Hepatocyt
Hepatocyt, neboli jaterní buňka, je buňka tvořící základ jaterní tkáně, která je zodpovědná za většinu metabolických pochodů, která probíhají v játrech. Tvoří 78 % objemu jater a 60 % všech buněk tohoto orgánu.[1][2] V každém miligramu lidských jater se nachází asi 171×103 hepatocytů.[1] Je to epitelová buňka, pro kterou je typická těsná vazba s ostatními buňkami a přítomnost keratinů.
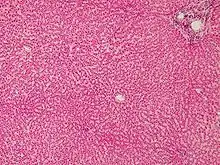
Vzhled
Hepatocyty jsou polyedrické, asi 30 µm velké[1] buňky se světlým, kulatým jádrem a jedním nebo dvěma výraznými jadérky.[3] Většinou mají pouze jedno jádro,[1][3] ale dvoujaderné hepatocyty jsou časté.[3]
Hepatocyty nenasedají na žádnou bazální membránu.[1] Jednou stranou směřují směrem ke krevní vlásečnici a boky se dotýkají ostatních hepatocytů. V místech, kde na sebe těsně naléhají jednotlivé hepatocyty, je jejich okraj hladký a jednotlivé buňky jsou spojené buněčnými spoji, desmozomy.[4] Výjimku tvoří místa, kde dva sousedící hepatocyty vytváří žlučovou kapiláru, kam je produkována žluč. Zde dva k sobě přiléhající hepatocyty vytvoří prostor vchlípením cytoplazmy a jeho utěsněním dalšími buněčnými spoji, kromě desmozomů také těsnými spoji (tight junction) a vodivými spoji (gap junction).[4] Cytoplazmatická membrána směřující dovnitř žlučové kapiláry se vychyluje v pravidelné mikroklky.[4]
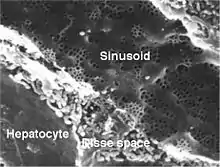
Strana směřující ke krevní vlásečnici je rovněž vybavena mikroklky, které jsou ale nepravidelné.[4] Mikroklky směřují do tzv. Disseho prostoru, což je 0,2-0,5 μm široký prostor mezi endotelovou buňkou vlásečnice, neboli sinusoidy, a hepatocytem.[5]
Organely hepatocytu a jejich funkce
Jádro hepatocytu je kulaté a tvoří 5–10 % objemu buňky.[6] Ve světelném mikroskopu se jeví světle, protože většina chromatinu je tzv. euchromatin, s aktivní transkipcí genů.[6] Asi 25 % hepatocytů má dvě jádra.[6] Vzácností není ani zvýšená ploidie jader, která je přímo úměrná velikosti jádra.[6] Můžou být tetraploidní, se čtyřmi chromozomovými sadami, a od dvaceti let věku člověka se objevují i oktoploidní jádra.[4] Velké množství chromozomových sad v jádrech hepatocytů je však považováno za prekancerózní změny.[4] Časté jsou jaderné inkluze tvořené glykogenovými zrny či tukovými kapénkami.[6]
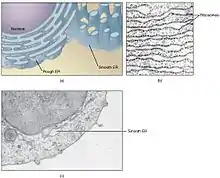
Bohatě rozvinuté je endoplazmatické retikulum. Je to soustava cisteren a kanálků tvořených membránou, která se napojuje na buněčné jádro. Tvoří 15–20 % objemu buňky[7] a jeho celkový povrch je 63000 μm2, což je asi 38× víc než povrch cytoplazmatické membrány celé buňky.[7] Při metabolickém stresu se dokáže ještě zvětšit.[7] 60 procent endoplazmatického retikula hepatocytu je tvořeno drsným endoplazmatickým retikulem, na jehož membráně jsou přítomné ribozomy.[7]
Zde jsou syntetizovány bílkoviny, a to především albumin, srážecí faktory a enzymy, jako je glukóza-6-fosfatáza.[8] V játrech jsou tvořeny všechny bílkoviny krevní plazmy kromě gama-globulinů, a jejich syntéza probíhá právě na drsném ER. Dále jsou zde tvořeny triacylglyceridy z volných masných kyselin a spojovány s proteiny za vzniku lipoproteinů, a tato organela se podílí i na syntéze glykogenu.[8]
Hladké endoplazmatické retikulum je místem, kde probíhá množství biochemických reakcí, na jeho membránách se nachází skupina cytochromů P450, kde dochází k oxidaci substrátů a detoxikaci cizorodých látek, je zde také syntetizován cholesterol, steroidy a žlučové kyseliny a probíhá zde degradace hemu.[8]
Soustava Golgiho aparátu slouží k transportu, přechovávání a úpravě látek syntetizovaných buňkou. V hepatocytu se váčky Goldiho aparátu nachází především v blízkosti žlučové kapiláry, podílí se zřejmě významně na produkci žluči.[8]
Mitochondrie hepatocytů jsou početné. Na kristách vnitřní membrány mitochondrií probíhají procesy, které zásobují hepatocyt energií, především pak oxidativní fosforylace.[9] Matrix mitochondrií je místem, kde se odehrává Krebsův cyklus, částečně i beta-oxidace mastných kyselin, část ornitinového cyklu, některé kroky syntézy steroidů a syntéza hemu.[9]
Peroxizomy se nachází především v blízkosti hladkého endoplazmatického retikula a glykogenových inkluzí.[8] Jejich funkce je spojena především s oxidací vyšších karboxylových kyselin a eikosanoidů a syntézou plazmalogenů a cholesterolu. Kromě toho obsahuje také enzymy, jako je glykolátoxidáza, kataláza, a oxidáza D-aminokyselin.[10]
Lyzozomy se rovněž nachází v blízkosti žlučové kapiláry. Obsahují proteolytické enzymy a slouží též jako zásobárna železa v podobě ferritinu, ukládají také měď, žlučové pigmenty a lipofuscin.[8]
V cytoplazmě hepatocytu jsou přítomné četné inkluze, především glykogenu,[3] dále lipidové kapénky a granula ferritinu.[8] Lipidové kapénky tvoří 0,3-2,1 % objemu buňky.[11]
Plazmatická membrána hepatocytů je vyztužena vláknitými bílkovinami, které tvoří podpůrný cytoskelet, udržující tvar buňky. Tuto funkci mají zvláště intermediární filamenta, u hepatocytů se jedná o cytokeratiny.[8][12] Cytokeratinová vlákna se táhnou od perinukleární oblasti k cytoplazmatické membráně, kde jsou ukotvena v pevných buněčných spojích, desmozomech. Udržují tak stabilní tvar buňky a celé tkáně. Lidské hepatocyty obsahují cytokeratiny typu 8 a 18.[12]
S desmozomy jsou asociována také mikrofilamenta, tvořená aktinem. Jsou kontraktilním elementem, v hepatocytu se vyskytují především pod cytoplazmatickou membránou v blízkosti žlučové kapiláry,[12] kterou tak vyztužují,[8] a pronikají také do mikroklků.[12] Zajišťují tak motilitu kapiláry a proudění žluči.[12][8]
Poslední složkou cytoskeletu hepatocytů jsou mikrotubuly, které slouží jako "koleje" k transportu organel a umožňuje také jaderné dělení tvorbou dělicího vřeténka.
Cytoplazmatická membrána je rozdělena na tři specializované oblasti podle toho, jestli směřuje do Dysseho prostoru, do žlučové kapiláry nebo k jiným hepatocytům. Tyto oblasti se liší jak vlastnostmi samotné membrány, jako je její fluidita, po enzymovou i receptorovou výbavu.[11]
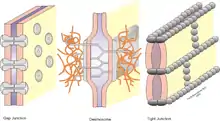
Část cytoplazmatické membrány, která tvoří žlučovou kapiláru, tvoří asi 15 % povrchu hepatocytu.[11] Transport látek na ní je jednosměrný, z hepatocytu směrem do lumina kapiláry.[11] V membráně jsou umístěné enzymy alkalická fosfomonoesteráza a ATP-áza. Ze všech stran je utěsněna těsnými spoji, které zabraňují úniku makromolekul do mezibuněčného prostoru mezi hepatocyty.[11] Na laterálních plochách hepatocytů jsou pravidelně rozmístěné desmozomy a vodivé spoje, kterými spolu komunikují sousedící buňky.[11] Na membráně přivrácené do Dysseho prostoru je možný oboustranný transport látek.[11]
Enzymatická výbava
Hepatocyty mají enzymatickou výbavu k průběhu většiny metabolických drah v organismu. Játra slouží jiným orgánům a tkáním, které tyto enzymy netvoří. Speciálními enzymy hepatocytů jsou:[13]
- Glukokináza, umožňuje rychle vychytávat glukózu z krve, je-li její koncentrace zvýšená (po jídle)
- Glukóza-6-fosfatáza, působí opačně, umožňuje uvolňovat glukózu do krve
- Glycerolkináza, umožňuje využití glycerolu buď při glykolýze, nebo pro syntézu triacylglycerolů a fosfoglycerolů
- Fosfoenolpyruvátkarboxykináza, řídící enzym metabolické dráhy syntézy glukózy, glukoneogeneze
- Fruktokináza, umožňuje vychytávat fruktózu z krve
- Argináza, poslední enzym v ornitinovém cyklu, ve kterém je tvořena močovina
- 3-hydroxy-3-methylglutaryl-CoA-syntáza, mitochondriální enzym umožňující vznik ketolátky acetacetátu
- 3-hydroxy-3-methylglutaryl-CoA-lyáza, mitochondriální enzym umožňující vznik ketolátky acetacetátu, při ketokegezi musí být přítomny oba enzymy, syntáza i lyáza
- 7-α-hydroxyláza, enzym umožňující vznik žlučových kyselin z cholesterolu
V játrech se pro užití v organismu zpracovávají volné mastné kyseliny, laktát, glycerol, fruktóza a aminokyseliny a nezastupitelná je role jater v udržování stálé koncentrace glukózy v krvi.
Uspořádání hepatocytů

Hepatocyty jsou v jaterní tkáni uspořádány do trámců, které se větví a zase se spojují v pruzích vedoucích od centrální žíly po okraj jaterního lalůčku, který má tvar šestibokého hranolu, s centrální žílou uprostřed. V místech, kde se dotýkají tři sousedící lalůčky, probíhá jaterním parenchymem větev jaterní tepny, portální žíly a žlučovod, toto uspořádání tvoří tzv. triádu. Trámce hepatocytů jsou obvykle jeden až dvě buňky tlusté, u novorozenců a dětí jsou však tvořeny dvěma nebo více buňkami.[5] Mezi centrální žílou a triádou se v jedné řadě nachází asi 24 hepatocytů.[5] Mezi trámci hepatocytů se proplétají sinusoidy.
Protože ne všechny hepatocyty se nachází stejně daleko od přívodní jaterní tepny, jsou nestejně zásobené kyslíkem a živinami. Hepatocyty se podle svého umístění vůči krevnímu zásobení liší i morfologicky a odlišná je i jejich enzymatická výbava a hlavní funkce, které vykonávají. Toto zohledňuje popis tzv. primárního jaterního acinu, který se dělí na tři zóny:
- 1. zóna, periportální, je nejlépe zásobená kyslíkem a živinami. Je tvořena hepatocyty, které se nacházejí nejblíž přívodní jaterní arterioly. Odehrává se v nich především glukoneogeneze, syntéza glutathionu a procesy vyžadující energii, jako je syntéza močoviny. Produkce žluči periportálními buňkami je závislá na žlučových kyselinách.[14]
- 2. zóna je zónou přechodnou
- 3. zóna, periacinární, je nejméně zásobená živinami a kyslíkem. Převládá zde aktivita cytochromu P-450 a detoxikace látek a glykolýza, tvoří se zde glutaminsyntetáza a sekrece žluči je nezávislá na žlučových kyselinách.[14]
Životní cyklus hepatocytů a jejich regenerace
Hepatocyty pokusných zvířat žijí asi 150 dní.[1] Staré či poškozené buňky zanikají apoptózou.[15] Ve zdravé jaterní tkáni vzniká stejně nových buněk, jako jich zaniká.[15] Hepatocyty se obnovují dělením: dle potřeby jsou schopny přejít z G0 fáze do G1 fáze. Za normálních okolností je mitotický index jaterní tkáně malý, jen asi 1:104–2,2×103,[6] ale rychlost obnovy jaterní tkáně může rychle stoupnout, například po odnětí části jaterní tkáně.[15]
Dojde-li k takovému poškození jaterní tkáně, že přežívající hepatocyty obnovu samy nemohou zvládnout, nastupují na jejich místo také kmenové buňky, které jsou v játrech připravené v blízkosti malých žlučových vývodů, mimo těchto kmenových buněk se v hepatocyt může diferencovat také kmenová buňka kostní dřeně.[15]
Odkazy
Reference
- Sherlock's Diseases of the Liver and Biliary System. Příprava vydání James S. Dooley, Anna Lok, Andrew K. Burroughs, Jenny Heathcot; autor kapitoly: Jay H. Lefkowitch. 12. vyd. [s.l.]: Wiley-Blackwell, 2011. 792 s. ISBN 978-1-4051-3489-7. Kapitola Anatomy and Function, s. 9. (anglicky)
- DANCYGIER, Henryk. Clinical Hepatology: Principles and Practice of Hepatobiliary Diseases: Volume 1. Berlín: Springer, 2009. 680 s. ISBN 978-3-540-93841-5. Kapitola Microscopic Anatomy, s. 16. (anglicky)
- BOWEN, R. Hepatic Histology: Hepatocytes [online]. Colorado State University, 23.6.1998 [cit. 2014-01-01]. Dostupné online. (anglicky)
- Sherlock's Diseases of the Liver and Biliary System. autor kapitoly: Jay H. Lefkowitch. Kapitola Anatomy and Function, s. 11. (anglicky)
- DANCYGIER, Henryk. Clinical Hepatology: Principles and Practice of Hepatobiliary Diseases: Volume 1. Kapitola Microscopic Anatomy, s. 17.
- DANCYGIER, Henryk. Clinical Hepatology: Principles and Practice of Hepatobiliary Diseases: Volume 1. Kapitola Microscopic Anatomy, s. 26.
- DANCYGIER, Henryk. Clinical Hepatology: Principles and Practice of Hepatobiliary Diseases: Volume 1. Kapitola Microscopic Anatomy, s. 27.
- Sherlock's Diseases of the Liver and Biliary System. autor kapitoly: Jay H. Lefkowitch. Kapitola Anatomy and Function, s. 12. (anglicky)
- DANCYGIER, Henryk. Clinical Hepatology: Principles and Practice of Hepatobiliary Diseases: Volume 1. Kapitola Microscopic Anatomy, s. 31.
- Chandoga J., Tomková M., Hlavatá A. PEROXIZÓMOVÉ DEDIÈNÉ OCHORENIA. S. 32–42. Bratislava Medical Journal [online]. [cit. 2014-01-02]. Roč. 1997, čís. 98, s. 32–42. Dostupné v archivu pořízeném dne 2007-06-06. (slovensky)
- DANCYGIER, Henryk. Clinical Hepatology: Principles and Practice of Hepatobiliary Diseases: Volume 1. Kapitola Microscopic Anatomy, s. 25.
- DANCYGIER, Henryk. Clinical Hepatology: Principles and Practice of Hepatobiliary Diseases: Volume 1. Kapitola Microscopic Anatomy, s. 32.
- MURRAY, K. Harperova biochemie. Praha: H & H, 2002. 872 s. ISBN 80-7319-013-3. Kapitola Vzájemná souvislost metabolismu a zajištění tkáňového paliva, s. 298. (česky)
- Sherlock's Diseases of the Liver and Biliary System. autor kapitoly: Jay H. Lefkowitch. Kapitola Anatomy and Function, s. 15.
- Sherlock's Diseases of the Liver and Biliary System. autor kapitoly: Jay H. Lefkowitch. Kapitola Anatomy and Function, s. 17.
Literatura
- Sherlock's Diseases of the Liver and Biliary System. Příprava vydání James S. Dooley, Anna Lok, Andrew K. Burroughs, Jenny Heathcot; autor kapitoly: Jay H. Lefkowitch. 12. vyd. [s.l.]: Wiley-Blackwell, 2011. 792 s. ISBN 978-1-4051-3489-7. (anglicky)
- DANCYGIER, Henryk. Clinical Hepatology: Principles and Practice of Hepatobiliary Diseases: Volume 1. Berlín: Springer, 2009. 680 s. ISBN 978-3-540-93841-5. (anglicky)
Externí odkazy
 Obrázky, zvuky či videa k tématu Hepatocyt na Wikimedia Commons
Obrázky, zvuky či videa k tématu Hepatocyt na Wikimedia Commons