Průduch
Průduch (odborně též stoma z řec. στόμα–ústa, množ. č. stomata) je struktura vyskytující se především na listech většiny vyšších rostlin, která slouží ke kontrolované výměně plynů (především CO2 a O2) mezi rostlinou a okolím a také k odevzdávání vody do ovzduší. Průduchy se tak zařazují z hlediska funkce mezi tzv. provětrávací pletiva.
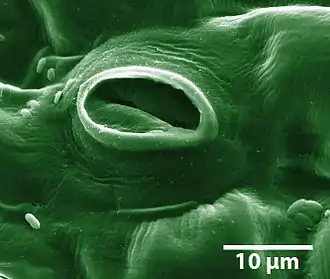
(kolorovaná mikrofotografie z rastrovacího elektronového mikroskopu)
Průduch dosahuje rozměrů v setinách milimetru. Skládá se především ze dvou specializovaných pokožkových buněk, označovaných jako svěrací buňky, mezi nimiž je tzv. průduchová štěrbina. Někdy se kolem svěracích buněk tvoří ještě buňky vedlejší, které se účastní otevírání průduchů. Stavba i počet průduchů jsou u rostlin různé, závislé na přizpůsobení abiotickým faktorům prostředí (hlavně vlhkost půdy a ovzduší).
Mechanismus otevírání a zavírání průduchů je založen na regulovaném příjmu a výdeji vody. Pokud svěrací buňky nasávají vodu, mění se jejich tvar a průduch se otevírá. V opačném případě se průduchová štěrbina uzavírá. V příjmu vody hrají roli především kationty draslíku, které podle potřeby zvyšují a snižují koncentraci uvnitř svěracích buněk. Z vnějších faktorů může uzavření průduchů způsobit hlavně vadnutí rostliny (nedostatek vody) a vysoká teplota; nízká koncentrace oxidu uhličitého v pletivech naopak svěrací buňky otevírá. Rostliny žijící v suchém prostředí si vyvinuly množství adaptací, jak nepřízním takového podnebí odolat.
Historie výzkumu

Ačkoliv si vědci všimli vypařování vody z listů již dříve, prvním, kdo pozoroval průduchy buněk, byl italský přírodovědec Marcello Malpighi, který tento objev publikoval ve svém díle Anatome plantarum (česky Rostlinná anatomie) v roce 1675. Ten však paradoxně nebyl schopen přisoudit průduchům jejich pravou funkci. Naopak jeho současník Nehemiah Grew již hypotetizoval, že jím pozorované průduchy slouží k ventilaci vnitřního prostředí rostliny a přirovnal průduchy k vzdušnicím hmyzu. Velký rozvoj znalostí o průduších nastal v 19. století, kdy také bylo slovo „stoma“ poprvé použito švýcarským botanikem de Candollem (v roce 1827). Průduchy se v té době zabývali například Hugo von Mohl (objevil základní princip otevírání průduchů), či anatomové jako Simon Schwendener (klasifikoval průduchy podle jejich stavby). Dodnes se některé záležitosti ohledně mechanismu funkce průduchů intenzívně zkoumají, a to zejména na rostlinách křížatka obecná (Commelina communis), bob obecný (Vicia faba) a kukuřice setá (Zea mays).[1]
Funkce
Rostlina je pomocí průduchů schopná regulovat výměnu plynů otevíráním a zavíráním skuliny průduchu, a to tak, aby měla maximální přísun oxidu uhličitého při únosných ztrátách vody. Rovnováha mezi příjmem CO2 pro fotosyntézu a výdejem vody pro pohánění transpiračního proudu rostlinou se označuje jako fotosynteticko–transpirační kompromis. Průduchy jsou velmi efektivní výměník plynů: projde jimi každoročně 3×1015 kilogramů uhlíku a celkový objem vody v atmosféře se jimi recykluje každý půlrok.[2] Tvoří tak významný prvek v koloběhu uhlíku i v planetárním oběhu vody. U většiny vyšších rostlin odchází voda převážně právě průduchy. Výdej vody přes pokožku (kutikulární transpirace) oproti tomu činí nanejvýš 30 % (obvykle mnohem méně a u sukulentů pouze 1–2 %), výdej vody přes lenticely u dřevin je zcela zanedbatelný.[3]
Průduchy však mají zřejmě i jiné, donedávna netušené funkce. Například ohýbání vzrostného vrcholu za světlem tzv. fototropismus) je zřejmě reakcí na skutečnost, že se voda vypařuje intenzivněji na osluněné straně rostliny, než na straně zastíněné.[1][4] Na druhou stranu mohou průduchy pro rostlinu představovat výrazné riziko, protože jsou „otevřenými dveřmi“ pro vstup patogenů do těla rostlin. V určitých případech jsou rostliny schopné patogenní bakterie detekovat a následně průduchy zavřít. To však určité bakterie obchází tím, že vylučují do okolí látku koronatin, která průduchy otevírá násilím.[5]
Evoluce
Průduchy se objevily již u prvních suchozemských rostlin, a tak se zdá, že jsou pro přežití fotosyntetizujících rostlin na souši zcela zásadní. Byly klíčovou adaptací v dobách, kdy rostliny dobývaly souš. Vznik průduchů se datuje do doby před přibližně 430–400 miliony lety[2][6] a objeveny byly již u fosílií silurské rostliny Cooksonia.[1] Z vyšších rostlin chybí zejména u játrovek, tří rodů hlevíků a některých bazálních (primitivních) mechů.[7] U mechorostů jsou obvykle průduchy přítomné na sporofytu (tzn. na tobolce), výjimečně i na gametofytu.[1]
Nejjednodušší vysvětlení evoluce průduchů spočívá v tom, že se vyvinuly u předka všech vyšších rostlin, načež u vyjmenovaných mechorostů druhotně zcela vymizely. Existují však i odlišné hypotézy. Je například možné, že se průduchy vyvinuly u různých rostlin nezávisle.[7] Druhotně vymizely průduchy také u plavuně druhu Stylites andicola, která pohlcuje oxid uhličitý pomocí svých kořenů,[1]) a u jistých nezelených druhů krytosemenných rostlin, jako je hlístník hnízdák (Neottia nidus-avis) či hnilák (Monotropa). Dále obvykle nemají průduchy ani vodní rostliny, vyjma listů plovoucích na hladině (leknín, Nymphaea).[3]
Stavba
Velikost průduchů (délka) obvykle kolísá mezi 0,01–0,06 milimetru (větší bývají průduchy polyploidních rostlin a zastíněných listů). Největší průduchy byly zjištěny u vyhynulých rostlin Zosterophyllum, které dosahovaly délky až 0,12 mm (120 µm).[1] Průduchy se skládají ze dvou buněk svěracích (cellulae claudentes), mezi nimiž je tzv. průduchová štěrbina (porus stomatalis). Svěrací buňky jsou obvykle v určitých částech ztlustlé, což podmiňuje mechanismus jejich funkce. Také plazmodezmy (kanálky ve stěně buněk) jsou u svěracích buněk nefunkční, což napomáhá udržovat nitrobuněčný tlak na požadované úrovni. Typickým rysem svěracích buněk je skutečnost, že obsahují chloroplasty, a proto mají zelenou barvu. Další uzpůsobení stavby průduchu obvykle úzce souvisí s prostředím, kde rostliny žijí.[3]
Vývoj průduchu
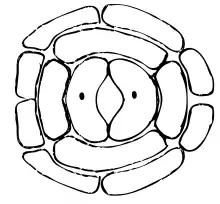
Průduch vzniká v rané fázi vývoje listu z části tuniky známé jako meristemoid. Buňky meristemoidu prochází asymetrickým dělením (inadekvátní mitóza) na jednu velkou epiteliální a jednu malou mateřskou buňku svěracích buněk. Tato mateřská buňka se následně opět rozdělí na dvě buňky svěrací. Mezi nimi vznikne (tzv. schizogenně, tedy rozpuštěním přepážky mezi buňkami a následným oddělením buněk od sebe) průduchová štěrbina, přes niž probíhá výměna plynů. Samotná mateřská buňka se však může rozdělit dvěma způsoby, na jejichž základě rozlišujeme:[3]
- průduchy haplocheilické – mateřská buňka se přímo rozdělí na dvě svěrací; například u jehličnanů (Pinophyta), cykasů (Cycadophyta), lilií (Lilium), kosatců (Iris) atp.;
- průduchy syndetocheilické – mateřská buňka se dělí několikrát, ve vzniklé populaci buněk se jedna prostřední rozdělí na vlastní svěrací buňky, zatímco okolní buňky se mění na buňky vedlejší; tyto vedlejší buňky se liší od typických pokožkových buněk a účastní se mechanismu otevírání a zavírání průduchů.
Podobné je dělení průduchů na základě počtu, uspořádání a tvaru pokožkových buněk kolem průduchů:
- průduchy izocytické (anomocytické) – buňky obklopující průduchy vypadají stejně jako běžné epidermální buňky; vývoj takových průduchů je nejčastěji haplocheilický (tzn. bez vedlejších buněk);
- průduchy anizocytické – buňky kolem průduchů se liší od běžných pokožkových buněk; obvykle se vyvíjí syndetocheilickým způsobem, tzn. buňky kolem svěracích buněk fungují jako pomocné. Popsáno bylo více než třicet subtypů anizocytických průduchů, které se navzájem liší tvarem vedlejších buněk (viz[8]).
Uzpůsobení k podmínkám prostředí

Mnohá uzpůsobení stavby průduchu jsou v silné korelaci s prostředím, kde dané rostliny žijí. Významná je z tohoto hlediska poloha průduchů vzhledem k rovině listu:[3]
- emerzní – nad úrovní listu (vyčnívají), typické pro vlhkomilné rostliny;
- submerzní (kryptoporní) – pod úrovní listu (ponořené), typické pro suchomilné rostliny (zamezuje nadměrným ztrátám vody);
- faneroporní – v rovině listu, nejčastější typ.[3]
U suchomilných rostlin bývá navíc na vnější straně průduchu vrstva vosku (jehličnany, trávy, sukulenty), nebo jsou navíc průduchy obklopeny trichomy (oleandr, Nerium). Bývá u nich také menší hustota průduchů na mm2. Některé rostliny si osvojily také jiné režimy otevírání průduchů, napomáhá tomu metabolicky zejména tzv. CAM cyklus.[3] Jiné jednoduše zavírají průduchy např. za horkého poledne.[9]
Tvar svěracích buněk
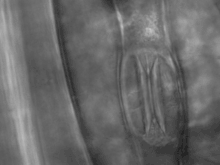
(světelný mikroskop)
Svěrací buňky jsou obvykle ledvinitého nebo piškotovitého tvaru, ale bylo popsáno množství odchylných druhů průduchů právě na základě jejich anatomie a mechanismu otevírání a zavírání:[3]
 | Typ Amaryllis (podle rostlin rodu Amaryllis) Svěrací buňky jsou ledvinité a nestejně ztlustlé. Vnitřní stěny (obrácené do štěrbiny) jsou totiž vyztužené dvěma podélnými lištami, naopak strana hřbetní (odvrácená od štěrbiny) je pružná. Když klesne turgor, buňky se napřimují a štěrbina se uzavírá. Jedná se o průduch typický pro jednoděložné rostliny, méně často je i u dvouděložných. |
 |
Typ Helleborus (podle rostlin rodu čemeřice, latinsky Helleborus) |
 |
Typ Gramineae (podle skupiny Gramineae, tzn. česky „trávy“) |
 |
Pteridofytní typ (podle skupiny Pteridophyta, tzn. česky kapraďorosty) |
 |
Gymnospermní typ (podle skupiny Gymnospermae, tedy česky nahosemenné) |
Umístění a počty průduchů

Průduchy se neomezují jen na povrch listů, ale je možné je najít také na některých částech květu (kališní a korunní lístky, plodolisty i tyčinky), na některých dalších částech květu trav a na povrchu nezralých plodů kdouloně (Cydonia), jablka (Malus domestica), banánu (Musa) či rajčete (Solanum lycopersicum), na vnitřní i vnější straně lusků i na samotných semenech hrachu. Navíc se nacházejí i na orgánech laiky považovaných za listy, jako jsou listeny a úponky. Výjimečně byly průduchy objeveny i na primárních kořenech, jako například u hrachu setého (Pisum sativum), nebo na hlízách, například u bramboru (Solanum tuberosum).[1]
Významné jsou však především průduchy na listech. Na 1 mm2 listu bývá obvykle 50–300 průduchů,[3] méně jich bývá u rostlin suchomilných[10] a pravděpodobně i u prehistorických rostlin, které žily v prostředí s vyšší koncentrací oxidu uhličitého.[2] Vyšší hustota průduchů je obvyklá u stromů.[1] Nejčastěji jsou rozloženy náhodně a přibližně rovnoměrně, ačkoliv u některých kapradin tvoří dvojice (kapraď samec, Dryopteris filix-mas) a u mnohých trav a jehličnanů jsou průduchy seřazeny v řadách za sebou. Obvykle jsou průduchy vyvinuty na spodní straně listů, ale zejména u některých jednoděložných jsou rozmístěny na obou stranách stejnoměrně.[10] Podle umístění průduchů se rozlišuje několik základních typů listů:[3]
- hypostomatické – nejčastější, průduchy převažují na spodní straně listu (spodní epidermis);
- amfistomatické – průduchy jsou rozloženy na spodní a horní pokožce přibližně ve stejném počtu; například kosatec (Iris) či většina lipnicovitých (Poaceae);
- epistomatické – průduchy jsou pouze na svrchní straně listu (svrchní epidermis); například listy leknínu (Nymphaea) a stulíku (Nuphar) plovoucí na hladině, dále traviny jako kostřava (Festuca), strdivka (Melica) a válečka (Brachypodium).
Uvádějí se například tyto poměry počtu průduchů na straně horní ku počtu průduchů na straně spodní: třešeň 0 : 253, jedle bělokorá 0 : 98, pšenice setá 50 : 40, rozchodník ostrý 21 : 14, borovice lesní 74 : 84, leknín bílý 460 : 0 (tyto poměry jsou spíše čísla udávající počet průduchů na 1 mm2, nula tedy znamená, že na dané straně nejsou průduchy vůbec).[10][11]
Mechanismus otevírání a zavírání
Většina rostlin je schopná regulovat velikost průduchové štěrbiny (výjimkou jsou vrby (Salix), jež tuto schopnost ztratily a bez vody rychle vadnou[3]). Mechanismus otevírání a zavírání průduchu je založen na změnách nitrobuněčného tlaku (turgoru) ve svěracích buňkách. Ty jsou vyztužené celulózovými fibrilami jen v jednom směru, a tak jsou poměrně pružné. Určité části stěny jsou však obvykle mnohem ohebnější než jiné části, a tak se změnou nitrobuněčného tlaku mění tvar buňky určitým požadovaným způsobem.[3] Celý mechanismus otevírání a zavírání průduchů je založen na regulovaném příjmu a výdeji vody ze svěracích buněk (případně také buněk vedlejších). Pokud svěrací buňky nasávají vodu, turgor se zvyšuje, mění se tvar svěracích buněk a průduch se otevírá. Při výdeji vody se naopak průduch zavírá. Rychlý příjem a výdej vody usnadňují akvaporinové kanály v cytoplazmatické membráně.
Velikost otevření průduchu závisí na mnoha vnějších faktorech prostředí. Patří k nim zejména světlo, teplota vzduchu a relativní vzdušná vlhkost. Tyto faktory ovlivňují celou řadu vnitřních faktorů, jako je koncentrace některých iontů uvnitř svěracích buněk (zejména draslíku) a produkce regulačních látek (kyseliny abscisové), stejně jako obsah oxidu uhličitého, pH v cytoplazmě a koncentrace škrobu či sacharózy ve svěracích buňkách.[3]
Ve dne se u většiny rostlin průduchy otevírají, aby mohly rostliny fotosyntetizovat. (Zcela naopak tomu je však u tzv. CAM rostlin, které žijí v suchých a horkých oblastech. Ty mají ve dne průduchy zavřené, aby neztrácely přebytečnou vodu, a v noci průduchy otevřou a zabudovávají oxid uhličitý do malátu.[3]) Co se týče vlivu slunečního záření na otevřenost průduchů, zdá se, že největší vliv má modrá složka spektra. Ta působí na receptor, jímž je v tomto případě rostlinný karotenoid jménem zeaxantin.[3] Ten ovlivňuje koncentraci draslíkových iontů v buňce. Ve dne jsou protonovými pumpami ionty K+ čerpány dovnitř buňky, ta je pak koncentrovanější (hypotonická, snižuje se její vodní potenciál[12]) a nasává (v procesu osmózy) vodu, čímž se průduch otevírá. Naopak po západu slunce draslíkové ionty z buněk pasivně (bez spotřeby energie[12]) vytékají a buňky ztrácí vodu (platí, že „voda následuje draslíkové ionty“). Možné také je, že světlo umožňuje v chloroplastech svěracích buněk výrobu ATP, který je nezbytný pro funkci protonových pump pro draslík, čímž by také mohlo být regulováno otevírání průduchů.[12]
Otevírání průduchů navozuje také nízká koncentrace oxidu uhličitého v houbovém parenchymu uvnitř listu, která signalizuje, že je třeba dodat do pletiv čerstvý vzduch. Rovněž se zdá, že v pravidelném každodenním otevírání průduchů hrají roli i biologické rytmy (tzv. vnitřní hodiny), které určitým způsobem zaznamenávají denní dobu. Průduchy může uzavřít prostý nedostatek vody ve svěracích buňkách, který signalizuje vadnutí rostliny. Vadnutí způsobuje navíc tvorbu kyseliny abscisové, která rovněž průduchy uzavírá. Také vysoké teploty, které znamenají rovněž hrozbu uschnutí, jsou schopné vyvolat zavření průduchů např. během poledního žáru.[12]
Průduch v kultuře
Slovo průduch se vyskytuje ve hře divadla Járy Cimrmana nazvané Posel z Liptákova. Ze hry však není jasné, zda se jedná o průduch rostlinný.[13]
Odkazy
Reference
- WILLMER, Colin Michael; FRICKER, Mark. Stomata. [s.l.]: Springer, 1996. Dostupné online. ISBN 0412574306.
- Keiko U. Torii. Stomatal development [online]. 2006. Dostupné online.
- VINTER, Vladimír. Rostliny pod mikroskopem; základy anatomie cévnatých rostlin. 2. vyd. Olomouc: Univerzita Palackého v Olomouci, 2009. ISBN 978-80-244-1972-5.
- MCINTYRE, G.I. Functional Plant Biol.. 1994-01-01, roč. 21, čís. 3, s. 359–375. Dostupné online.
- Maeli Melotto, William Underwood, Jessica Koczan, Kinya Nomura, Sheng Yang He. Plant Stomata Function in Innate Immunity against Bacterial Invasion. Cell. September 2006, roč. 126, s. 969–980. DOI 10.1016.
- WOODWARD, F. Do plants really need stomata?. J. Exp. Bot.. 1998-03-01, roč. 49, čís. suppl_1, s. 471–480. Dostupné online. DOI 10.1093/jexbot/49.suppl_1.471.[nedostupný zdroj]
- RENZAGLIA, K S, Duff RJT, D L Nickrent, D J Garbary. Vegetative and reproductive innovations of early land plants: implications for a unified phylogeny. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 2000-06-29, roč. 355, čís. 1398, s. 769–793. Dostupné online. ISSN 0962-8436. DOI 10.1098/rstb.2000.0615.
- Manual of Leaf Architecture [online]. Smithsonian Institution; Leaf Architecture Working Group, 1999 [cit. 2009-05-12]. Dostupné v archivu pořízeném dne 2009-05-10. ISBN 0-9677554-0-9.
- SLAVÍKOVÁ, Jiřina. Ekologie rostlin. Praha: Státní pedagogické nakladatelství, 1986. S. 366.
- ČERNOHORSKÝ, Zdeněk. Základy rostlinné morfologie. Praha: SPN, 1967.
- NĚMEC, Bohumil. Nauka o buňce, anatomie rostlin. Praha: Aventinum, 1930.
- CAMPBELL, Neil A.; REECE, Jane B. Biologie. Praha: Computer press, 2006. S. 1332.
- https://www.youtube.com/watch?v=YQa1R8xqnDo
Literatura
- (česky) VINTER, Vladimír. Rostliny pod mikroskopem; základy anatomie cévnatých rostlin. 2. vyd. Olomouc: Univerzita Palackého v Olomouci, 2009. ISBN 978-80-244-1972-5.
- (anglicky) WILLMER, Colin Michael; FRICKER, Mark. Stomata. [s.l.]: Chapman & Hall, 1995. Dostupné online. ISBN 0412574306.
- (anglicky) ZEIGER, Eduardo, G. D. Farquhar, I. R. Cowan. Stomatal function. Stanford: Stanford University Press, 1987. Dostupné online. ISBN 0804713472.
Externí odkazy
 Obrázky, zvuky či videa k tématu Průduch na Wikimedia Commons
Obrázky, zvuky či videa k tématu Průduch na Wikimedia Commons  Slovníkové heslo průduch ve Wikislovníku
Slovníkové heslo průduch ve Wikislovníku- Anatomie rostlin – stomata – na Katedře botaniky PřF UP Olomouc
- Videa zachycující pohyb svěracích buněk průduchů na Utah State University