Kvasinka
Kvasinky (staršie: kvasinky a kvasinkové mikroorganizmy) je odborné súhrnné označenie pre všetky tie jednobunkové (resp. prevažne jednobunkovo sa vyskytujúce) huby, ktorých nepohlavné rozmnožovanie sa uskutočňuje buď pučaním alebo jednoduchým delením bunky. Inými slovami: Kvasinky sú všetky pravé huby (v tradičnom chápaní, čiže bez bunkoviek), ktoré (za normálnych podmienok) nemajú pravé podhubie (resp. hýfy). Z taxonomického hľadiska vyššie uvedené znamená, že kvasinky tvoria:
- časť taxónu Ascomycota (po slovensky: vreckaté huby); a
- časť taxónu Basidiomycota (po slovensky: bazídiové huby); a
- časť taxónu Deuteromycota (po slovensky: nedokonalé huby).
- Kvasinka je aj slovenský názov rodu Saccharomyces[1], pozri kvasinka (Saccharomyces).
Keďže teda (takto definované) kvasinky patria do viacerých taxónov, nie sú taxonomickým pojmom. Taxón Deuteromycota je neprirodzený taxón, a preto sa v posledných desaťročiach spravidla považuje za nevhodný (resp. zrušený) a jeho obsah (resp. aspoň tá jeho časť, ktorú sa už podarilo prehodnotiť) bol presunutý do taxónov Ascomycota a Basidiomycota.
Kvasinky sú v podstate opak pojmu plesne (t. j. plesne v zmysle mikroskopická vláknitá huba alebo v zmysle saprofytická nevodná mikroskopická vláknitá huba - pozri plesne), pretože mikroskopické pravé huby (v tradičnom chápaní, čiže bez bunkoviek) sa delia na kvasinky, plesne a dermatofyty (pričom dermatofyty sa alternatívne považujú za súčasť plesní).[2][3][4][5][6][7][8][9][10][11][12][13][14][15]
Základná charakteristika
Kvasinky Netvoria plodnice, množia sa hlavne nepohlavne, a to - ako vyplýva z definície kvasiniek - pučaním alebo jednoduchým delením (výnimku tvorí atypické rozmnožovanie rodu Sterigmatomyces). Môžu sa množiť aj sexuálne tvorbou vreciek, ktoré však nie sú uzavreté v žiadnych plodniciach (teda askokarpoch).
Netvoria žiadne pravé podhubie (mycélium); niektoré kvasinky tvoria len pseudomycélium, ktoré sa podobá kolóniám jednobunkových organizmov.
Kvasinky sa hojne využívajú v potravinárstve a biotechnológiách. Používajú sa napríklad pri výrobe vína, piva alebo chleba. Využíva sa ich schopnosť kvasenia. Sú ale medzi nimi aj pôvodcovia chorôb, ako je napr. Candida albicans.
Výskum
Prvé jednotlivé kvasinky prvýkrát pozoroval Anton van Leeuwenhoek, ktorý opísal vo svojich listoch Kráľovskej spoločnosti v Londýne výsledky pozorovaní malých guličiek v pive pomocou primitívneho mikroskopu. Theodor Schwann roku 1837 poprel účasť kyslíku pri kvasnom procese a opísal v kvasiacej tekutine sa rozmnožujúce kvasinky, ktoré označil ako „cukornú hubu“ (odtiaľ rodové označenie Saccharomyces).
Objav moderných sekvenovacých princípov umožnil už v roku 1996[16] sekvenovanie genómu kvasinky ako vôbec prvého eukaryotného genómu.
Význam a využitie

Kvasinky sú mikroorganizmy v mnohých smeroch pre človeka veľmi užitočné. Už pomenovanie „kvasinky“ ukazuje na ich vzťah ku kvasným procesom. Najstarší nález nádobiek na víno pochádza z neolitickej kuchyne v Hajji Firuz Tepe (8500 – 4000 rokov pr. n. l.) na území dnešného Iránu.[17] Kvasenie bolo známe už v starom Babylone v období 6000 – 4000 rokov pred naším letopočtom, kedy sa zo skvaseného odvaru z naklíčeného obilia pripravoval nápoj, ktorý sa dá považovať za predchodcu piva.
V polovici 19. storočia vznikol droždiarenský priemysel[18]. Predtým bolo niekoľko pokusov o priemyselnú výrobu v Holandsku. Najskôr bolo droždie predávané v tekutej forme, neskôr po zdokonalení Tebbenhofom sa prešlo k lisovanému droždiu.
Prvoradý význam majú dnes v kvasnom priemysle pri výrobe liehu, piva, vína, pekárskeho droždia a tiež niektorých mliečnych nápojov. V kŕmnom priemysle naberajú význam kŕmne zmesi z kvasníc. Dosiaľ sú pomerne málo využívané pre potravinárske účely ako zdroj bielkovín a iných biologicky cenných látok. Majú bohatý obsah dobre stráviteľných bielkovín (okolo 40%), cukrov a hlavne komplexu vitamínu B, čo platí zvlášť pre pivovarské kvasinky. Tie sa využívajú aj pri liečbe nervových ochorení, pri zápalových kožných chorobách, pri poruchách tráviaceho traktu a pečeňových chorobách. Geneticky modifikované kvasinky bude možné v blízkej budúcnosti použiť aj na odhaľovanie výbušnín, presnejšie molekúl dinitrotoluénu (DNT).[19]
Vedľa prevažujúceho pozitívneho významu existuje aj negatívny vplyv kvasiniek patogénnych, či nevhodný výskyt kvasiniek čoby škodlivého kontaminantu vo výrobnom procese.
Stavba

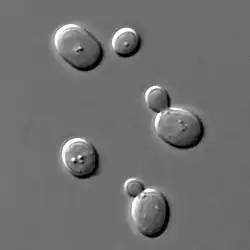
Kvasinky vykazujú veľkú tvarovú, rozmerovú či farebnú diverzitu. Obvykle sú guľaté alebo oválne, ale vyskytujú sa aj druhy, ktoré majú bunky charakteristického citrónovitého, vajíčkovitého, fľaškovitého či vláknitého tvaru. Dokonca medzi samotnými bunkami jedného kmeňa možno nájsť morfologické a farebné odlišnosti. To je spôsobené zmenami fyzikálnych a chemických podmienok v prostredí. Pre zjednodušenie ukážeme štruktúru kvasinky Saccharomyces cerevisiae.
S. cerevisiae sú vo všeobecnosti elipsoidné, v priemere od 5 – 10 μm na veľkej osi, a od 1 – 7 μm na menšej osi. Stredný objem buniek je 29 – 55 μm3 pre haploidné resp. diploidné bunky. Veľkosť bunky sa zväčšuje s vekom.
Makromolekulárne látky v kvasinke:
| typ makromolekuly | kategória | základné komponenty |
|---|---|---|
| proteíny | štruktúrne | aktín, tubulín, históny, ribozomálne proteíny |
| hormóny | alfa a a feromóny | |
| enzýmy | ||
| glykoproteíny | komponenty bunkovej steny | mannoproteíny |
| enzýmy | funkčné enzýmy (invertáza) | |
| polysacharidy | komponenty bunkovej steny | glukán, mannán, chitín |
| komponenty kapsulárne | ||
| zásobné | glykogén, trehalóza | |
| polyfosfáty | zásobné | polyfosfáty vo vakuole |
| lipidy | štruktúrne | voľné steroly v membráne |
| zásobné | triglyceridy, sterolové estery | |
| funkčné | deriváty fosfoglyceridov, voľné mastné kyseliny | |
| nukleové kyseliny | DNA | génová DNA (80%), mitochondriálna |
| RNA | rRNA (80%)m mRNA (5% cytosól, ER, mitochondrie), tRNA |
Bunková stena

V roku 1970 Kidby a Davis navrhli model bunkovej steny pre druh S. cerevisae, ktorý stále ešte rámcovo platí. Podľa tohto modelu má bunková stena tri vrstvy: vonkajšiu, strednú a vnútornú, pričom polysacharid-proteínové komplexy sú podľa tohto modelu spojené fosfodiesterovými väzbami:
- vonkajšia stena je orientovaná smerom do prostredia, obsahuje mannanproteíny spojené disulfidovými mostíkmi.
- stredná stena je tvorená β-1,6 glukánom, gluganproteínmi a mannanproteínmi
- vnútorná stena, priliehajúca na cytoplazmatickú membránu, je zložená z mikrokryštalického β-1,3-glukánu
V bunkovej stene sa nachádzajú taktiež hydrolytické enzýmy glykoproteínového typu, napr. β-fruktofuranosidasa (invertáza).
Polysacharidy bunkovej steny môžu byť jednozložkové (homopolysacharidy) alebo viaczložkové (heteropolysacharidy) – napr. galaktomannány, xylomannany, arabomannany aj. Polysacharidy určujú predovšetkým imunologické vlastnosti kvasiniek, pretože pôsobia ako antigény. Ide predovšetkým o mannanproteíny, ktorých antigénny charakter je určený dĺžkou bočných reťazcov mannosylových zvyškov, spojených α -1,2 a α -1,3 väzbami. Polysacharid – proteínové komponenty bunkovej steny sa taktiež podieľajú na flokulačných schopnostiach kvasiniek.
Jazvy
Ako stále štruktúry na bunkovej stene vznikajú jazvy po pučaní dcérskych buniek a ich separácii od materskej bunky, popr. po delení kvasinkových buniek. Pretrvávajú počas celého vývoja jednotlivej bunky a ovplyvňujú tzv. architektúru jej steny. Vznik jaziev a ich stavbu možno sledovať napr. v optickom mikroskope s využitím fluorescencie primulínu, čo je farbivo, ktoré sa špecificky viaže na mikrofibrily.
Jazva na dcérskej bunke sa volá jazva zrodu, a pretože je je tvorená iným bunkovým materiálom, možno ju odlíšiť od jaziev na miestach, kde sa oddelili nové výhonky. Z každej bunky môže vypučať len obmedzený počet nových buniek.
Cytoplazmatická membrána
Cytoplazmatická membrána (alebo plazmalema) tvorí elastický obal protoplastu, osmotickú bariéru a kontroluje transport látok. Je tvorená predovšetkým z fosfolipidov a bielkovín.
Štruktúra cytoplazmatickej membrány je zrejme najlepšie popísaná v rámci mozaikového modelu bunkových membrán, ktorý hovorí, že molekuly fosfolipidov sú usporiadané v dvoch vrstvách tak, že polárne časti sú na vonkajších stranách a nepolárne vo vnútri membrány.
Pre plazmalemu kvasiniek sú charakteristické invaginácie do cytoplazmy, ktoré sú početné taktiež v exponenciálnej fáze rastu.
Jadro
Jadro býva u eukaryotických organizmov zreteľne ohraničené jadrovou membránou a tvorí ho chromatín, teda komplex dsDNA, histónov a proteínov nehistónovej povahy. Na rozdiel od väčšiny eukaryoriotických buniek majú kvasinky tzv. endomitózu, takže delenie jadra prebieha bez rozrušenia jadrovej membrány. To je hlavný dôvod, prečo je u kvasiniek mimoriadne ťažké urobiť dobrý karyotyp a zistiť tak presný počet a stavbu chromozómov. Na jadrovej membráne sa diferencuje tzv. polárne teliesko, ktoré hrá významnú úlohu pri iniciácii tvorby výhonku.
Sekrečná dráha u kvasiniek
Biomembrány tvoria povrchové štruktúry organel aj systém vnútorných membrán. Tým bunku rozdeľujú na rôzne reakčné priestory. V niektorých z nich dochádza k sekrécii proteínov. V endoplazmatickom retikule dochádza k syntéze peptidov a bielkovín. Na vonkajšom povrchu ER (drsné ER) sú zakotvené ribozómy. Ďalšie ribozómy sa pohybujú voľne v cytoplazme. Rovnako ako u ostatných eukaryotov sa ribozóm skladá z malej (40S) a veľkej (60S) podjednotky. Veľká podjednotka katalyzuje syntézu bielkovín. Na tvorbe ribozómov sa podieľa jadierko. Vnútorný povrch ER tvorí hladké ER. V ER dochádza aj k posttranslačným úpravám. Na miesto určenia sa proteíny dostávajú prostredníctvom sekréčnych mechúrikov, ktoré tvoria Golgiho aparát.
Endoplazmatické retikulum
Endoplazmatické retikulum vytvára u kvasiniek lamely, cisterny a tubuly podobne ako u rastlinných a živočíšnych buniek. Jeho obsah (enchylema) je uzavrený dvoma membránami. ER nadväzuje na vnútorný povrch cytoplazmatickej membrány a niekedy je spojené s jadrovou membránou.
Ribozómy
Ribozómy kvasiniek majú obdobne ako u iných eukaryotov hodnotu sedimentačného koeficientu okolo 80S. Menšia podjednotka s koeficientom 40S je zložená z jedinej molekuly 18S rRNA a z 30 ± 5 bielkovín; väčšia podjednotka 60S obsahuje 28S rRNA, 5S rRNA, 5,8S rRNA a 40 ± 5 bielkovín.
Mitochondrie
Mitochondrie sú organely špecializované na respiráciu a oxidatívnu fosforyláciu. Majú vlastné gény a proteosyntézu. Tvar, štruktúra a počet mitochondrií môžu byť ovplyvňované rôznymi faktormi. Napr. v podmienkach reprimujúceho dýchania sa výrazne znižuje podiel, ktorý mitochondrie zaberajú v celkovom objeme bunky.
Vakuola
Kvasinková bunka spravidla obsahuje jednu veľkú guľatú vakuolu. V počiatočnej fáze pučania možno pozorovať väčšie množstvo malých vakuol, ktoré však počas rastu splývajú do jednej vakuoly, prípadne dvoch vakuol (v stacionárnej fáze rastu). Priemer vakuol je premenlivý od 0,3 do 3 μm. Ohraničené sú membránou zvanou tonoplast.
80 až 90 % rozpustných aminokyselín z celkového objemu voľných aminokyselín v kvasinkách je obsiahnutých vo vakuolách. Ďalej sa tu nachádzajú enzýmy hydrolázy, väčšina proteáz, ribonukleázy či esterázy. Týmito enzýmami sa vo vakuolách rozkladajú funkčne narušené organely.
Referencie
Tento článok je čiastočný alebo úplný preklad článku Kvasinky na českej Wikipédii (číslo revízie nebolo určené).
- Zoznam vyšších a nižších rastlín Slovenska 1998
- kvasinky. In: Malá slovenská encyklopédia. 1. vyd. Bratislava : Encyklopedický ústav SAV; Goldpress Publishers, 1993. 822 s. ISBN 80-85584-12-3. S. 377.
- Hefen. In: Lexikon der Biologie. [CD-ROM] München : Elsevier, Spektrum, Akad. Verl., 2005. ISBN 3-8274-0342-1.
- The Yeasts : a taxonomic study. Ed. N.J.W. Kreger-van Rij. 3rd rev. and enl. ed. Amsterdam : Elsevier Science Publishers, 1984. 1082 s. Dostupné online. ISBN 0-444-80421-8. S. 1 – 3.
- Michael R. McGinnis and Stephen K. Tyring. Introduction to Mycology. In: BARON, S. ed. Medical Microbiology. 4th edition. 1996 ,
- The Yeasts : a taxonomic study. Ed. Cletus P. Kurtzman ; Jack W. Fell. 4., rev. and enl. ed. Amsterdam : Elsevier, 1998. 1055 s. Dostupné online. ISBN 0-444-81312-8. S. 3.
- Miniatlas mikroorganismů [online]. is.muni.cz, [cit. 2020-08-12]. Dostupné online.
- yeast. In: McGraw-Hill Concise Encyclopedia of Bioscience
- Roche Lexikon Medizin (5. Aufl.) [online]. elsevier-data.de, [cit. 2020-08-12]. Dostupné online.
- MÜCKE, Wolfgang; LEMMEN, Christa. Schimmelpilze : Vorkommen, Gesundheitsgefahren, Schutzmaßnahmen. 3. überarb. und erw. Aufl. Landsberg : ecomed, 2004. 182 s. ISBN 978-3-609-68001-9. S. 17.
- SCHWANTES, Hans Otto. Biologie der Pilze : eine Einführung in die angewandte Mykologie. Stuttgart : Ulmer, 1996. 478 s. ISBN 978-3-8252-1871-3. S. 230 – 231, 240, 268.
- ADL, Sina M., at al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. Journal of Eukaryotic Microbiology, 2019, roč. 66, čís. 1, s. 4 – 119. ISSN 1550-7408. DOI: 10.1111/jeu.12691.
- SEDLÁŘ, Jiří. Odd. EUMYCOTA – pravé houby [online]. old.botany.upol.cz, 2007, [cit. 2020-08-12]. Dostupné online.
- Obecná mykologie : (místy se zvláštním zřetelem k makromycetům) [online]. sci.muni.cz, rev. 2015-05-13, [cit. 2020-08-12]. Dostupné online.
- IVARSSON, Magnus; DRAKE, Henrik; BENGTSON, Stefan; RASMUSSEN, Birger. A Cryptic Alternative for the Evolution of Hyphae. BioEssays, 2020, roč. 42, čís. 6. ISSN 1521-1878. DOI: 10.1002/bies.201900183.
- Čo sa o sebe dozvedáme z našej genetickej informácie
- Kvasinky človeku vernejšie než pes
- Kvasná chémia alebo vznik piva, vína u nás z historického hľadiska
- Populárny článok (na Idnes) o štúdii odborníkov Philadelphijskej univerzity, ktorá bola zverejnená v časopise Nature Chemical Biology.