Ediakarská fauna
Ediakarská fauna je označení pro tajemné přisedlé organismy, které na Zemi žily v období ediakara, jež probíhalo před asi 635 až 542 miliony lety. Fosilie těchto organismů pocházejí z celého světa; zástupci fauny představují nejstarší známé mnohobuněčné organismy. Před 575 miliony lety, kdy se Země vymanila z globálního zalednění, možná ediakarská fauna prodělala evoluční radiaci během takzvané avalonské exploze. Zanikla při rychlém rozvoji živých organismů známém jako kambrická exploze. Zřejmě poslední zástupci ediakarské fauny jsou vzácně objevováni ve vrstvách ze středního kambria starých 510 až 500 milionů let, avšak dřívější komunity zanikly na konci ediakary a zanechaly po sobě pouze otisky kdysi prosperujícího ekosystému.


Většina současných tělních plánů živočišných těl se poprvé objevila až u kambrijských živočichů a ne u ediakarské fauny. Zdá se, že kambrijské organismy plně nahradily organismy dominující během ediakary, stále však o tom probíhají debaty. Existuje několik hypotéz, proč ediakarská fauna z fosilních záznamů vymizela, včetně měnícího se prostředí, nástupů predátorů či konkurence jiných forem života. Jedna hypotéza tvrdí i to, že ediakarská fauna mohla dále prosperovat, pouze již okolní podmínky zabraňovaly její fosilizaci.
Zařazení ediakarské fauny do fylogenetického stromu se ukázalo býti náročné. Není dokonce ani prokázáno, že se jednalo o živočichy. Mohlo jít také o lišejníky, řasy, protisty známé jako Foraminifera, houby, mikrobiální kolonie či hypotetický mezistupeň mezi rostlinami a živočichy. Stavba těla a návyky některých druhů (například Funisia dorothea) vykazuje podobnost s houbovci (Porifera) nebo žahavci (Cnidaria), rod Kimberella se podobá měkkýšům (Mollusca). U některých ediakarských tvorů se mohla vyvinout bilaterální symetrie (dvoustranná souměrnost), i když to je sporné. Většina makroskopických ediakarských fosilií je stavbou těla odlišná od pozdějších živočišných forem: připomínají disky, trubky či baňky. Vzhledem k obtížnosti stanovení evolučních vztahů mezi těmito tvory a současnými organismy někteří paleontologové usoudili, že představují zvláštní vyhynulou linii, která není blízce příbuzná žádným jiným tvorům a navrhli je zařadit do samostatné říše Vendozoa (nyní přejmenované na Vendobionta). Pokud ediakarská fauna nazanechala žádné potomky, její vývoj by mohl být považován za „neúspěšný experiment“ v evoluci mnohobuněčného života.
Objev

Poprvé objevil nějakého zástupce ediakarské fauny, a sice druh Aspidella terranovica, roku 1868 skotský geolog Alexander Murray na Newfoundlandu. Protože se však nález nacházel pod vrstvou nazvanou „Primordial Strata“, která dle tehdejších názorů obsahovala první známky života, názor kanadského paleontologa Elkanaha Billingse z roku 1872, že se jedná o živočichy, byl odmítnut a útvar tehdejší vědci považovali za bubliny po úniku plynu či konkreci. Jinde na světě podobné útvary nebyly pozorovány, a proto se o tento nález již nikdo nezajímal.[1] Až roku 1933 objevil Georg Gürich objevil podobný otisk v Namibii,[2] ale v souladu s všeobecným přesvědčením o vzniku mnohobuněčných organismů v kambriu jej datoval do tohoto období. Další nález učinil Reginald Sprigg, kterého v roce 1946 jihoaustralská vláda vyslala, aby prozkoumal ziskovost opuštěných dolů v oblasti Ediacara Hills ve Flindersově pohoří. Sprigg zde nalezl fosilie velmi starých organismů, které datoval do raného kambria až prekambria.[3] Jeho práce ale nevzbudila velkou pozornost vědců a článek pro časopis Nature byl odmítnut. Sprigg představil své nálezy na Mezinárodním geologickému kongresu v Londýně v roce 1948, ale jeho přednášky se setkaly s nedůvěrou nebo nezájmem.[4]
Prekambrium nebylo za období, v němž existoval život, považováno až do roku 1957. Tehdy byla v Charnwoodském lese objevena slavná Charnia[5] a díky pečlivému geologickému mapování od British Geological Survey bylo jisté, že se nález nacházel v prekambrijské vrstvě. Na spojitost mezi tímto a staršími nálezy přišel australský geolog a paleontolog Martin Glaessner roku 1959. V souvislosti se zlepšeným datováním a rozpoutáním zájmu okolo prekambrijských tvorů pak bylo nalezeno mnoho dalších zástupců fauny.[6][7][8]
Všechny exempláře objevené před rokem 1967 pocházely z hrubozrnného pískovce, kvůli čemuž nebyly zachovány jemné detaily, což ztěžovalo výzkum. Shiva Balák Misra však tehdy objevil v popelu v Newfoundlandu dobře zachovalé fosilie s viditelnými detaily, což umožnilo podrobnější studium fosilií.[9]
Špatná komunikace mezi paleontology společně s odlišnými formacemi způsobila, že faunu nazývali různě. Roku 1960 bylo ke dvěma hlavním názvům: sinianská a vendianská, přidáno i francouzské slovo ediacarien po Ediacaran Hills v jižní Austrálii, kde fosilie nalezl Sprigg. Toto jméno pochází z domorodého jazyka, znamená přítomnost vody a přeneseno bylo na faunu. Název byl následně použit pro označení geologického období.[10][11] Mezinárodní komise pro stratigrafii (ICS) uznala období v roce 2004. Šlo o první nově definované geologické období po více než sto letech.
Zachování
Fosilizace

Fosilizace ediakarské fauny je jedním z velkých divů vědy. Jako měkké organismy by normálně fosilie tvořit neměly a na rozdíl od jiných měkkých tvorů zachovaných například v Burgesských břidlicích neměly na jejich fosilizaci vliv neobvyklé místní podmínky, jednalo se o globální jev. Procesy, které při jejich fosilizaci probíhaly, musely být systémové a celosvětové. Během ediakary musely zřejmě panovat odlišné podmínky, které dovolily fosilizaci těchto tvorů. Pravděpodobně byli zachováni kvůli rychlému pokrytí popelem nebo pískem, které organismy pohřbily v bahně či mikrobiálních povlacích, kde ediakarská fauna žila.[12] Jejich zachování snad napomohla vyšší koncentrace křemíku v oceánech, možná před širokým rozšířením organismů jako jsou mořské houby či rozsivky, které křemík pohlcují (a používají k výstavbě schránek).[13] Fosilie s více zachovanými detaily jsou nacházeny ve vrstvách popelů, které mohou být snadno a přesně datovány do příslušného geologického období.[14]
Mikrobiální povlaky

Mikrobiální povlak je označení pro sedimenty stabilizované pomocí kolonií mikrobů, kteří pomocí vylučovaných lepkavých kapalin či jiných způsobů vážou částice sedimentů k sobě. Je-li uložena příliš hustá vrstva sedimentu, která těmto mikroorganismům brání v růstu či rozmnožování, uhynou a zanechávají fosilie s charakteristickým vrásčitým povrchem (přirovnávaným ke sloní kůži) a bradavičnatou strukturou.[15]
Některé vrstvy z ediakary s touto charakteristickou texturou obsahují fosilie a fosilie ediakarské fauny jsou téměř vždy nacházeny v usazeninách společně s mikrobiálními povlaky. Přestože kdysi byly mikrobiální povlaky rozšířené, po rozšíření kambrijských organismů, kteří se těmito mikroorganismy živily, poklesl jejich počet. Nyní jsou mikrobiální povlaky rozšířeny v nehostinných refugiích, jako jsou stromatolity v rezervaci Hamelin Pool Marine Nature Reserve v Žraločí zátoce při pobřeží západní Austrálie, kde salinita vody může až dvakrát přesahovat obsah soli v okolním moři.[16]
Míra zachování
Rychlost usazení sedimentů společně s rychlostí rozkladu měkkých organismů určuje, jestli se zachová jejich horní nebo spodní část. Většina kruhovitých organismů se začala rozkládat ještě před usazením sedimentů a následně byly zasypány popelem či pískem. Tak byla zachována spodní strana daného tvora. Fosilie matracovitých organismů mají tendenci se rozložit i po usazení a stmelení nadložního sedimentu, díky čemuž jsou zachovány otisky jejich svrchních částí. Odolnost jejich těl se odráží v tom, že ve vzácných případech jsou jejich fosilie nacházeny v sedimentech uložených během splachu materiálu během silných bouří, kdy může docházet k rapidní sedimentaci. To, že byly schopné tuto rapidní sedimentaci přežít, napovídá, že musely být odolné. V některých případech srážení minerálů působením bakterií vytvořilo „posmrtnou masku“ vedoucí ke vzniku pozitivního fosilního otisku tehdejšího organismu.[17][18]
Popis
Ediakarská fauna vykazovala širokou škálu morfologických charakteristik. Velikost těchto organismů sahala od milimetrů do metrů, jejich těla měla tvar od jednoduchých kapkovitých těl až po těla složité stavby a mohla být jak tvrdá a odolná, tak měkká. Vyvinuly se téměř všechny formy symetrie.[12] Rozlišeno bylo několik typů morfologií:





- Embrya
- Ve zprávách o objevech předkambrijského mnohobuněčného života dominovaly informace o fosiliích podobných embryím, nalezené převážně v čínském souvrství Tou-šan-tchuo (v jihozápadní Číně, v provincii Kuej-čou). Některé objevy[19] vyvolávají velký mediální rozruch,[20] ačkoli o jejich původu existují různé názory a dle některých se může jednat i o anorganické struktury vytvořené vysrážením minerálů.[21] Jiná „embrya“ byla interpretována jako pozůstatky obrovských síru redukujících bakterií (jako je Thiomargarita namibiensis),[22] tento názor měl od roku 2007 velké množství podporovatelů,[23][24] po dalším výzkumu, který srovnával vzhled daných fosilií s bakterií Thiomargarita, však tato teorie začala upadat.[25]
- Mikrofosilie, které pocházejí z období před 632,5 miliony lety – pouhé tři miliony let po konci velkého zalednění, mohou představovat „klidové stadium“ ve vývoji nejstarších známých živočichů. Podle alternativního návrhu pak může jít i o dospělé mnohobuněčné organismy.[26][27]
- Disky
- Diskovité fosilie, jako jsou Ediacaria, Cyclomedusa a Rugoconites, byly zpočátku identifikovány jako zástupci žahavců (Cnidaria), mezi které patří medúzy a koráli.[3] Další výzkumy těchto fosilií navrhly jiné interpretace. Žádná z těchto fosilií již není považována za medúzu; může se jednat například i o zástupce protistů (Protista).[28] Mnoho jednotlivých fosilií bylo určeno jako mikrobiální kolonie,[29][30] jiné mohou být škrábanci vzniklými otáčením organismů kolem jejich „stonku“, který je poutal k mořskému dnu.[31] Některé znaky, pomocí kterých by šlo druh lépe identifikovat, často chybí, protože se obyčejně zachovala pouze spodní část organismu.
- Taškovité fosilie
- Fosilie, jako je Pterinidium, zachované ve vrstvách sedimentů, připomínají „bahnem naplněné tašky“. Na interpretaci těchto organismů není shoda.[32]
- Toroidní fosilie
- Fosilie živočicha popsaného jako Vendoglossa tuberculata, pocházející z Namibie, byla identifikována jako dorzoventrálně stlačený torus. Fosilie má tvar jazyka a zužuje se k tupě špičatému konci.[33]
- Matracovité organismy
- Ediakarské organismy řazené do Seilacherovy revidované skupiny Vendobionta[34] mají vzhled podobný nafukovací matraci. Někdy se těla těchto tvorů ještě před fosilizací roztrhala či popraskala, pomocí těchto poškozených vzorků lze však organismy snadněji rekonstruovat. Například uspořádání „listů“ tvora Swartpuntia germsi (tři a více listů dokola podél stonku) mohlo být rozpoznáno pouze z podobných poškozených fosilií, neboť fosilie jsou obvykle bahnem a pískem stlačeny do roviny.[35]
- Tyto organismy byly rozděleny do dvou kladů: Rangeomorpha a Erniettomorpha.[36] Zahrnují taxony jako Charnia a Swartpuntia, které jsou nejznámější z celé ediakarské fauny, zároveň je však lze těžko umístit do současného fylogenetického stromu. Neexistuje žádný důkaz ohledně jejich vnitřní anatomie. Nejčastější, avšak někdy zamítanou teorií, je, že tito tvorové získávali živiny z okolní mořské vody pomocí osmotrofie nebo osmózy.[37][38]
- Další fauna
- U některých organismů z ediakary se zachovaly složitější detaily, což je umožnilo identifikovat jako možné předky některého žijícího kmenu, čímž však zároveň neodpovídají některým definicím ediakarské fauny.
- Jednou z prvních z těchto fosilií je Vernanimalcula o možné bilaterální symetrii.[21][39] Později nalezené fosilie jsou téměř všeobecně považovány za dvojstranně souměrné. Jedná se například o rod Kimberella podobající se měkkýšům,[40] Spriggina[41] a štítovité organismy Parvancorina.[42] O příbuznosti těchto tvorů probíhají diskuse.[43] Mezi další organismy té doby patří fosilie pojmenované anglickým termínem Small shelly fossils (nejslavnějším zástupcem této skupiny je Cloudina)[44]. Tyto organismy, chráněné tvrdou skořápkou, naznačují, že přestože nebyla predace v ediakaře běžná, byla minimálně přítomná. Během ediakary žili i někteří zástupci moderních taxonů, jako houbovci, červené a zelené řasy, protisté a bakterie. Byli popsáni také možní členovci.[45]
- Fosilie stop
- Z období ediakara pocházejí pouze nálezy horizontálních cest či chodeb[46] na nebo těsně pod povrchem mořského dna, pouze velmi málo jich je vertikálních. Podobné nálezy mohly zanechat pohyblivé organismy, pravděpodobně o bilaterální symetrii,[47] je však také možné, že je vytvořily jednodušší organismy, které se pomalu válením přesunovaly po mořském dně, přičemž sbíraly potravu.[48]
- Některé fosilie, převážně diskovité, byly zkresleně interpretovány jako fosilie stop, ale tato hypotéza nedosáhla širokého přijetí. Stejně jako nalezené cesty, i jiné fosilie stop byly přímo spojeny s ediakarskou faunou. Fosilie rodů Yorgia a Dickinsonia jsou často nalézány na koncích dlouhých cest, které odpovídají tvaru těchto živočichů; předpokládá se, že tyto organismy sbíraly potravu pomocí řasinek a tak tyto stopy vytvořily,[49] avšak přesný způsob vytvoření fosilií zůstává z velké části záhadou.[50] V roce 2019 přišli čínští vědci s objevem nového červovitého druhu dvojstranné souměrnosti, který byl pojmenován Yilingia spiciformis a žil na konci období ediakary. Pozůstatky tohoto druhu byly objeveny na konci fosilizovaných stop, které vytvořil.[51] Potenciálnímu měkkýši rodu Kimberella jsou zase přisuzovány škrábance, které mohly být vytvořeny pomocí raduly, orgánu měkkýšů sloužícímu ke konzumaci potravy.[52]
Klasifikace

Systematika ediakarské fauny je obtížná, a proto existují nejrůznější teorie o způsobech, jak je zařadit do fylogenetického stromu.
V roce 1998 americký paleontolog a profesor geologie Mark McMenamin navrhl předpoklad, že se u ediakarské fauny nevyvinulo embryonální stádium, nešlo tedy o živočichy, ale zároveň šlo o organismy s nervovým systémem a mozkem, což znamená, že „cesta k inteligentnímu životu na této planetě byla zahájena více než jednou“.[28]
Žahavci
Vzhledem k tomu, že nejprimitivnější zástupci podříše Eumetazoa jsou žahavci (Cnidaria), byli zprvu zástupci ediakarské fauny řazeni mezi medúzy či pérovníky.[53] Příbuznost mezi některými zástupci ediakarské fauny a pérovníky byla nicméně zpochybněna několika důkazy.[54] Další objevy prokázaly, že mnoho kruhovitých pozůstatků, považovaných za pozůstatek medúz, jsou pískem plněné vaky, které přidržovaly některé čeledi ediakarské fauny u mořského dna. Pozoruhodným příkladem této formy je tvor s názvem Charniodiscus, kruhová fosilie, ke které byl později nalezen dlouhý „stonek“.[55][56]
Protisté

Německý paleontolog Adolf Seilacher navrhl, že ediakarská fauna představovala obří protisty.[57] Například moderní zástupci třídy Xenophyophorea jsou obřími prvoky, kteří žijí v oceánech celého světa, především u abyssálních plošin. Genetická studie naznačila, že se řadí do kmenu dírkonošců (Foraminifera). Existuje přibližně 42 uznaných druhů ve 13 rodech a 2 řádech. Jeden z nich, Syringammina fragilissima, patří se svou velikostí až 20 cm mezi největší známé prvoky.
Nový vyšší taxon
Seilacher také navrhl, že by ediakarská fauna mohla představovat jedinečné a vyhynulé seskupení příbuzných forem vycházejících z jednoho společného předka, pro které vytvořil novou říši Vendozoa,[58][59] pojmenovanou podle nyní zastaralého názvu fauny vendianská. Později z této systematiky vyřadil organismy identifikované jako živočichy a pro zbylé vytvořil kmen Vendobionta.
Tvory v něm popsal jako žahavce prošívaných tvarů, kteří však nemají knidocyty. Jejich nepřítomnost jim bránila současnému způsobu získávání potravy, proto Seilacher usoudil, že zástupci ediakarské fauny uzavírali symbiotické vztahy mezi fotosyntetizujícími a chemotrofními organismy.[60] Americký paleontolog Mark McMenamin považoval tento způsob získávání potravy za typický pro celou ediakarskou faunu a tyto mořské organismy označil za „ediakarskou zahradu“.[61]
Lišejníky

Australský paleontolog a geolog Greg Retallack navrhl zástupce ediakarské fauny zařadit mezi lišejníky.[62][63] Tato hypotéza však nezískala širší podporu.[64][65] Podle argumentů Retallacka nejsou fosilie ediakarské fauny tak rozmačkané jako fosilie známých medúz a spíše připomínají stlačené dřeviny. Retallack poukazuje na to, že pokud by byly některé lišejníkové kolonie chráněny chitinem, poskytlo by jim to větší odolnost proti stlačení. Také tvrdí, že velká velikost organismů (až 1,5 m) odporuje zařazení mezi živočichy, stejně tak poukazuje na velkou rozmanitost ediakarských fosilií, jež je srovnatelná s rozmanitostí forem hub, řas a lišejníků.[62] Odpůrci naopak většinu důkazů hodnotí jako chybné, nejednoznačné nebo nedostatečné.[64] Retallack rovněž představil hypotézu, podle níž někteří zástupci ediakarské bioty nežili v moři, ale byly to přisedlé suchozemské organismy chladných a suchých půd, což má opět potvrzovat, že fosilie byly vzhledem i ochranou podobné spíše lišejníkům.[66][67]
Jiné interpretace
Ediakarská fauna byla řazena i do jiných skupin,[68] od řas,[69] prvoků,[70] hub,[71] bakteriálních či mikrobiálních kolonií[29] až k hypotetickému mezistupni mezi rostlinami a živočichy.[72]
Roku 2014 byl objeven nový žijící rod Dendrogramma. Tým objevitelů ukázal na podobnost s některými medúzoidními zástupci známými z fosilního záznamu ediakarské fauny, jako jsou Albumares brunsae, Anfesta stankovskii nebo Rugoconites enigmaticus,[73][74] a jednou z fylogenetických hypotéz proto bylo, že se jedná o přežívající zástupce některé z bazálních větví živočichů. Pozdější molekulárně genetická analýza ukázala, že se nejedná o neznámou vývojovou linii, ale prokázala příslušnost k řádu trubýšů, tedy žahavců třídy polypovci, a byla diskutována možnost, že i někteří zástupci ediakarské fauny patřili mezi podobné živočichy. V dendrogrammách však byly nakonec rozpoznány pouhé odlomené zploštělé konce obranných chapadel trubýšů, tedy nikoli dospělí jedinci ani jejich larvární stadia. Jako indicie pro zařazení některých ediakarských rodů do žahavců jsou proto dendrogrammy nepoužitelné.[75][76]
Vývoj
První zástupci ediakarské fauny se objevili před 655 miliony lety, asi 4 miliardy let po vzniku Země. Přestože údajné pozůstatky života jsou hlášeny do doby před 3 460 miliony lety,[77][78] nepochybné důkazy pocházejí až z dob před 2 700 miliony lety,[79] přičemž eukaryota existovala před 1 200 miliony lety.[80] Existuje několik hypotéz, proč vývoj složitějších organismů, jako ediakarské fauny, trval tak dlouho.
Žádné zvláštní vysvětlení nemusí být zapotřebí. Pomalý proces evoluce jednoduše potřeboval 4 miliardy let k vývoji nezbytných adaptací. Zdá se, že se úroveň složitosti organismů pomalu zvyšovala a v průběhu času se vyvíjelo stále více složitějších forem života, jak ukazují stopy dřívějšího složitějšího života, jako Nimbia nalezená v 610 milionů let starém souvrství Twityy[81] (a možná i ve starším souvrství z dob před 770 miliony lety[82]), které pravděpodobně představovaly nejsložitější organismy doby. Globální zalednění mohla vývoj mnohobuněčných organismů brzdit nebo dočasně zastavit.
Další možností je, že zvětšování těl nebylo do vzniku ediakarské fauny výhodné, protože prostředí upřednostňovalo malé tvory před velkými. V současné době lze mezi podobné organismy zařadit plankton, jehož malá velikost mu umožňuje rychle se rozmnožovat a využívat na živiny bohatý vodní květ. Ovšem pokud by tehdejší doba vůbec nepodporovala velké organismy, muselo by být tehdejší prostředí velmi odlišné.
Primárním limitujícím faktorem bylo množství atmosférického kyslíku. Bez komplexního oběhového systému nebyly schopny organismy jeho malé množství čerpat dostatečně rychle, aby pokryl jejich metabolickou potřebu.
Na rané Zemi existovaly reaktivní prvky, jako železo a uran, které reagovaly s atmosférickým kyslíkem produkovaným fotosyntetizujícími organismy. Kyslík se nebyl schopen udržet v atmosféře, dokud železo a všechny ostatní reaktivní prvky nezoxidovaly. Geolog Donald Canfield zaznamenal, že v atmosféře poprvé výrazně narostlo množství kyslíku těsně předtím, než se objevila ediakarská fauna[83] − přítomnost atmosférického kyslíku byla možným spouštěčem evoluční radiace ediakarských organismů.[84] Zvyšování podílu kyslíku v atmosféře přišlo ve dvou etapách. Při první se vyvinuly první, přisedlé, organismy, pri druhé větší a pohyblivější tvorové.[85] Předpoklady vlivu atmosférického kyslíku však sklízejí určitou kritiku opírající se o to, že jeho snížené množství během kambria či křídy nemělo vliv na tehdejší život.[86]
Jako další překážka ve vývoji mnohobuněčných organismů byla navržena období glaciálů. Nejstarší známá „embrya“ z Číny se objevují pouze milion let poté, co se Země vymanila z možného celkového zalednění, což by naznačovalo, že ledová pokrývka a studené oceány mohly zabránit vzniku mnohobuněčného života.[87] Teoreticky se mohl komplexní život vyvinout ještě před tímto zaledněním a později být zničen. Teorie však sklidila i kritiku; například studené podmínky dnešní Antarktidy nikterak neovlivňují rychlost vývoje tamějších organismů.
Na začátku roku 2008 zkoumala skupina vědců fosilie ediakarských organismů ze tří různých vrstev. Zkoumány byly fosilie z Avalonského poloostrova (Kanada) staré 575 až 565 milionů let, dále z Bílého moře na území Ruska z doby před 560 až 550 miliony lety a z Nama v Namibii (fosilie staré 550 až 542 milionů let), z období bezprostředně před začátkem kambria. Mezi těmito třemi skupinami nebyl objeven žádný významný rozdíl, z čehož vyplývá, že musely projít evoluční „explozí“, analogické slavné kambrické explozi.[88]
Zachování fauny

Nedostatek fosilií ediakarské fauny po kambriu může být jednoduše způsoben změněnými podmínkami, které již neumožňovaly fosilizaci těchto organismů, které mohly nadále prosperovat, pouze se nedochovaly jejich fosilie.[15] Pokud by však v této době byly běžné, lze přesto nějaké exempláře očekávat,[89] například ve výjimečně dobře zachovalých vrstvách, jako jsou Burgesské břidlice v Kanadě či naleziště z čínského Čcheng-ťiangu v maotchienšanských břidlicích.[90] Obecně se neuznává, že by ediakarská fauna přežívala ještě v kambrijském období, i když existuje několik sporných zpráv, například nepublikované pozorování „vendobionty“ z doby před 535 miliony lety v Číně.[91]
Predace a pastva
Časné kambrijské organismy, stojící v potravinovém řetězci výše, mohly likvidovat mikrobiální povlaky. Když se tito tvorové s pasoucím se stylem života prvně objevili, počet ediakarské fauny začal klesat, což by mohlo naznačovat, že došlo k destabilizaci mikrobiálního substrátu. Mohlo dojít až k narušení ekosystému a vymírání.
Alternativně by mohli obrnění tvorové lovit přímo ediakarské organismy, které nebyly před útoky predátorů dostatečně chráněné. Nicméně, pokud je interpretace rodu Kimberella jako pasoucího se tvora správná, byly ediakarské organismy již v předkambrijském období vystaveny predaci, byť omezené.[28][40]
Kompetice

Je možné, že zvýšená konkurence v důsledku vývoje nových inovací u ostatních živočišných skupin (možná jako reakce na predaci)[92] vedla až k vytlačení ediakarské fauny. Tento argument však nevysvětlil podobné jevy. Například převaha mlžů nad ramenonožci je nakonec považována za náhodný výsledek dvou nesouvisejících trendů.[93]
Změna přirozeného prostředí
Je těžké odvodit vliv měnících měnících se planetárních podmínek na organismy, komunity a ekosystému, avšak na konci prekambria a na začátku kambria došlo k velkým změnám. Rozpad superkontinentu Rodinie,[94] stoupající hladiny oceánů (vytváření mělkých moří),[95] nedostatek živin,[96] kolísání složení vzduchu včetně kyslíku a oxidu uhličitého[97] či změny chemického složení oceánů[98] mohly hrát roli při vymizení ediakarské fauny.
Nálezy
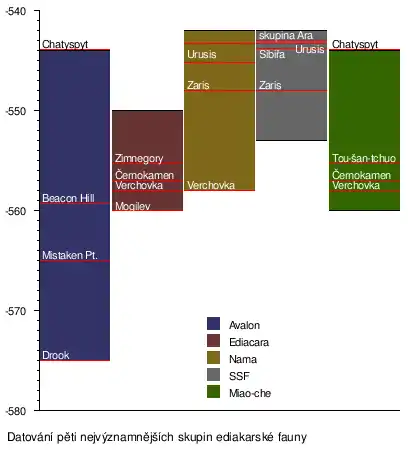
Ediakarské fosilie byly nalezeny v 25 oblastech[11] po celém světě v různých usazeninách a jsou uskupené do tří hlavních skupin pojmenované podle typických lokalit. Každá z těchto skupin se po počátečním výbuchu diverzifikace po zbytek své existence změnila jenom málo.[99]
Skupina avalonského typu
Skupina avalonského typu je definována podle nálezů v Mistaken Point na území Kanady, kde se nachází velké množství pozůstatků této fauny.[100] Datování zdejších organismů je jednoduché, protože tamější horniny obsahují mnoho jemných popílků s množstvím zirkonů, které lze využít při radioaktivní dataci. Tyto jemné zrnité horniny zároveň také fosilie výborně zachovávají a patrné jsou i detaily. Zdá se, že zástupci této skupiny ediakarské fauny přežili až do začátku kambria.[99]
Biota zahrnuje zástupce kladu Rangeomorpha[101] (jako charnie), které vykazují fraktální růst. Zástupci byli pravděpodobně zachováni in situ, i když to není všeobecně přijato. Zástupci této skupiny byli méně rozmanití než organismy z Ediacara Hills či Namibie. Je možné, že se živili pomocí filtrace vody;[102] většina těchto tvorů zřejmě žila ve vodách příliš hlubokých, než aby v nich mohla probíhat fotosyntéza. S hloubkou vody, jež mohla zpomalit vývoj, možná souvisí i nízká rozmanitost avalonských tvorů. Druhá hypotéza hovoří o tom, že tyto druhy existovaly příliš krátce, než aby se mohla rozvinout bohatá biota. Uznávány jsou oba protichůdné názory.[103]
Skupina ediakarského typu
Skupina ediakarského typu je pojmenována po australském pohoří Ediacara Hills a tvoří ji zkameněliny zachované ve faciích pobřežních lagun a řek. Typicky se nacházejí v červených sádrovcových a vápencových fosilních půdách, které se vytvořily na spraších a náplavových usazeninách během suchého, chladného a mírného paleoklimatu.[63] Většina fosilií se zachovala jako otisky v mikrobiálních půdách,[104] některé i v písečných celcích.[99][103]
Skupina typu Nama
Skupina typu Nama je nejvíce zastoupena v Namibii. Zdejší ediakarské organismy se nejčastěji zachovaly trojrozměrně v písečných vrstvách. Geolog Dima Grazhdankin se domnívá, že tyto fosilie představují organismy žijící v norách,[32] jiný vědec, Guy Narbonne, je toho názoru, že žily na povrchu mořského dna.[105] Tyto vrstvy se nacházejí mezi jednotkami tvořenými pískovci, břidlicemi a prachovci. Nacházejí se zde také mikrobiální povlaky, v nichž jsou obvykle nalézány fosilie.[103]
Význam
V Bílém moři na území Ruska byli nalezeni zástupci všech těchto tři typů ediakarské fauny. To společně s velkým časovým překrytím značí, že pravděpodobně nepředstavovaly různé evoluční fáze či časově odlišné komunity. Jelikož byly fosilie těchto organismů nalezeny na všech kontinentech mimo Antarktidy, geologické hranice se nezdají býti faktorem; stejné fosilie se nalézají u všech tehdy shodných zeměpisných šířek a v oddělených sedimentárních pánvích.[103][106]
Je pravděpodobné, že tyto tři skupiny představují ve skutečnosti tvory adaptované na život v různých prostředích a že zdánlivá vysoká rozmanitost tvarů a stáří je ve skutečnosti dána tím, že se fosilií našlo málo. Analýza jedné pánve v Bílém moři – v níž se opakovaně střídala období, kdy byla částí kontinentálního šelfu a kdy příbřežní zóny – zjistila, že s každým prostředím byla spojená jiná skupina ediakarských organismů.[103]
Vzhledem k tomu, že ediakarská fauna představuje počáteční fázi vývoje mnohobuněčného života, není překvapující, že se nevyvinuly jeho všechny možné způsoby. Na konci ediakary jich bylo vyvinuto ne více než tucet z celkem 92 odhadnutých. V avalonské formaci byly zaznamenány jen čtyři.[107] Největším faktorem omezujícím rozmanitost těchto tvorů byla možná absence tlaku ze strany predátorů.
Odkazy
Reference
V tomto článku byl použit překlad textu z článku Ediacaran_biota na anglické Wikipedii.
- GEHLING, James G.; NARBONNE, Guy M.; ANDERSON, Michael M. The first named Ediacaran body fossil,Aspidella Terranovica. Palaeontology. 2000-09-01, roč. 43, čís. 3, s. 427–456. Dostupné online [cit. 2018-03-24]. ISSN 1475-4983. DOI 10.1111/j.0031-0239.2000.00134.x. (anglicky)
- GÜRICH, G. Die Kuibis-Fossilien der Nama-Formation von Südwestafrika. Palaeontologische Zeitschrift. 1933-08-01, roč. 15, čís. 2–3, s. 137–154. Dostupné online [cit. 2018-03-24]. ISSN 0031-0220. DOI 10.1007/bf03041648. (německy)
- Sprigg RC. Early Cambrian (?) jellyfishes from the Flinders Ranges, South Australia. Transactions of the Royal Society of South Australia. 1947, s. 212–224. (anglicky)
- BRYSON, Bill. A Short History of Nearly Everything. [s.l.]: Doubleday, 2003. ISBN 0-385-60961-2. S. 410. (anglicky)
- Leicester’s fossil celebrity: Charnia and the evolution of early life [online]. Leicester Literary and Philosophical Society Section C (Geology) in conjunction with Dept of Geology, University of Leicester & Leicester Museums and Galleries, 2007 [cit. 2018-03-24]. Dostupné online.
- SPRIGG, R. C. Martin F Glaessner: Palaeontologist extraordinaire. Mem. Geol. Soc. India. 1991, s. 13–20.
- GLAESSNER, M. F. The oldest fossil faunas of South Australia. Geologische Rundschau. 1959-06-01, roč. 47, čís. 2, s. 522–531. Dostupné online [cit. 2018-03-24]. ISSN 0016-7835. DOI 10.1007/bf01800671. (anglicky)
- GLAESSNER, Martin F. Pre-Cambrian Animals. Scientific American. Roč. 204, čís. 3, s. 72–78. Dostupné online. DOI 10.1038/scientificamerican0361-72.
- MISRA, S. B. Late Precambrian (?) Fossils from Southeastern Newfoundland. Geological Society of America Bulletin. 1969-11-01, roč. 80, čís. 11. Dostupné online [cit. 2018-03-24]. ISSN 0016-7606. DOI 10.1130/0016-7606(1969)80[2133:lpffsn2.0.co;2]. (anglicky)
- TERMIER, H.; TERMIER, G. L'Ediacarien, premier etage paleontologique. Rev. Gen. Sci. Et Bull. Assoc. Francaise Avan. Sci. 1960, s. 175–192.
- KNOLL, Andrew; WALTER, Malcolm; NARBONNE, Guy. The Ediacaran Period: a new addition to the geologic time scale. Lethaia. 2006-03-01, roč. 39, čís. 1, s. 13–30. Dostupné online [cit. 2018-03-24]. ISSN 1502-3931. DOI 10.1080/00241160500409223. (anglicky)
- NARBONNE, Guy. The Ediacara biota: A terminal Neoproterozoic experiment in the evolution of life. GSA. S. 1–6. ISSN 1052-5173.
- TARHAN, Lidya G.; HOOD, Ashleigh v S.; DROSER, Mary L. Exceptional preservation of soft-bodied Ediacara Biota promoted by silica-rich oceans. Geology. 2016-11-01, roč. 44, čís. 11, s. 951–954. Dostupné online [cit. 2018-03-24]. ISSN 0091-7613. DOI 10.1130/g38542.1. (anglicky)
- BOWRING, S. A.; MARTIN, M. W. Palaeobiology II. Osney Mead, Oxford: Blackwell Science 583 s. ISBN 9780632051496. OCLC 43945263
- RUNNEGAR, B. N.; FEDONKIN, M. A. The Proterozoic biosphere : a multidisciplinary study. Cambridge: Cambridge University Press, 1992. Dostupné online. ISBN 9780521366151. OCLC 23583672 S. 369–388.
- BURZIN, M. B. & kol. The ecology of the Cambrian radiation. New York: Columbia University Press 525 s. ISBN 0231505167. OCLC 51852000 S. 216–237.
- GEHLING, James G. Microbial Mats in Terminal Proterozoic Siliciclastics: Ediacaran Death Masks. PALAIOS. Roč. 14, čís. 1. Dostupné online [cit. 2018-03-24]. DOI 10.2307/3515360.
- LIU, ALEXANDER G. FRAMBOIDAL PYRITE SHROUD CONFIRMS THE ‘DEATH MASK' MODEL FOR MOLDIC PRESERVATION OF EDIACARAN SOFT-BODIED ORGANISMS. PALAIOS. Roč. 31, čís. 5, s. 259–274. Dostupné online [cit. 2018-03-24]. DOI 10.2110/palo.2015.095.
- CHEN, Jun-Yuan; BOTTJER, David J.; OLIVERI, Paola. Small Bilaterian Fossils from 40 to 55 Million Years Before the Cambrian. Science. 2004-07-09, roč. 305, čís. 5681, s. 218–222. PMID 15178752. Dostupné online [cit. 2018-03-25]. ISSN 0036-8075. DOI 10.1126/science.1099213. PMID 15178752. (anglicky)
- Fossil may be ancestor of most animals. msnbc.com. 2004-06-03. Dostupné online [cit. 2018-03-25]. (anglicky)
- BENGTSON, Stefan; BUDD, Graham. Comment on "Small Bilaterian Fossils from 40 to 55 Million Years Before the Cambrian". Science. 2004-11-19, roč. 306, čís. 5700, s. 1291–1291. PMID 15550644. Dostupné online [cit. 2018-03-25]. ISSN 0036-8075. DOI 10.1126/science.1101338. PMID 15550644. (anglicky)
- DONOGHUE, Philip C. J. Embryonic identity crisis. Nature. 2007/01, roč. 445, čís. 7124, s. 155–156. Dostupné online [cit. 2018-03-25]. ISSN 1476-4687. DOI 10.1038/nature05520. (En)
- BAILEY, Jake V.; JOYE, Samantha B.; KALANETRA, Karen M. Undressing and redressing Ediacaran embryos (Reply). Nature. 2007/04, roč. 446, čís. 7136, s. E10–E11. Dostupné online [cit. 2018-03-25]. ISSN 1476-4687. DOI 10.1038/nature05754. (En)
- KNOLL, A. H.; JAVAUX, E. J.; HEWITT, D. Eukaryotic organisms in Proterozoic oceans. Philosophical Transactions of the Royal Society of London B: Biological Sciences. 2006-06-29, roč. 361, čís. 1470, s. 1023–1038. PMID 16754612. Dostupné online [cit. 2018-03-25]. ISSN 0962-8436. DOI 10.1098/rstb.2006.1843. PMID 16754612. (anglicky)
- CUNNINGHAM, J. A.; THOMAS, C.-W.; BENGTSON, S. Experimental taphonomy of giant sulphur bacteria: implications for the interpretation of the embryo-like Ediacaran Doushantuo fossils. Proc. R. Soc. B. 2012-05-07, roč. 279, čís. 1734, s. 1857–1864. PMID 22158954. Dostupné online [cit. 2018-03-25]. ISSN 0962-8452. DOI 10.1098/rspb.2011.2064. PMID 22158954. (anglicky)
- YIN, Leiming; ZHU, Maoyan; KNOLL, Andrew H. Doushantuo embryos preserved inside diapause egg cysts. Nature. 2007/04, roč. 446, čís. 7136, s. 661–663. Dostupné online [cit. 2018-03-25]. ISSN 1476-4687. DOI 10.1038/nature05682. (En)
- NEWMAN, Stuart A.; FORGACS, Gabor; MULLER, Gerd B. Before programs: The physical origination of multicellular forms. The International Journal of Developmental Biology. 2003-01-01, roč. 50, čís. 2–3, s. 289–299. Dostupné online [cit. 2018-03-25]. ISSN 0214-6282. DOI 10.1387/ijdb.052049sn. (anglicky)
- A., McMenamin, Mark. The garden of Ediacara : discovering the first complex life. New York: Columbia University Press xii, 295 pages s. ISBN 9780231105590. OCLC 37588521
- MICROBIAL ORIGIN OF SOME OF THE EDIACARAN FOSSILS. gsa.confex.com [online]. [cit. 2018-03-25]. Dostupné v archivu pořízeném dne 2014-09-11.
- GRAZHDANKIN, DMITRIY; GERDES, GISELA. Ediacaran microbial colonies. Lethaia. 2007-09-01, roč. 40, čís. 3, s. 201–210. Dostupné online [cit. 2018-03-25]. ISSN 1502-3931. DOI 10.1111/j.1502-3931.2007.00025.x. (anglicky)
- JENSEN, S.; GEHLING, J. G.; DROSER, M. L.; GRANT, S. W. F. A scratch circle origin for the medusoid fossil Kullingia. Lethaia. 2002, roč. 35, čís. 4, s. 291–299. Dostupné online. DOI 10.1080/002411602320790616.
- GRAZHDANKIN, Dmitri; SEILACHER, Adolf. Underground Vendobionta From Namibia. Palaeontology. Roč. 45, čís. 1, s. 57–78. Dostupné online [cit. 2018-03-25]. ISSN 1475-4983. DOI 10.1111/1475-4983.00227. (anglicky)
- MCMENAMIN, M. A. S. Paleotorus: The Laws of Morphogenetic Evolution. [s.l.]: Meanma Press, 2009. ISBN 978-1-893882-18-8.
- SEILACHER, A. Vendobionta and Psammocorallia: lost constructions of Precambrian evolution. Journal of the Geological Society. Roč. 149, čís. 4, s. 607–613. Dostupné online [cit. 2018-03-25]. DOI 10.1144/gsjgs.149.4.0607.
- NARBONNE, Guy M.; SAYLOR, Beverly Z.; GROTZINGER, John P. The Youngest Ediacaran Fossils from Southern Africa. Journal of Paleontology. 1997, roč. 71, čís. 6, s. 953–967. Dostupné online [cit. 2018-03-25].
- XIAO, Shuhai; LAFLAMME, Marc. On the eve of animal radiation: phylogeny, ecology and evolution of the Ediacara biota. Trends in Ecology & Evolution. Roč. 24, čís. 1, s. 31–40. Dostupné online [cit. 2018-03-25]. DOI 10.1016/j.tree.2008.07.015.
- MCMENAMIN, Mark. Osmotrophy in fossil protoctists and early animals. Invertebrate Reproduction & Development. 1993-08-01, roč. 23, s. 165–166. Dostupné online [cit. 2018-03-25]. DOI 10.1080/07924259.1993.9672308.
- LAFLAMME, Marc; XIAO, Shuhai; KOWALEWSKI, Michał. Osmotrophy in modular Ediacara organisms. Proceedings of the National Academy of Sciences. 2009-08-25, roč. 106, čís. 34, s. 14438–14443. Dostupné v archivu pořízeném dne 2020-06-07. DOI 10.1073/pnas.0904836106. Archivováno 7. 6. 2020 na Wayback Machine
- CHEN, Jun-Yuan; OLIVERI, Paola; DAVIDSON, Eric. Response to Comment on "Small Bilaterian Fossils from 40 to 55 Million Years Before the Cambrian". Science. 2004-11-19, roč. 306, čís. 5700, s. 1291–1291. Dostupné online [cit. 2018-03-25]. ISSN 0036-8075. DOI 10.1126/science.1102328. (anglicky)
- FEDONKIN, Mikhail A.; WAGGONER, Benjamin M. The Late Precambrian fossil Kimberella is a mollusc-like bilaterian organism. Nature. 1997/08, roč. 388, čís. 6645, s. 868–871. Dostupné online [cit. 2018-03-25]. ISSN 1476-4687. DOI 10.1038/42242. (En)
- SPRIGGINA IS A TRILOBITOID ECDYSOZOAN. gsa.confex.com [online]. [cit. 2018-03-25]. Dostupné v archivu pořízeném dne 2016-04-12.
- F GLAESSNEE, Martin. Parvancorina — an arthropod from the Late Precambrian (Ediacarian) of South Australia. Ann. Nat. Hist. Mus. Wien. 2018-03-25. Dostupné online [cit. 2018-03-25].
- IVANTSOV, Andrey; MALAKHOVSKAYA, Ya.E.; SEREZHNIKOVA, E.A. Some problematic fossils from the Vendian of the southeastern White Sea Region. Paleontological Journal. 2004-01-01, roč. 38, s. 1–9. Dostupné online [cit. 2018-03-25].
- GERMS, Gerard J. B. New shelly fossils from Nama Group, South West Africa. American Journal of Science. 1972-10-01, roč. 272, čís. 8, s. 752–761. Dostupné online [cit. 2018-03-25]. ISSN 0002-9599. DOI 10.2475/ajs.272.8.752. (anglicky)
- IVANTSOV, Andrey Yu. New find of Cambrian type arthropoda from the Vendian of the White Sea, Russia [online]. Dostupné v archivu pořízeném z originálu.
- FEDONKIN, M. A. Paleoichnology of Vendian Metazoa. In Sokolov, B. S.; Iwanowski, A. B. Vendian System: Historical–Geological and Paleontological Foundation, Vol. 1: Paleontology. Moskva: Nauka, 1985. S. 112–116.
- FEDONKIN, M. A. Origin and early evolution of the Metazoa. New York: Plenum Press 570 s. ISBN 9780306440670. OCLC 25873335 S. 87–129.
- MATZ, Mikhail V.; FRANK, Tamara M.; MARSHALL, N. Justin. Giant Deep-Sea Protist Produces Bilaterian-like Traces. Current Biology. Roč. 18, čís. 23, s. 1849–1854. Dostupné online [cit. 2018-03-25]. DOI 10.1016/j.cub.2008.10.028.
- IVANTSOV, Andrey; MALAKHOVSKAYA, Ya.E. Giant Traces of Vendian Animals. Doklady Earth Sciences. 2002-07-01, roč. 385, s. 618–622. Dostupné online [cit. 2018-03-25].
- IVANTSOV, A. Y. Feeding traces of the Ediacaran animals. www.cprm.gov.br [online]. [cit. 2018-03-25]. Dostupné online.
- https://www.nature.com/articles/d41586-019-02556-x
- FEDONKIN, M. A.; SIMONETTA, A.; IVANTSOV, A. Y. New data onKimberella, the Vendian mollusc-like organism (White Sea region, Russia): palaeoecological and evolutionary implications. Geological Society, London, Special Publications. Roč. 286, čís. 1, s. 157–179. Dostupné online [cit. 2018-03-25]. DOI 10.1144/sp286.12.
- DONOVAN, Stephen K.; LEWIS, David N. Fossils explained 35. Geology Today. 2001-05-01, roč. 17, čís. 3, s. 115–120. Dostupné online [cit. 2018-03-25]. ISSN 1365-2451. DOI 10.1046/j.0266-6979.2001.00285.x. (anglicky)
- WILLIAMS, G. C. Preliminary assessment of the phylogenetics of pennatulacean octocorals, with a reevaluation of Ediacaran frond-like fossils, and a synthesis of the history of evolutionary thought regarding the sea pens. Proceedings of the Sixth International Conference of Coelenterate Biology. [s.l.]: [s.n.], 1997. S. 497–509.
- FORD, T. D. PRE-CAMBRIAN FOSSILS FROM CHARNWOOD FOREST. Proceedings of the Yorkshire Geological Society. Roč. 31, čís. 3, s. 211–217. Dostupné online [cit. 2018-03-25]. DOI 10.1144/pygs.31.3.211.
- LAFLAMME, Marc; NARBONNE, Guy M.; ANDERSON, Michael M. Morphometric analysis of the ediacaran frond charniodiscus from the mistaken point formation, newfoundland. Journal of Paleontology. 2004-09-01, roč. 78, čís. 5, s. 827–837. Dostupné online [cit. 2018-03-25]. ISSN 0022-3360. DOI 10.1666/0022-3360(2004)078%3C0827:maotef%3E2.0.co;2.
- SEILACHER, Adolf; GRAZHDANKIN, Dmitri; LEGOUTA, Anton. Ediacaran biota: The dawn of animal life in the shadow of giant protists. Paleontological Research. 2003-03-31, roč. 7, čís. 1, s. 43–54. Dostupné online [cit. 2018-03-25]. ISSN 1342-8144. DOI 10.2517/prpsj.7.43.
- SEILACHER, A. Patterns of change in earth evolution : report of the Dahlem Workshop on Patterns of Change in Earth Evolution, Berlin, 1983, May 1-6. Berlin: Springer-Verlag 431 s. ISBN 0387127496. OCLC 11202424 S. 159–168.
- SEILACHER, ADOLF. Vendozoa: Organismic construction in the Proterozoic biosphere. Lethaia. 1989-07-01, roč. 22, čís. 3, s. 229–239. Dostupné online [cit. 2018-03-25]. ISSN 1502-3931. DOI 10.1111/j.1502-3931.1989.tb01332.x. (anglicky)
- BUSS, Leo W.; SEILACHER, Adolf. The Phylum Vendobionta: A Sister Group of the Eumetazoa?. Paleobiology. 1994, roč. 20, čís. 1, s. 1–4. Dostupné online [cit. 2018-03-25].
- MCMENAMIN, Mark A. S. The Garden of Ediacara. PALAIOS. Roč. 1, čís. 2. Dostupné online [cit. 2018-03-25]. DOI 10.2307/3514512.
- RETALLACK, Gregory J. Were the Ediacaran Fossils Lichens?. Paleobiology. 1994, roč. 20, čís. 4, s. 523–544. Dostupné online [cit. 2018-03-25].
- RETALLACK, Gregory J. Ediacaran life on land. Nature. 2013/01, roč. 493, čís. 7430, s. 89–92. Dostupné online [cit. 2018-03-25]. ISSN 1476-4687. DOI 10.1038/nature11777. (En)
- WAGGONER, Benjamin M. Ediacaran Lichens: A Critique. Paleobiology. 1995, roč. 21, čís. 3, s. 393–397. Dostupné online [cit. 2018-03-25].
- WAGGONER, Ben; COLLINS, Allen G. Reductio ad absurdum: testing the evolutionary relationships of ediacaran and paleozoic problematic fossils using molecular divergence dates. Journal of Paleontology. 2004-01-01, roč. 78, čís. 1, s. 51–61. Dostupné online [cit. 2018-03-25]. ISSN 0022-3360. DOI 10.1666/0022-3360(2004)078%3C0051:raatte%3E2.0.co;2.
- Australian multicellular fossils point to life on land, not at sea, geologist proposes. ScienceDaily [online]. [cit. 2021-07-29]. Dostupné online. (anglicky)
- RETALLACK, Gregory J. Ediacaran life on land. Nature. 2013-01, roč. 493, čís. 7430, s. 89–92. Dostupné online [cit. 2021-07-29]. ISSN 1476-4687. DOI 10.1038/nature11777. (anglicky)
- WAGGONER, Ben. Interpreting the Earliest Metazoan Fossils: What Can We Learn?. Integrative and Comparative Biology. 1998-12-01, roč. 38, čís. 6, s. 975–982. Dostupné online [cit. 2018-03-25]. ISSN 1540-7063. DOI 10.1093/icb/38.6.975. (anglicky)
- FORD, Trevor D. Topics The Precambrian fossils of Charnwood Forest. Geology Today. 1999-12-01, roč. 15, čís. 6, s. 230–234. Dostupné online [cit. 2018-03-25]. ISSN 1365-2451. DOI 10.1046/j.1365-2451.1999.00007.x. (anglicky)
- ZHURALEV. Were Vend-Ediacaran multicellulars metazoa?. 29th International Geological Congress, Kyoto, Japan. [s.l.]: [s.n.], 1992. S. 339.
- PETERSON, Kevin J.; WAGGONER, Ben; HAGADORN, James W. A Fungal Analog for Newfoundland Ediacaran Fossils?. Integrative and Comparative Biology. 2003-02-01, roč. 43, čís. 1, s. 127–136. Dostupné online [cit. 2018-03-25]. ISSN 1540-7063. DOI 10.1093/icb/43.1.127. (anglicky)
- PFLUG. Zur fauna der Nama-Schichten in Südwest-Afrika. IV. Mikroscopische anatomie der petalo-organisme. Paleontographica. 1973, s. 166–202.
- JUST, Jean; KRISTENSEN, Reinhardt Møbjerg; OLESEN, Jørgen. Dendrogramma, New Genus, with Two New Non-Bilaterian Species from the Marine Bathyal of Southeastern Australia (Animalia, Metazoa incertae sedis) – with Similarities to Some Medusoids from the Precambrian Ediacara. S. 1–11. PLoS ONE [online]. 3. září 2014. Svazek 9, čís. 9: e102976, s. 1–11. Dostupné online. ISSN 1932-6203. DOI 10.1371/journal.pone.0102976. (anglicky)
- New deep sea mushroom-shaped organisms discovered. PhysOrg [online]. 3. září 2014. Dostupné online. (anglicky)
- GOUGH, Myles. Origin of mystery deep-sea mushroom revealed. BBC News [online]. 7. červen 2016. Dostupné online. (anglicky)
- O'HARA, Tim. Solved mystery of the deep-sea mushroom just raises new questions. The Conversation [online]. 7. červen 2016. Dostupné online. Dostupné také na: . (anglicky)
- SCHOPF, J. W.; PACKER, B. M. Early Archean (3.3-billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia. Science. 1987-07-03, roč. 237, čís. 4810, s. 70–73. PMID 11539686. Dostupné online [cit. 2018-03-25]. ISSN 0036-8075. DOI 10.1126/science.11539686. PMID 11539686. (anglicky)
- HOFMANN, H. J.; GREY, K.; HICKMAN, A. H. Origin of 3.45 Ga coniform stromatolites in Warrawoona Group, Western Australia. GSA Bulletin. 1999-08-01, roč. 111, čís. 8. Dostupné online [cit. 2018-03-25]. ISSN 0016-7606. DOI 10.1130/0016-7606(1999)111%3C1256:oogcsi%3E2.3.co;2. (anglicky)
- ARCHER, Corey; VANCE, Derek. Coupled Fe and S isotope evidence for Archean microbial Fe(III) and sulfate reduction. Geology. 2006-03-01, roč. 34, čís. 3. Dostupné online [cit. 2018-03-25]. ISSN 0091-7613. DOI 10.1130/g22067.1. (anglicky)
- BUTTERFIELD, Nicholas J. Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes. Paleobiology. 2000-09-01, roč. 26, čís. 3, s. 386–404. Dostupné online [cit. 2018-03-25]. ISSN 0094-8373. DOI 10.1666/0094-8373(2000)0262.0.CO;2.
- FEDONKIN, M. A. New representatives of the Precambrian coelenterates in the northern Russian platform. Paleontologicheskii Zhurnal. S. 7–15. ISSN 0031-031X.
- MEERT, Joseph G.; GIBSHER, Anatoly S.; LEVASHOVA, Natalia M. Glaciation and ~770Ma Ediacara (?) Fossils from the Lesser Karatau Microcontinent, Kazakhstan. Gondwana Research. Roč. 19, čís. 4, s. 867–880. Dostupné online [cit. 2018-03-25]. DOI 10.1016/j.gr.2010.11.008.
- CANFIELD, Donald E.; TESKE, Andreas. Late Proterozoic rise in atmospheric oxygen concentration inferred from phylogenetic and sulphur-isotope studies. Nature. 1996/07, roč. 382, čís. 6587, s. 127–132. Dostupné online [cit. 2018-03-25]. ISSN 1476-4687. DOI 10.1038/382127a0. (En)
- CANFIELD, Don E.; POULTON, Simon W.; NARBONNE, Guy M. Late-Neoproterozoic Deep-Ocean Oxygenation and the Rise of Animal Life. Science. 2007-01-05, roč. 315, čís. 5808, s. 92–95. PMID 17158290. Dostupné online [cit. 2018-03-25]. ISSN 0036-8075. DOI 10.1126/science.1135013. PMID 17158290. (anglicky)
- FIKE, D. A.; GROTZINGER, J. P.; PRATT, L. M. Oxidation of the Ediacaran Ocean. Nature. 2006/12, roč. 444, čís. 7120, s. 744–747. Dostupné online [cit. 2018-03-25]. ISSN 1476-4687. DOI 10.1038/nature05345. (En)
- BUTTERFIELD, N. J. Oxygen, animals and oceanic ventilation: an alternative view. Geobiology. 2009-01-01, roč. 7, čís. 1, s. 1–7. Dostupné online [cit. 2018-03-25]. ISSN 1472-4669. DOI 10.1111/j.1472-4669.2009.00188.x. (anglicky)
- NARBONNE, Guy. Life after Snowball: The Mistaken Point biota and the origin of animal ecosystems. Geological Society of America Abstracts with Programs. 2003, s. 516.
- SHEN, Bing; DONG, Lin; XIAO, Shuhai. The Avalon Explosion: Evolution of Ediacara Morphospace. Science. 2008-01-04, roč. 319, čís. 5859, s. 81–84. PMID 18174439. Dostupné online [cit. 2018-03-25]. ISSN 0036-8075. DOI 10.1126/science.1150279. PMID 18174439. (anglicky)
- Ediacaran-like fossils in Cambrian Burgess Shale-type faunas of North America | The Palaeontological Association. www.palass.org [online]. [cit. 2018-03-25]. Dostupné online. (anglicky)
- SHU, D.-G.; MORRIS, S. Conway; HAN, J. Lower Cambrian Vendobionts from China and Early Diploblast Evolution. Science. 2006-05-05, roč. 312, čís. 5774, s. 731–734. PMID 16675697. Dostupné online [cit. 2018-03-25]. ISSN 0036-8075. DOI 10.1126/science.1124565. PMID 16675697. (anglicky)
- HAN, Jian; LI, Guoxiang; WANG, Xing. Dwarfed vendobionts from the Cambrian Kuanchuanpu Formation in South China. In: [s.l.]: [s.n.], 2016-07-11. Dostupné online.
- STANLEY, S. M. An Ecological Theory for the Sudden Origin of Multicellular Life in the Late Precambrian. Proceedings of the National Academy of Sciences. Roč. 70, čís. 5, s. 1486–1489. Dostupné v archivu pořízeném dne 2020-06-07. DOI 10.1073/pnas.70.5.1486. (anglicky) Archivováno 7. 6. 2020 na Wayback Machine
- GOULD, Stephen Jay; CALLOWAY, C. Bradford. Clams and Brachiopods-Ships that Pass in the Night. Paleobiology. 1980, roč. 6, čís. 4, s. 383–396. Dostupné online [cit. 2018-03-25].
- MCKERROW, W. S.; SCOTESE, C. R.; BRASIER, M. D. Early Cambrian continental reconstructions. Journal of the Geological Society. Roč. 149, čís. 4, s. 599–606. Dostupné online [cit. 2018-03-25]. DOI 10.1144/gsjgs.149.4.0599.
- HALLAM, A. Pre-Quaternary Sea-Level Changes. Annual Review of Earth and Planetary Sciences. 1984-05-01, roč. 12, čís. 1, s. 205–243. Dostupné online [cit. 2018-03-25]. ISSN 0084-6597. DOI 10.1146/annurev.ea.12.050184.001225.
- BRASIER, M. D. Background to the Cambrian Explosion. Journal of the Geological Society. Roč. 149, čís. 4, s. 585–587. Dostupné online [cit. 2018-03-25]. DOI 10.1144/gsjgs.149.4.0585.
- BRASIER, M. D. Global ocean—atmosphere change across the Precambrian—Cambrian transition. Geological Magazine. 1992/03, roč. 129, čís. 2, s. 161–168. Dostupné online [cit. 2018-03-25]. ISSN 1469-5081. DOI 10.1017/s0016756800008256. (anglicky)
- LOWENSTEIN, Tim K.; TIMOFEEFF, Michael N.; BRENNAN, Sean T. Oscillations in Phanerozoic Seawater Chemistry: Evidence from Fluid Inclusions. Science. 2001-11-02, roč. 294, čís. 5544, s. 1086–1088. PMID 11691988. Dostupné online [cit. 2018-03-25]. ISSN 0036-8075. DOI 10.1126/science.1064280. PMID 11691988. (anglicky)
- ERWIN, Douglas H. Wonderful Ediacarans, wonderful cnidarians?. Evolution & Development. 2008-05-01, roč. 10, čís. 3, s. 263–264. Dostupné online [cit. 2018-03-25]. ISSN 1525-142X. DOI 10.1111/j.1525-142x.2008.00234.x. (anglicky)
- BENUS. Trace fossils, small shelly fossils, and the Precambrian-Cambrian boundary : proceedings, August 8-18, 1987, Memorial University. Albany: University of the State of New York ISBN 9781555571788. OCLC 18527555
- CLAPHAM, Matthew E.; NARBONNE, Guy M.; GEHLING, James G. Paleoecology of the Oldest Known Animal Communities: Ediacaran Assemblages at Mistaken Point, Newfoundland. Paleobiology. 2003, roč. 29, čís. 4, s. 527–544. Dostupné online [cit. 2018-03-25].
- CLAPHAM, M.E.; NARBONNE, G.M. Ediacaran epifaunal tiering. Geology. 2002-07-01, roč. 30, čís. 7. Dostupné online [cit. 2018-03-25]. ISSN 0091-7613. DOI 10.1130/0091-7613(2002)030%3C0627:eet%3E2.0.co;2. (anglicky)
- GRAZHDANKIN, Dmitriy. Patterns of distribution in the Ediacaran biotas: Facies versus biogeography and evolution. Paleobiology. 2004-06-01, roč. 30, s. 203–221. Dostupné online [cit. 2018-03-25]. DOI 10.1666/0094-8373(2004)0302.0.CO;2.
- RETTALACK, G. J. Criteria for distinguishing microbial mats and earths. Society of Economic Paleontologists and Mineralogists Special Paper. Tulsa: Society of Economic Paleontologists and Mineralogists S. 136–152.
- NARBONNE, Guy M. THE EDIACARA BIOTA: Neoproterozoic Origin of Animals and Their Ecosystems. Annual Review of Earth and Planetary Sciences. 2004-09-24, roč. 33, čís. 1, s. 421–442. Dostupné online [cit. 2018-03-25]. ISSN 0084-6597. DOI 10.1146/annurev.earth.33.092203.122519.
- WAGGONER, Ben. Biogeographic Analyses of the Ediacara Biota: A Conflict with Paleotectonic Reconstructions. Paleobiology. 1999, roč. 25, čís. 4, s. 440–458. Dostupné online [cit. 2018-03-25].
- BAMBACH, Richard K.; BUSH, Andrew M.; ERWIN, Douglas H. AUTECOLOGY AND THE FILLING OF ECOSPACE: KEY METAZOAN RADIATIONS. Palaeontology. 2007-1, roč. 50, čís. 1, s. 1–22. Dostupné online [cit. 2019-08-26]. ISSN 0031-0239. DOI 10.1111/j.1475-4983.2006.00611.x. (anglicky)
Externí odkazy
 Obrázky, zvuky či videa k tématu Ediakarská fauna na Wikimedia Commons
Obrázky, zvuky či videa k tématu Ediakarská fauna na Wikimedia Commons