Bílkovina
Bílkoviny, odborně proteiny (z řeckého πρώτειος próteios „prvotní, primární, hlavní“), patří mezi biopolymery. Proteiny se řadí mezi makromolekuly, které obsahují jeden nebo více dlouhých řetězců aminokyselinových zbytků. Plní v organismech širokou škálu funkcí, včetně katalýzy metabolických reakcí, replikace DNA, reakce na podněty, poskytování struktury buňkám a organismům a také transport molekul z jedné polohy do polohy druhé.

Proteiny se od sebe navzájem liší především svou sekvencí aminokyselin, která je dána nukleotidovou sekvencí jejich genů, a které obvykle vede ke sbalení proteinu do specifických 3D struktur, které určují jeho aktivitu. Lineární řetězec aminokyselinových zbytků se nazývá polypeptid. Proteiny obsahují alespoň jeden dlouhá polypeptid. Krátké polypeptid, obsahující méně než 20–30 zbytků jsou zřídka považovány za proteiny a často se nazývají peptidy nebo někdy oligopeptidy. Jednotlivé aminokyselinové zbytky jsou spolu vázány peptidovými vazbami a sousedními aminokyselinovými zbytky. sekvence aminokyselinových zbytků v proteinu je definována sekvencí genu, který je zakódován v genetickém kódu. Obecně genetický kód specifikuje 20 standardních aminokyselin, ale u určitých organismů může genetický kód zahrnovat selenocystein a pyrrolysin. Krátce po syntéze nebo dokonce během syntézy jsou zbytky v proteinu často chemicky modifikovány posttranslační modifikací, která mění fyzikální a chemické vlastnosti, skládání, stabilitu, aktivitu a nakonec i funkci proteinů. Některé proteiny mají připojené nepeptidové skupiny, které lze nazvat prostetické skupiny nebo kofaktory. Proteiny mohou také spolupracovat na dosažení konkrétní funkce a často se spojují a vytvářejí stabilní proteinové komplexy.
Jakmile se proteiny vytvoří, existují pouze po určitou dobu a pak jsou degradovány a recyklovány buněčným aparátem prostřednictvím procesu přeměny proteinů. Životnost proteinu se měří podle jeho poločasu rozpadu a pokrývá široký rozsah. Mohou existovat minuty nebo roky s průměrnou délkou života 1–2 dny v savčích buňkách. Abnormální nebo špatně poskládané proteiny jsou degradovány rychleji buď kvůli tomu, že jsou cíleny pro destrukci, nebo kvůli tomu, že jsou nestabilní.
Jedná se o vysokomolekulární přírodní látky s relativní molekulovou hmotností 103 až 106 složené z aminokyselin spojených peptidovou vazbou mezi karboxylem jedné aminokyseliny a aminoskupinou následující aminokyseliny.
Proteiny jsou podstatou všech živých organismů. Jejich základní povahu rozpoznal Henri Braconnot již v roce 1820 při zahřívání klihu s kyselinou sírovou. Podrobněji strukturu bílkovin popsali Hermann Emil Fischer a Linus Pauling.
Stejně jako jiné biologické makromolekuly, jako jsou polysacharidy a nukleové kyseliny, jsou proteiny nezbytnou součástí organismů a účastní se prakticky všech procesů v buňkách. Mnoho proteinů jsou enzymy, které katalyzují biochemické reakce a jsou životně důležité pro metabolismus. Proteiny mají také strukturální nebo mechanické funkce, jako je aktin a myosin ve svalu a proteiny v cytoskeletu, které tvoří systém lešení, který udržuje tvar buňky. Jiné proteiny jsou důležité v buněčné signalizaci, imunitních reakcích, buněčné adhezi a buněčném cyklu. U zvířat jsou bílkoviny potřebné ve stravě, aby poskytly esenciální aminokyseliny, které nemohou být syntetizovány. Trávení rozkládá proteiny pro metabolické využití.
Proteiny mohou být purifikovány z jiných buněčných složek použitím různých technik, jako je ultracentrifugace, precipitace, elektroforéza a chromatografie; příchod genetického inženýrství umožnil řadu metod pro usnadnění čištění. Metody běžně používané ke studiu struktury a funkce proteinu zahrnují imunohistochemii, místně řízenou mutagenezi, rentgenovou krystalografii, nukleární magnetickou rezonanci a hmotnostní spektrometrii.
Historie
Proteiny byly uznány jako odlišná třída biologických molekul v 18. století Antoinem Fourcroyem.
Poprvé byli bílkoviny popsány holandským chemikem Gerardusem Johannesem Mulderem a pojmenovány švédským chemikem Jönsem Jacobem Berzeluisem v roce 1838.[1][2] Mulder provedl elementární analýzu běžných proteinů a zjistil, že téměř všechny proteiny mají stejný empirický vzorec, C400H620N100O120P1S1.[3] Došel k mylnému názoru, že mohou být složeny z jediného typu (velmi velké) molekuly. Termín "protein" pro popis těchto molekul navrhl Mulderův spolupracovník Berzelius; protein je odvozen z řeckého slova πρώτειος (proteios), což znamená „primární“[4] v čele“ nebo „stát vpředu.“ [5] Mulder pokračoval v identifikaci produktů degradace proteinů, jako je aminokyselina leucin, pro kterou našel (téměř správnou) molekulovou hmotnost 131 Da.[3] Před „proteinem“ se používaly jiné názvy, jako „albuminy“ nebo „albuminové materiály“ (Eiweisskörper, v němčině).[6]
Raní vědci jako německý vědec Carl von Voit zabývající se výživou věřili, že bílkoviny jsou nejdůležitější živinou pro udržení struktury těla, jelikož se obecně věřilo, že „maso dělá maso.“ [7] Karl Heinrich Ritthausen rozšířil známe proteinové formy s identifikací tzv. kyseliny glutamové. Na zemědělské experimentální stanici Connecticut sestavil Thomas Burr Osborne detailní přehled rostlinných proteinů. Ve spolupráci s Lafayttem Mendelem a aplikací Leibigova zákona minima při krmení laboratorních potkanů byly stanoveny nutriční esenciální aminokyseliny. V práci pokračoval a komunikoval s Williamem Cummingem Rosem. K pochopení proteinů jako polypeptid došlo díky práci Franze Hofmeistra a Hermanna Emila Fishera v roce 1902. Hlavní role proteinů jako enzymů v živých organismech byla plně doceněna až v roce 1926, kdy James B. Sumner ukázal, že enzym ureáza je ve skutečnosti protein.[8]
První studie se zaměřily na proteiny, které by mohly být purifikovány ve velkém množství, např. ty z krve, vaječného bílku, různých toxinů a trávicích/metabolických enzymů získaných z jatek. V 50. letech 20. století společnost Armor Hot Dog Co. vyčistila 1 kg čisté hovězí pankreatické ribonukleázy A a dala ji volně k dispozici vědcům; toto gesto pomohlo ribonukleáze A stát se hlavním cílem biochemického studia v následujících desetiletích.[3]
Linus Pauling je připočítán s úspěšnou predikcí pravidelných proteinových sekundárních struktur založených na vodíkových vazbách, což byla myšlenka, kterou poprvé předložil William Astbury v roce 1933.[5] Pozdější práce Waltera Kauzmanna o denaturaci[9][10] částečně založené na předchozích studiích Kaj Linderstrøm-Langa[11] přispěly k pochopení skládání proteinů a struktury zprostředkované hydrofobními interakcemi.
Prvním proteinem, který byl sekvenován, byl inzulin, který sekventoval Frederick Sanger v roce 1949. Sanger správně určil aminokyselinovou sekvenci inzulinu, čímž přesvědčivě prokázal, že proteiny se skládaly spíše z lineárních polymerů aminokyselin než z rozvětvených řetězců, koloidů nebo cyklů.[12] Za tento úspěch získal v roce 1958 Nobelovu cenu.[13]
S rozvojem rentgenové krystalografie bylo možné sekvenovat proteinové struktury.[14] První proteinové struktury, které byly vyřešeny, byly hemoglobin od Maxe Perutze a myoglobin od sira Johna Cowderyho Kendrewa v roce 1958.[15][16] Využití počítačů a zvyšující se výpočetní výkon podpořily i sekvenování komplexních proteinů. V roce 1999 Roger Kornberg uspěl v sekvenování vysoce komplexní struktury RNA polymerázy pomocí vysoce intenzivního rentgenového záření ze synchrotronů.[14]
Od té doby byla vyvinuta kryo-elektronová mikroskopie velkých makromolekulárních celků.[17] Cryo-EM používá vzorky proteinů, které jsou zmrazené spíše než krystaly, a paprsky elektronů spíše než rentgenové záření. Způsobuje menší poškození vzorku, což umožňuje vědcům získat více informací a analyzovat větší struktury. Výpočetní predikce proteinové struktury malých proteinových domén[18] také pomohla výzkumníkům přiblížit se k rozlišení proteinových struktur na atomární úrovni. Od roku 2017 má Protein Data Bank více než 126 060 struktur proteinů s atomárním rozlišením.[19]
Základní vlastnosti bílkovin
V proteinech jsou aminokyseliny vzájemně vázány aminoskupinami –NH2 a karboxylovými skupinami –COOH amidovou vazbou –NH–CO– (amidy), která se v případě proteinů nazývá peptidová vazba.
Podle počtu aminokyselin, které jsou v molekule takto navázány rozlišujeme
- oligopeptidy (obsahují 2–10 aminokyselin)
- polypeptidy (obsahují 11–100 aminokyselin, podle některých zdrojů 11–50 aminokyselin )
- vlastní bílkoviny – proteiny (více než 100 aminokyselin, podle některých více než 50 aminokyselin).
Nejednotnost hranice mezi peptidy a bílkovinami je dána tím, že dříve platilo: do počtu 50 aminokyselin se jedná o peptid, při vyšším počtu pak o bílkovinu. V současnosti je posuzována poměrná molekulová hmotnost (Mr), kdy do hodnoty Mr=10 000 jde o peptid, nad tuto hodnotu o bílkovinu. To odpovídá zhruba 100 aminokyselinám.
Pořadí aminokyselin v řetězci proteinu se označuje jako primární struktura nebo také sekvence. Z 20 obvyklých proteinogenních aminokyselin, které se vždy vyskytují v lidském organismu, může v případě jednoduchého proteinu, složeného ze 100 aminokyselin, vzniknout 20100 (tedy zhruba 10130) rozdílných primárních proteinových struktur. Z toho vyplývá, že může existovat nesrovnatelně větší množství různých proteinů, než je jich obsaženo ve všech živých organismech na Zemi. Struktura mnoha proteinů, například myoglobinu a hemoglobinu, je již známá; u blízce příbuzných živočišných druhů jsou si struktury velmi podobné.
Molekuly proteinů mohou vytvářet protáhlé, vláknité, ve vodě nerozpustné struktury, skleroproteiny (též nazývané fibrilární), a kulovité nebo elipsoidní, ve vodě rozpustné sferoproteiny (též nazývané globulární). V protikladu ke skleroproteinům (kolagen, keratin, fibroin, tvořící vlasy, rohovinu, chrupavky…) lze téměř u všech sferoproteinů (například enzymy, svalová tkáň) varem nebo působením kyselin a louhů (změnou hodnoty pH) rozrušit jejich terciární a sekundární strukturu (srážení, denaturace). Přitom se ztrácejí některé biologické vlastnosti proteinů, jako schopnost enzymů štěpit potravu nebo vyvolávat svalovou kontraktivitu. Tělu cizí proteiny vyvolávají svou přítomností reakci antigen–protilátka, a proto nesmí být nikdy přímo vpraveny do krevního oběhu.
Struktura bílkovin

Rozlišujeme primární, sekundární, terciární a u některých složitějších proteinů ještě kvartérní strukturu bílkovinových řetězců.
Primární struktura
Primární struktura je dána pořadím aminokyselin v polypeptidovém řetězci. Standardně se zapisuje od N-konce k C-konci proteinu. První určení primární struktury provedl v roce 1953 Frederick Sanger. Primární struktura udává chemické vlastnosti bílkoviny a také determinuje vyšší struktury, viz článek skládání bílkovin.
Sekundární struktura
Sekundární struktura je geometrické uspořádání polypeptidového řetězce „na krátké vzdálenosti“, tzn. mezi několika po sobě jdoucími aminokyselinami. První studie sekundárních struktur proběhly v 30. a 40. letech 20. století. Jsou rozpoznávány různé druhy těchto stavebních motivů: alfa šroubovice (alfa-helix), struktura skládaného listu (beta-sheet), neuspořádaná struktura (random coil) a další.
Někdy se definuje i "strukturní motiv" nebo supersekundární struktura jako přechod mezi sekundární a terciární strukturou, což je jednoduché uspořádání několika sekundárních struktur, jako je řecký klíč složený z několika beta listů, beta-vlásenka (beta-hairpin) ze dvou beta listů a otočky nebo helix-smyčka-helix tvořící základ jedné z významných rodin transkripčních faktorů (nazývaných v angličtině basic helix-loop-helix).
Terciární struktura
Tímto pojmem se označuje trojrozměrné uspořádání celého peptidového řetězce. Je tvořena střídáním sekundárních struktur. Podle tvaru a vlastností rozlišujeme strukturu globulární (albumin), která má tvar klubka a je rozpustná ve vodě, a fibrilární (myosin) vláknitou strukturu ve vodě nerozpustnou. Celá struktura je stabilizována kovalentními vazbami (např.: vazba S-S tzv. disulfidový můstek) v postranních řetězcích aminokyselin.
Kvartérní struktura
Řeší uspořádání podjednotek v proteinových aglomerátech, tvořících jednu funkční bílkovinu. Podjednotky jsou samostatné polypeptidické struktury, které jsou navzájem spojeny nekovalentními interakcemi. Kvartérní struktura též řeší prostorové uspořádání těchto podjednotek. Takovéto uspořádání vykazují jen složitější komplexy bílkovin, např. fibrily kolagenu, nebo lidské DNA polymerázy.
Rozdělení na podjednotky přináší mnohé evoluční výhody oproti existenci jednoho ohromného řetězce. Při výskytu poruchy ve stavbě stačí nahradit poškozenou podjednotku, což je podobné stavbě budov za použití prefabrikátů. Místo výstavby podjednotky může být navíc odlišné od místa jejího výskytu. Bílkoviny se skládají z podjednotek buď odlišných (heteromultimery) nebo shodných (homomultimery). Oblasti styku jednotlivých podjednotek jsou tvořeny slabými vazbami (nekovalentními interakcemi), především vodíkovými můstky nebo hydrofóbním efektem.
Symetrie
Bílkoviny většinou vykazují v oblasti spoje prvky symetrie. Nejjednodušší symetrií je cyklická symetrie. Označuje se Cn, kde n je počet protomerů uspořádaných v kruhu, který je středem symetrie. Jednotlivé protomery spolu svírají úhel 360°/n. Nejobvyklejší je C2 symetrie, kde jsou dvě podjednotky přímo proti sobě. Vyšší cyklické symetrie jsou poměrně vzácné.
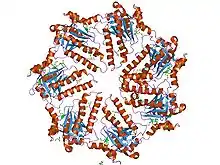
Složitější symetrie se nazývá diedrální a značí se Dn. Takové dimery jsou v podstatě tvořeny dvěma cyklicky symetrickými polovinami, jedna z nich leží pod a druhá nad rovinou symetrie. Střed symetrie se pak nachází v polovině spojnice středů symetrie obou cyklicky symetrických polovin. Polypeptidy s takovýmto uspořádáním jsou poměrně snadno disociovatelné na dva cyklicky symetrické oligomery. K další disociaci na protomery v přírodních podmínkách obvykle nedochází, protože k jejímu dosažení by už bylo zapotřebí značně drastických podmínek.
Funkce bílkovin
Bílkoviny jsou základem všech známých organismů, a proto v něm plní různé funkce.
- Stavební (kolagen, elastin, keratin)
- Transportní a skladovací (hemoglobin, transferin)
- Zajišťující pohyb (aktin, myosin)
- Katalytické, řídící a regulační (enzymy, hormony, receptory, …)
- Ochranné a obranné (imunoglobulin, fibrin, fibrinogen)
Proteinogenní aminokyseliny
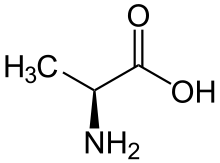
V proteinech se vyskytuje 20 kódovaných proteinogenních aminokyselin. K tomu jsou organismy schopné zainkorporovat do proteinů speciálními mechanismy další tři aminokyseliny. Jako tzv. 21. proteinogenní aminokyselina se označuje selenocystein (Sec). Tato aminokyselina nahrazuje cystein v lidském enzymu glutathionperoxidáze a v enzymech některých bakterií.[20] Jako 22. proteinogenní aminokyselina se označuje pyrolysin. U bakteriích, ale i plastidů a mitochondrií se vyskytuje N-formylmethionin, který je důležitý pro iniciaci translace a je první aminokyselinou zařazenou při tvorbě proteinu. U člověka se vyskytuje 21 proteinogenních aminokyselin (standardních 20 + selenocystein).
Ty aminokyseliny, které si organismus neumí sám syntetizovat a musí je přijímat, se označují jako esenciální aminokyseliny.

Důkaz bílkovin
Pro důkaz bílkovin se používají následující reakce:
Metabolismus bílkovin
Metabolismus bílkovin je souhrn různých biochemických procesů, při nichž jsou syntetizovány a rozkládány bílkoviny.
Syntéza bílkovin
Základní stavební částicí bílkovin jsou aminokyseliny, a tak je zřejmé, že se bez nich tzv. proteosyntéza neobejde. Některé aminokyseliny je schopné tělo vyrábět samo, jiné musí přijímat v potravě (k těmto tzv. esenciálním aminokyselinám patří u člověka v dospělosti 12 aminokyselin, v dětství 14[21]). Bílkoviny jsou ve většině případů kódovány v specifických úsecích v DNA organismů. Tyto úseky (tzv. geny) jsou přepisovány v procesu transkripce do mRNA a na ribozomu následně dochází k výrobě proteinů (translaci) za účasti této mRNA a jednotlivých aminokyselin napojených na specifické tRNA.
Vzácně některé relativně krátké polypeptidy nevznikají podle mRNA na ribozomu, ale v cytoplazmě syntézou pomocí složitých komplexů enzymů (NonRibosomal Peptide Synthetase), proto se nazývají neribozomální peptidy. Patří k nim některá polypeptidová antibiotika. Jejich produkce zůstane zachována i tehdy, zablokujeme-li ribozomální mašinérii. K těmto polypeptidům patří chloramfenikol a graminicin S. Mechanismus syntézy těchto antibiotik je trochu podobný syntéze mastných kyselin.[22]
Rozklad proteinů
Rozklad proteinů se také nazývá proteolýza. Bílkoviny se tráví na aminokyseliny, v krvi je určitá stálá hladina aminokyselin. Zdrojem aminokyselin jsou jednak bílkoviny z potravy, jednak opotřebované bílkoviny z tkání; malé množství aminokyselin vzniká při přeměně sacharidů. Aminokyseliny jsou potřebné: k syntéze stavebních bílkovin těla, k syntéze enzymů a hormonů, k syntéze plazmatických bílkovin, k přeměně na sacharidy. Část aminokyselin se odbourává na jednodušší látky a při tom se získává energie. Bílkoviny se neukládají do zásob.[zdroj?] Při katabolickém odbourání aminokyselin dochází nejdříve k jejich deaminaci. Aminové skupiny se odštěpují ve formě toxického amoniaku, který je v jaterních buňkách v tzv. ornitinovém cyklu přeměněn na močovinu, jež je krví zanesena do ledvin a vyloučena močí z těla. Uhlíkaté zbytky aminokyselin se začleňují do Krebsova cyklu, kde jsou dekarboxylovány a dehydrogenovány.
Pozn.: 1 molekula močoviny = 3 ATP = ornitinový cyklus.
Mezi enzymy štěpící bílkoviny patří např. pepsin (v žaludku), směs enzymů zvaná erepsin (v tenkém střevě) nebo také trypsin a chymotrypsin (produkované slinivkou břišní).[zdroj?] Mezi hormony řídící štěpení bílkovin patří glukokortikoidy, somatotropin, testosteron a insulin.[zdroj?]
Odkazy
Reference
- Mulder GJ (1838). "Sur la composition de quelques substances animales". Bulletin des Sciences Physiques et Naturelles en Néerlande: 104.
- Harold H (1951). "Origin of the Word 'Protein.'". Nature. 168 (4267): 244. Bibcode:1951Natur.168..244H. doi:10.1038/168244a0. PMID 14875059. S2CID 4271525.
- Perrett D (August 2007). "From 'protein' to the beginnings of clinical proteomics". Proteomics: Clinical Applications. 1 (8): 720–38. doi:10.1002/prca.200700525. PMID 21136729. S2CID 32843102.
- New Oxford Dictionary of English
- Reynolds JA, Tanford C (2003). Nature's Robots: A History of Proteins (Oxford Paperbacks). New York, New York: Oxford University Press. p. 15. ISBN 978-0-19-860694-9.
- Reynolds and Tanford (2003).
- Bischoff TL, Voit C (1860). Die Gesetze der Ernaehrung des Pflanzenfressers durch neue Untersuchungen festgestellt (in German). Leipzig, Heidelberg.
- Sumner JB (1926). "The isolation and crystallization of the enzyme urease. Preliminary paper" (PDF). Journal of Biological Chemistry. 69 (2): 435–41. doi:10.1016/S0021-9258(18)84560-4. Archived from the original on 2011-03-25. Retrieved 2011-01-16.
- Kauzmann W (May 1956). "Structural factors in protein denaturation". Journal of Cellular Physiology. 47 (Suppl 1): 113–31. doi:10.1002/jcp.1030470410. PMID 13332017.
- Kauzmann W (1959). "Some factors in the interpretation of protein denaturation". Advances in Protein Chemistry Volume 14. Advances in Protein Chemistry. Vol. 14. pp. 1–63. doi:10.1016/S0065-3233(08)60608-7. ISBN 978-0-12-034214-3. PMID 14404936.
- Kalman SM, Linderstrøm-Lang K, Ottesen M, Richards FM (February 1955). "Degradation of ribonuclease by subtilisin". Biochimica et Biophysica Acta. 16 (2): 297–99. doi:10.1016/0006-3002(55)90224-9. PMID 14363272.
- Sanger F (1949). "The terminal peptides of insulin". The Biochemical Journal. 45 (5): 563–74. doi:10.1042/bj0450563. PMC 1275055. PMID 15396627.
- Sanger F. (1958), Nobel lecture: The chemistry of insulin(PDF), Nobelprize.org, archived (PDF) from the original on 2013-01-05, retrieved 2016-02-09
- Stoddart, Charlotte (1 March 2022). "Structural biology: How proteins got their close-up". Knowable Magazine. doi:10.1146/knowable-022822-1. Retrieved 25 March 2022.
- Muirhead H, Perutz MF (August 1963). "Structure of hemoglobin. A three-dimensional fourier synthesis of reduced human hemoglobin at 5.5 Å resolution". Nature. 199 (4894): 633–38. Bibcode:1963Natur.199..633M. doi:10.1038/199633a0. PMID 14074546. S2CID 4257461.
- Kendrew JC, Bodo G, Dintzis HM, Parrish RG, Wyckoff H, Phillips DC (March 1958). "A three-dimensional model of the myoglobin molecule obtained by x-ray analysis". Nature. 181(4610): 662–66. Bibcode:1958Natur.181..662K. doi:10.1038/181662a0. PMID 13517261. S2CID 4162786.
- Zhou ZH (April 2008). "Towards atomic resolution structural determination by single-particle cryo-electron microscopy". Current Opinion in Structural Biology. 18 (2): 218–28. doi:10.1016/j.sbi.2008.03.004. PMC 2714865. PMID 18403197.
- Keskin O, Tuncbag N, Gursoy A (April 2008). "Characterization and prediction of protein interfaces to infer protein-protein interaction networks". Current Pharmaceutical Biotechnology. 9 (2): 67–76. doi:10.2174/138920108783955191. hdl:11511/32640. PMID 18393863.
- "RCSB Protein Data Bank". Archived from the originalon 2015-04-18. Retrieved 2017-01-19.
- KODÍČEK, M. selenocystein. From Biochemické pojmy : výkladový slovník [online]. Praha: VŠCHT Praha, 2007 [cit. 2008-10-21]. Dostupné online
- VODRÁŽKA, Zdeněk. Biochemie. Praha: Academia, 2007. ISBN 978-80-200-0600-4.
- VOET, Donald; VOET, Judith. Biochemie. 1.. vyd. Praha: Victoria Publishing, 1995. ISBN 80-85605-44-9.
Literatura
- Reisenauer R. et al.: CO JE CO? (1) Příručka pro každý den. Pressfoto - vydavatelství ČTK, Praha, 1982
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu bílkovina na Wikimedia Commons
Obrázky, zvuky či videa k tématu bílkovina na Wikimedia Commons  Slovníkové heslo bílkovina ve Wikislovníku
Slovníkové heslo bílkovina ve Wikislovníku- http://e-chembook.eu/bilkoviny