Žahavá buňka
Žahavé buňky, též zvané knidocyty (z řec. knide, kopřiva) nebo nematocyty (z řec. nema, vlákno), jsou specializované buňky, které se nacházejí na povrchu těla žahavců. Obsahují organely, které po podráždění vymršťují z buňky harpunovitá vlákna a vypouštějí toxiny. Žahavcům tyto buňky slouží zejména k uchvácení kořisti a k obraně proti predátorům.
Stavba žahavé buňky
Žahavé buňky se tvoří v průběhu celého života z intersticiálních buněk. Vyvíjející se žahavá buňka se nazývá knidoblast. Z Golgiho aparátu v ní postupně vzniká zvláštní organela, tzv. knidocysta (též knida). Jedná se typicky o váček naplněný toxiny, který ústí na povrchu buňky a v klidovém stavu je uzavřen víčkem (operculum). Ve váčku je dále stočeno duté vlákno, na němž se mohou nacházet různé háčky či jiné povrchové struktury.
Na povrchu žahavé buňky se nachází citlivý výběžek (knidocil), který slouží jako mechanoreceptor, jehož podráždění může aktivovat knidocystu a vyvolat vymrštění vlákna.
Vývin žahavých buněk probíhá uvnitř těla žahavců. Těsně před dozráním pak tyto buňky migrují na místo určení (například na povrch chapadel), kde se formují do tzv. baterií obsahujících obvykle několik typů knidocytů sdružených kolem smyslových buněk.[1] Žahavé buňky jsou určeny jen k jednomu použití, po vymrštění vlákna odumírají a musí být nahrazeny novými.
Typy knidocyst

Na základě morfologie rozlišujeme tři základní typy knidocyst:
- nematocysty – vyskytují se u všech skupin žahavců, a to zejména na chapadlech a v okolí ústního otvoru. Patří sem typické velké knidy obsahující toxiny a harpunovitě zakončené vlákno, které slouží k lovu kořisti. Na základě vzhledu vlákna (otevřený či uzavřený konec, háčky a další povrchové struktury) se nematocysty dále dělí do více než dvaceti skupin.[2]
- spirocysty – vyskytují se pouze u korálnatců (Anthozoa). Obsahují velmi dlouhá a silná přilnavá vlákna bez výrazných povrchových struktur. Slouží k zachycení kořisti, do které je zároveň z nematocyst vpravován toxin.
- ptychocysty – vyskytují se pouze u červnatců (Ceriantharia), kteří je používají především k přichycení povrchu a také při tvorbě trubičky, ve které žijí. Obsahují tenká vlákna, která se po vymrštění spojují do sítí.[3]
Někdy se uvádí též jiné dělení podle funkce vláken:
- penetranty – vlákna s ostrým koncem, kterým pronikají do kořisti. Jedná se vždy o nematocysty.
- volventy – ovíjivá vlákna. Může se jednat o spirocysty, ale patří sem i nematocysty s uzavřeným koncem vlákna.
- glutinanty – lepivá vlákna. Někdy synonymum ptychocyst, ale přilnavé mohou být opět i některé typy nematocyst.
U různých druhů žahavců se nacházejí různé typy knidocytů, celkový soubor všech typů žahavých buněk určitého druhu se někdy nazývá knidom.
Funkce
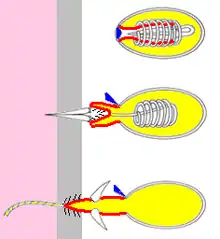
Modře je označeno operculum,
červeně bazální část vlákna, která se během reakce obrací naruby,
žlutě tekutý obsah cysty,
šedě povrch těla kořisti.
Po dostatečném podráždění žahavé buňky dojde k otevření víčka a vymrštění vlákna. U jeho báze se mohou nacházet bodce (stylety), které pronikají do těla kořisti, kde se vlákno dále rozvíjí a vypouští se přes něj toxický obsah knidocysty.
Vymrštění vlákna z knidocysty může být vyvoláno různými podněty[4] :
- mechanicky – například přímý kontakt s kořistí či detekce jejího pohybu v blízkosti žahavce
- chemicky – zachycení "pachu" kořisti nebo látek, které začnou unikat z jejího těla po prvním zásahu
- na základě signálu z nervové soustavy
Jednotlivé druhy stimulů působí v synergii – pokud se například k pouhému mechanickému podráždění přidá i chemické, výrazně se zvýší frekvence vystřelování vláken. Naopak pokud je žahavec již nasycen, molekuly uvolňované při trávení potravy mohou chemicky brzdit aktivaci žahavých buněk i když je kořist stále přítomna.
Samotná reakce je extrémně rychlá – vymrštění vlákna trvá maximálně několik mikrosekund, při zásahu kořisti může špička vlákna dosahovat rychlosti přes 30 m/s a zrychlení až 5 milionů g, proto bez problémů projde i pevnou kutikulou.[5]
Přesný mechanismus reakce je stále předmětem zkoumání. Bylo zjištěno, že těsně po podráždění dojde k výrazné změně pH uvnitř knidocysty a přes membránu, která ji odděluje od cytoplazmy žahavé buňky, začnou přecházet protony vodíku a vápenaté kationty.[6] Následně začne do knidocysty osmoticky pronikat voda z cytoplazmy, čímž dojde ke zvětšení objemu knidocysty, otevření víčka a vymrštění vlákna.
Toxické účinky
Chemické složení tekutého obsahu knidocysty, a tedy i jeho působení na zasažené organismy, je různé u různých druhů žahavců. Mezi hlavní účinné látky nalezené v knidocystách patří[7] :
- enzymy (fosfolipáza, metaloproteázy) – rozkládají složky buněčných membrán i mezibuněčné hmoty a tím přímo poškozují zasažené tkáně
- toxiny tvořící póry (Pore forming toxins – PFT) – shlukují se v cytoplazmatické membráně a vytváří v ní otvory, kterými začnou nekontrolovaně procházet ionty a malé molekuly, což vede k narušení vnitřních procesů a následně zániku napadené buňky
- neurotoxiny – blokují iontové kanály nervových buněk, což vyvolá velmi rychlou paralýzu kořisti
- neproteinové menší molekuly (serotonin, histamin aj.) – způsobují bolest a rozšíření cév, které urychluje účinek toxinů
Toxiny často účinkují zejména na typickou kořist, např. členovce, a jiné skupiny příliš nepoškozují. Pro člověka je nebezpečný kontakt s některými druhy medúz a korálnatců, a především se čtyřhrankami, jejichž jed může člověku přivodit kardiovaskulární kolaps a smrt již několik minut po požahání.[8]
Účinné látky z knidocyst žahavců jsou ale také předmětem farmakologických výzkumů s potenciálem pro léčbu neurodegenerativních onemocnění a některých typů rakoviny, či pro tlumení bolesti.[7]
Využití žahavých buněk
Žahavci používají knidocyty k několika účelům:
- lov potravy – hlavní účel žahavých buněk, jsou k němu využívány všechny jejich typy. Např. nezmaři kořist často nejprve zachytí a znehybní ovíjivými vlákny, a teprve poté se aktivují penetranty a do kořisti jsou vpuštěny toxiny.
- boj s predátory a konkurenty – žahavé buňky jsou samozřejmě účinnou obranou proti predátorům. U sasanek bylo také pozorováno jejich používání v konkurenčním boji. Pokud se v jejich blízkosti usadil jiný, nepříbuzný jedinec, aktivně jej napadaly zvláštními dlouhými chapadly, která obsahují mnoho velikých žahavých buněk, a dokonce z těchto chapadel odvrhovaly na konkurenta žahavou tkáň, což vedlo k jeho výraznému poškození.[1]
- přichycení k podkladu a pohyb – červnatci používají vlákna produkovaná ptychocystami k trvalejšímu přichycení k podkladu. Nezmaři se běžně pohybují na delší vzdálenosti tak, že dělají "salta", při kterých se dočasně přichytí k povrchu pomocí lepivých vláken na chapadlech a přemístí nožní terč.
Kleptoknidy

Jedinou skupinou živočichů, která vytváří žahavé buňky, jsou žahavci. Někteří jejich predátoři (nahožábří plži, ploštěnci či žebernatky) jsou však schopni nejen odolat účinku žahavých buněk a zkonzumovat žahavce, ale navíc i izolovat jejich nepoužité knidocysty, zabudovat je do buněk vlastní pokožky a používat k obraně.[9] Tyto "odcizené" knidocysty se nazývají kleptoknidy (z řec. kleptein, krást).
Reference
- KASS-SIMON, G.; SCAPPATICCI, JR, A.A. The behavioral and developmental physiology of nematocysts. Canadian Journal of Zoology. 2002, roč. 80, s. 1772–1794. Dostupné v archivu pořízeném dne 2016-08-16. DOI 10.1139/Z02-135.
- ÖSTMAN, Carina. A guideline to nematocyst nomenclature and classification, and some notes on the systematic value of nematocysts. Scientia Marina. 2000, čís. 64, s. 31–46. Dostupné online [cit. 2016-05-01].
- MARISCAL, Richard N. et al. The Ptychocyst, a Major New Category of Cnida Used in Tube Construction by a Cerianthid Anemone. Biological Bulletin. 1977, roč. 152, čís. 3, s. 392–405. Dostupné online [cit. 2016-05-01]. DOI 10.2307/1540427.
- CANNON, Quinn; WAGNER, Eric. Comparison of Discharge Mechanisms of Cnidarian Cnidae and Myxozoan Polar Capsules. Reviews in Fisheries Science. 2003, roč. 11, čís. 3, s. 185–219. Dostupné v archivu pořízeném dne 2016-12-21. DOI 10.1080/10641260390244305.
- NÜCHTER, Timm et al. Nanosecond-scale kinetics of nematocyst discharge. Current Biology. 2006, roč. 16, čís. 9. Dostupné online [cit. 2016-05-01]. DOI 10.1016/j.cub.2006.03.089.
- BERKING, Stefan; HERRMANN, Klaus. Formation and discharge of nematocysts is controlled by a proton gradient across the cyst membrane. Helgoland Marine Research. 2006, roč. 60, čís. 3, s. 180–188. Dostupné v archivu pořízeném dne 2016-03-04. DOI 10.1007/s10152-005-0019-y.
- JOUIAEI, M. et al. Ancient Venom Systems: A Review on Cnidaria Toxins. Toxins. 2015, čís. 7, s. 2251–2271. Dostupné online [cit. 2016-05-01]. DOI 10.3390/toxins7062251.
- YANAGIHARA, A. A.; SHOHET, R. V. Cubozoan Venom-Induced Cardiovascular Collapse Is Caused by Hyperkalemia and Prevented by Zinc Gluconate in Mice. PLoS ONE. 2012. Dostupné online. DOI 10.1371/journal.pone.0051368.
- GREENWOOD, Paul E. Acquisition and Use of Nematocysts by Cnidarian Predators. Toxicon. 2009, roč. 54, čís. 8, s. 1065–1070. Postprint dostupný na adrese http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2783962/. DOI 10.1016/j.toxicon.2009.02.029.
Externí odkazy
 Obrázky, zvuky či videa k tématu žahavá buňka na Wikimedia Commons
Obrázky, zvuky či videa k tématu žahavá buňka na Wikimedia Commons