozdiely v permeabilite membrány živých, mŕtvych a apoptotických buniek pre DNA farbivá (napríklad propídium jodid, etídium bromid, farbivo HO33342) môžu rozlíšiť živé, apoptotické a nekrotické bunky. Propídium jodid nie je vylúčený z nekrotických buniek – po vstupe do bunky sa interkaluje do DNA a spôsobuje červenú fluorescenciu nekrotického jadra. Apoptotické bunky majúce neporušenú funkciu plazmatickej membrány ostanú nezafarbené. Živé bunky aktívne pohlcujú farbivo HO33342 a vykazujú modrú fluorescenciu DNA. Aplikácia farbiva HO33342 na apoptotické bunky sa prejaví modrou fluorescenciou, ktorá po čase slabne. Touto metódou sa dajú rozlíšiť živé bunky (modrá fluorescencia), nekrotické bunky (červená fluorescencia) a apoptotické bunky (slabá modrá a žiadna(slabá červená fluorescencia) .
Kaspázy
Kaspázy hrajú dôležitú úlohu v apoptóze. Normálne sa vyskytujú ako proenzýmy, aktivované sú proteolýzou . Identifikácia ich aktívnej formy je založená na naviazaní polyklonálnych protilátok proti ľudskej kaspáze-3 – kľúčovej efektorovej kaspáze v apoptóze buniek .
Mitochondriálne funkcie
Mitochondrie sú považované za základ biochemických procesov apoptózy. Za normálneho stavu je vnútorná strana vnútornej membrány nabitá záporne – to umožňuje kladne nabitým lipofilným fluorochómom akumuláciu v mitochondrii. Kľúčovým bodom v apoptóze je formovanie pórov v ich vonkajšej membráne umožňujúcim únik mitochondriálnych proteínov do cytoplazmy . Otvorenie pórov spôsobí kolaps asymetrického rozloženia protónov na oboch stranách vnútornej membrány a zmenu mitochondriálneho membránového potenciálu . Kladne nabité lipofilné fluorochrómy sa tak nemôžu akumulovať v mitochondrii. Táto zmena potenciálu je merateľná pomocou markerov prietokovou cytometriou .
Únik Ca2+ do cytoplazmy
Transportné systémy pri normálnych podmienkach udržujú intracelulárnu koncentráciu Ca2+ na hladine 100 nM – je to najmenej o štyri rády nižšia koncentrácia ako v extracelulárnom priestore. Počas apoptózy sa koncentrácia cytoplazmatického Ca2+ zvýši. Na demonštráciu toho, že došlo k zvýšeniu intracelulárnej koncentrácie Ca2+ metódou prietokovej cytometrie, sa používajú Ca2+ selektívne fluorescenčné sondy .
Lyzozomálne protónové pumpy
Lyzozómy majú vďaka protónovým pumpám, ktoré sú závislé na ATP, nízke pH . To umožní akumuláciu akridínovej oranže – bázickému fluorescenčnému farbivu – v lyzozóme. Farbivo je v organele kvôli protonácii uväznené. Prítomnosť červenej fluorescencie je známkou aktivity protónových púmp. Schopnosť akumulácie farbiva ostáva zachovaná aj v skorých štádiách apoptózy, zatiaľ čo pri nekróze sa okamžite stráca. Tento rozdiel sa využíva na rozlíšenie medzi apoptotickými a nekrotickými bunkami. Naviazanie farbiva na DNA sa prejaví ako zelená fluorescencia, zatiaľ čo akumulácia v lyzozómoch sa prejaví červenou fluorescenciou. Počas apoptózy sa červená fluorescencia nemení (membrány lyzozómov ostávajú intaktné), zatiaľ čo v nekrotických bunkách sa zníži kvôli porušeniu lyzozomálnej membrány. Zelená fluorescencia ostáva počas skorých štádií apoptózy tiež nemenná .
Literatúra
- https://www.sciencedirect.com/science/article/pii/S0022175900002337#BIB99
- https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2117903/
- https://www.flowjo.com/learn/flowjo-university/flowjo/getting-started-with-flowjo/58
- https://cshperspectives.cshlp.org/content/5/4/a008656.full
- https://www.sciencedirect.com/science/article/pii/S0005272806000727
- https://www.sciencedirect.com/science/article/pii/S0143416011000522?via%3Dihub
- https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4524569/
Zdroj
- KITTNAR, Otomar a kol. Lékařská fyziologie. Praha : Grada, 2011. ISBN 978-80-247-3068-4. (po česky)