Metody výzkumu protein-proteinových interakcí
Metody výzkumu protein-proteinových interakcí nám umožňují detekovat a hlouběji zkoumat interakce mezi rozmanitými proteiny. I přesto, že nukleotidová sekvence lidského genomu byla již zcela vyřešena, doposud nám nejsou známy funkce některých v ní zakódovaných proteinů. Funkci proteinu lze dobře zjistit, pokud se nám podaří odhalit jeho interakční partnery. Zároveň variabilita protein-proteinových interakcí, jejich regulace v čase a prostoru se podílejí na komplexitě celé řady dějů v organismech[1]. K účelu výzkumu protein-proteinových interakcí byla (a stále je) vyvíjena celá paleta metod. Každá z nich má své výhody i nevýhody a je vhodná pro určitý typ proteinu.
Základní rozdělení metod výzkumu protein-proteinových interakcí (PPI) je:
- metody, které umožňují interakci objevit
- metody, které umožňují zjištěnou interakci ověřit/potvrdit a lépe charakterizovat (např. Kd, kvantifikace, stechiometrie, struktura...)
V souvislosti s výzkumem PPI se užívá pojmu "bait" (návnada) pro protein, který studujeme a jehož interakční partneři nás zajímají. "Prey" (dravec) označuje ony interakční partnery. Cílem naší metody je zjednodušeně "nalákat dravce na návnadu, chytit ho a identifikovat".
Metody detekce PPI
Detekovat nové PPI umožňují zejména metody afinitní purifikace, ko-imunoprecipitace, kvasinkový dvouhybridový systém a podobné komplementační metody, proteinové čipy (array), blízkostí indukovaná ligace či značení[1].
Pro všechny tyto metody je typická nízká specificita, výsledkem je značné množství tzv. false-pozitiv výsledků[1]. Je proto třeba detekované interakce vždy ověřit nějakou specifičtější metodou. Proč ji nevyužijeme už na začátku? Je pro ni potřebné znát potenciální interakční patnery, které neznáme. Metody detekce PPI jsou primárním screeningem, který nám umožní vyloučit obrovské množství potenciálních interakčních partnerů a vytipuje užší skupinu, kterou můžeme dále analyzovat.
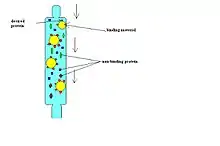
Afinitní purifikace (AP)
Tato metoda může být aplikována jak v in vivo, tak v in vitro podobě. Je relativně nenáročná, levná, nízkoprůchodná – tzn. hodí se pro analýzu jednotlivých proteinů, nikoliv na detekci celého interaktomu. Má nízkou specificitu i senzitivitu - detekovány jsou převážně stabilní, silné interakce. Senzitivitu je možné zvýšit při in vitro provedení[1].
In vitro varianta
Protein našeho zájmu (návnada) je připojen na pevnou matrici (agarózové/magnetické kuličky). Může být připojen kovalentně, či vysokoafinní nekovalentní vazbou (například přes protilátku). Následně aplikujeme směs buněčných proteinů na matrici s návnadou. Ty proteiny, které s návnadou interagují, se na ní zachytí. Zbytek neinteragujících proteinů odmyjeme. Dalším krokem je eluce proteinů, které se na naši návnadu chytily, obvykle pomocí roztoku s vysokou koncentrací solí, detergentem, či molekulou kompetující o vazbu návnady. Eluované proteiny je nakonec potřeba identifikovat - typicky pomocí hmotnostní spektrometrie[1].
Tato metoda má jeden značný problém – proteiny nám interagují v nefyziologických podmínkách. Výsledné interakce mohou být tedy pouze důsledkem denaturace proteinu během izolace, nebo se k sobě dostanou proteiny vyskytující se běžně ve zcela odlišných buněčných kompartmentech[2].
Zvýšit senzitivitu metody je možné prostřednictvím vysoké koncentrace proteinu našeho zájmu na matrici. V takovém případě je možné zachytit i relativně slabé a transientní interakce[1].
In vivo varianta
Cílem in vivo afinitní purifikace je izolovat skutečné, fyziologické interakční komplexy, ve kterých se vyskytuje protein našeho zájmu v buňce. Za tím účelem je typicky geneticky modifikován - připojíme k němu určitou značku, s jejíž pomocí ho budeme moct snadno izolovat včetně jeho interakčních partnerů (His tag, FLAG, Myc, HA, GFP, GST, PrA). Nebo ho nemodifikujeme a izolaci provádíme pomocí specifických protilátek. Proces purifikace je v principu stejný jako v in vitro případě, opět máme pevnou matrici, na které jsou připojeny specifické protilátky či jiné vysokoafinní molekuly. Ty nekovalentní vazbou zachytí náš protein společně s jeho interakčními partnery. Nespecifické interakce jsou odmyty a to co zůstane eluujeme a identifikujeme hmotnostní spektrometrií[2].
Hlavní rozdíl od in vitro metody je tedy v tom, odkud přichází protein našeho zájmu. V in vitro případě musí být nejprve purifikován a připojen na matrici. V in vivo případě je společně s ostatními proteiny součástí buněčného lyzátu a na matrici se váže později, rovnou ve svém komplexu interakčních partnerů.
Proteinové značky
Využití protilátek, které zachytí neznačené endogenní proteiny má své výhody - není potřeba nic klonovat, protein je exprimován ve fyziologickém množství a podobě. Jen malé množství dostupných protilátek je ale vskutku specifické[2].
Z toho důvodu je častější využití proteinových značek, ke kterým existují skvělé protilátky či jiní vysokoafinní partneři, zaběhnuté protokoly. Nezáleží pak tolik na charakteru našeho proteinu a s vysokou pravděpodobností ho bez problémů izolujeme. Slabina tohoto přístupu je exprese fúzního proteinu (náš protein-značka). Značka může měnit vlastnosti našeho proteinu a fúzní protein je exprimován z nefyziologického promotoru. V důsledku je ho v buňce víc, než by mělo a to může vyústit v nefyziologické interakce. Při použití tohoto přístupu je tedy třeba ověřit, že fúzní protein se chová zcela fyziologicky a zajistit jeho nízkou expresi[2].
Proteinové značky mohou být krátké aminokyselinové úseky - FLAG, HA, Myc, nebo se může jednat o celé proteiny - GFP, GST. Krátké značky obvykle nijak nevadí funkci proteinu, lze jich umístit i víc za sebou a zvýšit tak účinnost izolace. GFP je sice velkou značkou a vadit funkci proteinu může, ale přináší nám možnost značený protein pozorovat in vivo v buňkách díky fluorescenci[2].
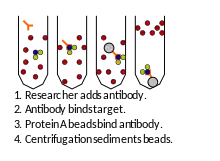
Ko-imunoprecipitace (co-IP)
Metoda je hodně podobná afinitní purifikaci. Do buněčného lyzátu přidáme protilátku specifickou k proteinu našeho zájmu. Následně přidáme sekundární protilátku či protein A připojené na agarózu. Náš protein společně se svými interakčními partnery se zachytí na agaróze a vzniká imunoprecipitát. Vzorek promyjeme, imunoprecipitáty lze snadno oddělit centrifugací[3].
Imunoprecipitát můžeme nakonec analyzovat pomocí hmotnostní spektrometrie[1].
Kvasinkový dvouhybridový systém
Více na samostatné stránce Kvasinkový dvouhybridový systém.
Jedná se o náročnější, senzitivní, genetickou, in vivo metodu. Detekujeme expresi určitého kvasinkového markeru (např. β galaktosidázy), která je spouštěna funkčním transkripčním faktorem. Transkripční faktory mají dvě důležité domény, bez nichž nemohou fungovat - transaktivační a DNA-vazebnou. DNA-vazebná doména je fúzována s proteinem našeho zájmu, transaktivační doména s potenciálním interakčním partnerem. Pouze pokud proteiny interagují, domény se dostanou k sobě a spouští expresi markeru, kterou detekujeme. Existují velké knihovny expresních vektorů s fúzemi rozmanitých proteinů k doménám transkripčních faktorů a lze proto simultánně testovat spoustu potenciálních proteinových interakcí. Metoda je vhodná i pro analýzu komplexních interaktomů[4][1].
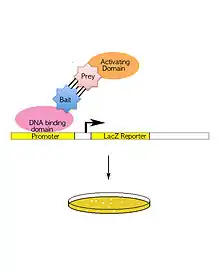
Nevýhod tohoto systému je několik. K interakci proteinových partnerů musí dojít v jádře, aby transkripční faktor fungoval. Metodu proto nelze použít pro proteiny membránové a vyskytující se v jiných buněčných kompatmentech. Kvasinky neumí vytvářet posttranslační modifikace stejné jako savčí buňky - pokud je tedy interakce založena na savčí modifikaci, v kvasince k interakci nedojde. Navíc fúze přídatné domény k proteinu může pozměnit jeho vlastnosti[4]. Typická je také velmi nízká specifita, vedoucí ke vzniku velkého množství false-pozitiv výsledků[1].
V důsledku slabin tradiční metody bylo odvozeno velké množství vylepšených přístupů, například[4]:
- Rekrutace proteinu Sos (son of sevenles) / Ras
- vhodné pro cytoplazmatické proteiny
- G proteinový fúzní systém
- vhodné pro interakci mezi membránovým (návnada) a cytoplazmatickým proteinem
- Savčí dvouhybridový systém
- fyziologické podmínky, savčí modifikace, folding
- A další...
Proteinové čipy
Jedná se relativně drahou, složitou, vysoko-průchodnou in vitro metodu. Umožňuje analyzovat opravdu velké množství proteinových interakcí zároveň. Fungují analogicky jako DNA čipy. Kromě studia interakcí s proteiny je lze použít také k detekci interakcí s oligonukleotidy[1].
Proteinové čipy se skládají z pevné matrice, na níž jsou ukotveny stovky až tisíce různých proteinů. Na jednom čipu se tak může nacházet celý proteom daného organismu. Nejnáročnější je tvorba čipu. Jednotlivé proteiny je potřeba vypurifikovat z buněk, nebo je in vitro syntetizovat. Pomocí in vitro syntézy je možné přepsat celou cDNA knihovnu do podoby čipu. Vazba interakčního partnera na čip je detekována typicky fluorescenčně[4][1].
Slabinou čipů je, že se jedná o in vitro proces. Imobilizace proteinů na matrici může ovlivnit jejich strukturu a interakční vlastnosti. Navíc pokud jsou proteiny syntetizovány in vitro, nebo exprimovány v bakteriálním systému, nebudou posttranslačně modifikovány.[4]
Blízkostí indukovaná ligace
Tato metoda nám umožňuje kovalentně provázat proteinový komplex in vivo, v buňce. Díky tomu jsme schopni zachytit i proteiny interagující slabě, či pouze dočasně s proteinem našeho zájmu. Pevně svázaný komplex pak můžeme izolovat třeba pomocí afinitní purifikace a analyzovat na hmotnostním spektrometru. Díky kovalentnímu provázání nám při izolaci "neodpadne" žádný z interagujících proteinů a my tak můžeme intenzivně promývat a zbavit se pozadí[4].
Ke kovalentnímu propojení se používá obvykle bifunkčních činidel, kdy dvě reaktivní funkční skupiny jsou odděleny řetízkem (linkerem) určité délky. Délka řetízku určuje maximální vzdálenost, ve které je možné proteiny propojit. Reaktivní skupiny mohou být různého typu a cílit tak na konkrétní aminokyseliny proteinu - typicky jsou propojovány amino- a sulfhydrylové skupiny. Některá činidla jsou UV-aktivovatelná a ligaci je tak možné spustit v konkrétní čas. Jiná prochází samovolně přes membrány a buňky není třeba permeabilizovat[4].
Provazování proteinů, které jsou v blízkosti proteinu našeho zájmu, s sebou nese nebezpečí propojení pouze náhodně "kolemjdoucích" proteinů[4].
Blízkostí indukované značení
Podobně jako předchozí metoda je blízkostí indukované značení zacíleno na zachycení všech interakcí našeho proteinu, včetně slabých a transientních in vivo. Je založena na fúzi proteinu našeho zájmu k enzymu, který připojí afinitní značku na proteiny v jeho bezprostředním okolí. Prostřednictvím značky je následně snadné označené interakční partnery vyizolovat a analyzovat na hmotnostním spektrometru. Tuto metodu lze dobře použít i pro membránové proteiny, které lze jen těžko solubilizovat. Nevýhodou je opět značení všech proteinů do určité vzdálenosti, tedy ne nutně pouze těch, které s naším proteinem opravdu interagují[5].
Jako afinitní značka je užíván zejména biotin. Biotinylace je v savčích buňkách vzácnou modifikací, navíc afinita biotinu k avidinu umožňuje snadnou izolaci. Některé enzymy vyloženě katalyzují připojení biotinu - tzv. biotin-ligázy, jiné tvoří reaktivní biotinové molekuly, které se následně ochotně vážou na aminokyseliny ve svém okolí. Dvěma nejznámějšími metodami jsou BioID (biotin-ligáza) a APEX (askorbát-peroxidáza)[5].
Další metody pro analýzu protein-proteinových interakcí
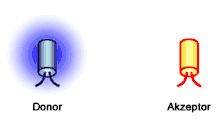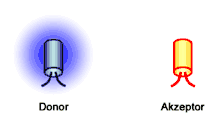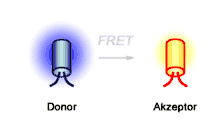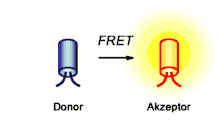
V okamžiku, kdy známe dva potenciální interakční partnery, nabízí se nám nepřeberné množství metod, jimiž je můžeme dále studovat a ověřovat.
Skvělou metodou je v tomto směru FRET(Försterův rezonanční přenos energie), či FCS(fluorescenční korelační spektroskopie), které nám umožňují pozorovat proteinový komplex pomocí fluorescence in vivo v buňkách, v reálném čase. FRET navíc umožňuje určení například KD komplexu[4][1].
In vitro metodami jsou například NMR, rentgenová krystalografie, CD (cirkulární dichroismus), SPR(povrchová plazmonová rezonance) a další. Mohou nám poskytnout velmi detailní pohled na interakční rozhraní partnerů[4][1].
Reference
- SYAFRIZAYANTI, null; BETZEN, Christian; HOHEISEL, Jörg D. Methods for analyzing and quantifying protein-protein interaction. Expert Review of Proteomics. February 2014, roč. 11, čís. 1, s. 107–120. PMID: 24393018. Dostupné online [cit. 2018-02-07]. ISSN 1744-8387. DOI 10.1586/14789450.2014.875857. PMID 24393018.
- MITEVA, Yana V.; BUDAYEVA, Hanna G.; CRISTEA, Ileana M. Proteomics-Based Methods for Discovery, Quantification, and Validation of Protein–Protein Interactions. Analytical Chemistry. 2013-01-15, roč. 85, čís. 2, s. 749–768. Dostupné online [cit. 2018-02-07]. ISSN 0003-2700. DOI 10.1021/ac3033257.
- VAN DER GEER, Peter. Analysis of protein-protein interactions by coimmunoprecipitation. Methods in Enzymology. 2014, roč. 541, s. 35–47. PMID: 24674061. Dostupné online [cit. 2018-02-07]. ISSN 1557-7988. DOI 10.1016/B978-0-12-420119-4.00004-5. PMID 24674061.
- ZHOU, Mi; LI, Qing; WANG, Renxiao. Current Experimental Methods for Characterizing Protein-Protein Interactions. ChemMedChem. 2016-04-19, roč. 11, čís. 8, s. 738–756. PMID: 26864455. Dostupné online [cit. 2018-02-07]. ISSN 1860-7187. DOI 10.1002/cmdc.201500495. PMID 26864455.
- KIM, Dae In; ROUX, Kyle J. Filling the Void: Proximity-Based Labeling of Proteins in Living Cells. Trends in Cell Biology. 11 2016, roč. 26, čís. 11, s. 804–817. PMID: 27667171 PMCID: PMC5077660. Dostupné online [cit. 2018-02-07]. ISSN 1879-3088. DOI 10.1016/j.tcb.2016.09.004. PMID 27667171.