Kvasinky
Kvasinky jsou jednobuněčné houbové mikroorganismy. Většina kvasinek patří do třídy vřeckovýtrusných hub, některé však i do třídy hub stopkovýtrusných, a proto společně netvoří taxonomickou skupinu. Netvoří plodnice, množí se zejména nepohlavně a je pro ně charakteristický způsob dělení buněk, takzvané pučení. Mohou se množit i sexuálně tvorbou vřecek, která však nejsou uzavřená v žádných plodnicích (tedy askokarpech).
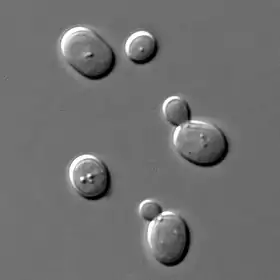
Netvoří žádné pravé myceliální struktury, pouze pseudomycelium, které se podobá koloniím jednobuněčných organismů.
Kvasinky jsou hojně využívány v potravinářství a biotechnologiích. Používají se například při výrobě vína, piva nebo chleba. Kvasinky pro kávu a kakao mají ale větší biodiverzitu.[1] Využívá se jejich schopnosti kvašení. Jsou ale mezi nimi i původci nemocí, jako je např. Candida albicans.
Výzkum a historie

Kvasinky jsou lidmi využívány nejméně od neolitu. Podle chemické analýzy keramiky pocházející z pravěké Číny[2] a oblasti dnešní Gruzie[3] a Íránu[4] byly kvašené nápoje používány nejméně 7000 let před naším letopočtem. Keramika z Číny uchovávala alkoholický nápoj ze směsi kvašené rýže, medu a ovoce s obsahem kyseliny vinné (vinnou révu nebo spíše Crataegus pinnatifida či jiný druh Čínského hlohu).
První jednotlivé kvasinky poprvé pozoroval Antoni van Leeuwenhoek, který popsal ve svých dopisech Královské společnosti v Londýně výsledky pozorování malých kuliček v pivě pomocí primitivního mikroskopu. Theodor Schwann roku 1837 popřel účast kyslíku při kvasném procesu a popsal v kvasící tekutině se rozmnožující kvasinky, které označil za „cukernou houbu“ (odtud rodové označení Saccharomyces).
Objev moderních sekvenovacích principů umožnil již v roce 1996[5] sekvenování genomu kvasinky jako vůbec prvního eukaryotního genomu.
Význam a využití

Kvasinky jsou mikroorganismy v mnohém směru pro člověka velmi užitečné. Už pojmenování „kvasinky“ ukazuje na jejich vztah ke kvasným procesům. Nejstarší nález nádobek na víno pochází z neolitické kuchyně v Hajji Firuz Tepe (8500–4000 let př. n. l.) na území dnešního Íránu.[6] Kvašení bylo známo již ve starém Babylónu v období 6000–4000 let před naším letopočtem, kdy se ze zkvašeného odvaru z naklíčeného obilí připravoval nápoj, jenž se dá považovat za předchůdce piva.
V polovině 19. století vznikl drožďárenský průmysl[7]. Předtím proběhlo několik pokusů o průmyslovou výrobu v Holandsku. Nejdříve bylo droždí prodáváno v tekuté formě, později po zdokonalení Tebbenhofem se přešlo k lisovanému droždí.
Prvořadý význam mají dnes v kvasném průmyslu při výrobě lihu, piva, vína, pekařského droždí (viz článek Saccharomyces cerevisiae) a také některých mléčných nápojů. V krmivářském průmyslu nabývají význam krmné směsi z kvasnic. Doposud jsou poměrně málo využívány pro potravinářské účely jako zdroj bílkovin a jiných biologicky cenných látek. Mají bohatý obsah dobře stravitelných bílkovin (kolem 40 procent), cukrů a zejména komplexu vitamínu B, což platí zvláště pro pivovarské kvasinky. Ty se využívají i při léčbě nervových onemocnění, při zánětlivých kožních chorobách, při poruchách zažívacího traktu a jaterních chorobách. Geneticky modifikované kvasinky bude možné v blízké budoucnosti použít i na odhalování výbušnin, přesněji molekul dinitrotoluenu (DNT).[8]
Vedle převažujícího pozitivního významu existuje i negativní vliv kvasinek patogenních, či nevhodný výskyt kvasinek coby škodlivého kontaminantu ve výrobním procesu.
Strukturní komponenty
Kvasinky vykazují velkou tvarovou, velikostní či barevnou diverzitu. Obvykle jsou kvasinky kulaté nebo oválné, ale vyskytují se i druhy, které mají buňky charakteristického citronovitého, vajíčkovitého, lahvovitého či vláknitého tvaru. Dokonce mezi samotnými buňkami jednoho kmene lze nalézt morfologické a barevné heterogenity. To je zapříčiněno změnami fyzikálních a chemických podmínek v prostředí. Pro jednoduchost ukažme strukturu kvasinky Saccharomyces cerevisiae.
S. cerevisiae jsou obecně elipsoidní, v průměru od 5–10 μm na velké ose, a od 1–7 μm na menší ose. Střední objem buněk je 29-55 μm3 pro haploidni resp. diploidní buňky. Velikost buňky se zvětšuje s věkem.
Shrnutí makromolekulárních látek v kvasince viz tabulka:
| typ makromolekuly | kategorie | základní komponenty |
|---|---|---|
| proteiny | strukturní | aktin, tubulin, histony, ribosomální proteiny |
| hormony | alfa a feromony | |
| enzymy | ||
| Glykoproteiny | komponenty buněčné stěny | mannoproteiny |
| enzymy | funkční enzymy (invertáza) | |
| polysacharidy | komponenty buněčné stěny | glukan, mannan, chitin |
| komponenty kapsulární | ||
| zásobní | glykogen, trehalóza | |
| polyfosfáty | zásobní | polyfosfáty ve vakuole |
| lipidy | strukturní | volné steroly v membráně |
| zásobní | triglyceridym, sterolové estery | |
| funkční | deriváty fosfoglyceridů, volné mastné kyseliny | |
| nukleové kyseliny | DNA | genová DNA (80%), mitochondriální |
| RNA | rRNA (80%)m mRNA (5% cytosol, ER, mitochondrie), tRNA |
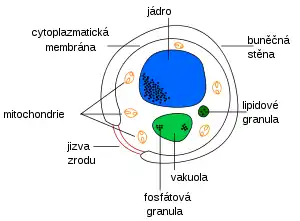
Buněčná stěna

V roce 1970 Kidby a Davis navrhl model buněčné stěny pro rod S. cerevisae, který stále ještě rámcově platí. Podle tohoto modelu má buněčná stěna tři vrstvy: vnější, střední a vnitřní, přičemž polysacharid-proteinové komplexy jsou podle tohoto modelu spojeny fosfodiesterovými vazbami.
- vnější, jež je orientována směrem do prostředí, obsahuje mannanproteiny spojené disulfidovými můstky.
- střední je tvořena β-1,6 glukanem, glukanproteiny a mannanproteiny
- vnitřní, přiléhající na cytoplazmatickou membránu, je složena z mikrokrystalického β-1,3-glukanu
V buněčné stěně se nacházejí také hydrolytické enzymy glykoproteinového typu, např. β-fruktofuranosidasa (invertáza).
Polysacharidy buněčné stěny mohou být jednosložkové (homopolysacharidy) nebo vícesložkové (heteropolysacharidy) – např. galaktomannany, xylomannany, arabomannany aj. Polysacharidy určují především imunologické vlastnosti kvasinek, neboť působí jako antigeny. Jedná se především o mannanproteiny, jejichž antigenní charakter je určen délkou bočních řetězců mannosylových zbytků, spojených α -1,2 a α -1,3 vazbami. Polysacharid – proteinové komponenty buněčné stěny se taktéž podílejí na flokulačních schopnostech kvasinek.
Jizvy
Jako stálé struktury na buněčné stěně vznikají jizvy po pučení dceřiných buněk a jejich separaci od mateřské buňky, popř. po dělení kvasinkových buněk. Přetrvávají po celý vývoj jednotlivé buňky a ovlivňují tzv. architekturu její stěny. Vznik jizev a jejich stavbu lze sledovat např. v optickém mikroskopu s využitím fluorescence primulinu, což je barvivo, které se specificky váže na mikrofibrily.
Jizva na dceřiné buňce se jmenuje „jizva zrodu“, a protože je tvořena jiným buněčným materiálem, lze ji odlišit od jizev na místech, kde se oddělily nové pupeny. Z každé buňky může vypučet jen omezený počet nových buněk, v souvislosti s tím, jak buňka stárne.[9][10]
Cytoplazmatická membrána
Cytoplazmatická membrána (čili plazmalema) tvoří elastický obal protoplastu, osmotickou bariéru a kontroluje transport látek. Je tvořena především z fosfolipidů a bílkovin.
Struktura cytoplazmatické membrány je patrně nejlépe popsána v rámci mozaikového modelu buněčných membrán, který říká, že molekuly fosfolipidů jsou uspořádány ve dvou vrstvách tak, že polární části jsou na vnější stranách a nepolární uvnitř membrány (viz též Lipidová dvouvrstva).
Pro plazmalemu kvasinek jsou charakteristické invaginace do cytoplasmy, které jsou četné rovněž v exponenciální fázi růstu.
Jádro
Jádro bývá u eukaryotních organismů zřetelně ohraničeno jadernou membránou a tvoří ho chromatin, tedy komplex dsDNA, histonů a proteinů nehistonové povahy. Na rozdíl od většiny eukaryoriotických buněk mají kvasinky tzv. endomitózu, takže dělení jádra probíhá bez rozrušení jaderné membrány. To je hlavní důvod, proč je u kvasinek mimořádně obtížné udělat dobrý karyotyp a zjistit tak přesný počet a stavbu chromozomů. Na jaderné membráně se diferencuje tzv. polární tělísko, které hraje významnou roli při iniciaci tvorby pupene.
Sekreční dráha u kvasinek
Biomembrány tvoří povrchové struktury organel i systém vnitřních membrán. Tím buňku rozdělují na různé reakční prostory. V některých z nich dochází k sekreci proteinů. V endoplazmatickém retikulu jsou syntetizovány peptidy a bílkoviny. Na vnějším povrchu ER (drsné ER) jsou zakotveny ribozomy, pohybují se ale i volně v cytoplazmě. Stejně jako u ostatních eukaryot se ribozom skládá z malé (40S) a velké (60S) podjednotky. Velká podjednotka katalyzuje syntézu bílkovin. Na tvorbě ribozomů se podílí jadérko. Vnitřní povrch ER tvoří hladké ER. V ER dochází i k posttranslačním úpravám, na místo určení se proteiny dostávají pomocí sekrečních měchýřků. Ty tvoří Golgiho aparát.
Endoplazmatické retikulum
Endoplazmatické retikulum vytváří u kvasinek lamely, cisterny a tubuly podobně jako u rostlinných a živočišných buněk. Jeho obsah (enchylema) je uzavřen dvěma membránami. ER navazuje na vnitřní povrch cytoplazmatické membrány a někdy je spojeno s jadernou membránou.
Ribozomy
Ribozomy kvasinek mají obdobně jako u jiných eukaryot hodnotu sedimentačního koeficientu kolem 80S. Menší podjednotka o koeficientu 40S je složena z jediné molekuly 18S rRNA a z 30 ± 5 bílkovin; větší podjednotka o 60S obsahuje 28S rRNA, 5S rRNA, 5,8S rRNA a 40 ± 5 bílkovin.
Mitochondrie
Mitochondrie jsou organely specializované na respiraci a oxidativní fosforylaci s vlastním genetickým systémem a proteosyntézou. Tvar, struktura a počet mitochondrií mohou být ovlivňovány různými faktory. Např. v podmínkách reprimujícího dýchání se výrazně snižuje podíl, který mitochondrie zaujímají v celkovém objemu buňky.
Vakuola
Kvasinková buňka zpravidla obsahuje jednu velkou kulatou vakuolu. V počáteční fázi pučení lze pozorovat větší množství malých vakuol, které však s růstem splývají v jednu vakuolu, případně dvě vakuoly (ve stacionární fázi růstu). Průměr vakuol je proměnlivý od 0,3 do 3 μm. Ohraničené jsou membránou zvanou tonoplast.
80 až 90 % rozpustných aminokyselin z celkového objemu volných aminokyselin v kvasinkách je obsaženo ve vakuolách. Dále se zde nacházejí enzymy hydrolázy, většina proteáz, ribonukleázy či esterázy. Těmito enzymy se ve vakuolách rozkládají funkčně narušené organely.
Buněčný cyklus
Buněčný cyklus obecně
Mnohé kvasinky jsou schopné za určitých okolností omezit rozmnožování buněk a začít buňky prodlužovat. Tímto způsobem vznikají vláknitá pseudomycelia, jejichž tvar je pro jednotlivé druhy charakteristický.
Buněčným cyklem rozumíme obecně buněčné procesy mezi dvěma následujícími děleními buňky. Podrobně je tento cyklus znám především u Saccharomyces cerevisae a u Schizosaccharomyces pombe.
Životní cyklus kvasinek Schizosaccharomyces pombe
V roce 1893 Lindner objevil Schizosaccharomyces pombe v prosném pivu ve východní Africe, jehož místní jméno bylo pombe. Kmen užívaný v současnosti pro genetické výzkumy byl izolován Ursem Leupoldem (Univerzita Berne, Švýcarsko) v roce 1950 z kvasničné kultury S. liquefacis.
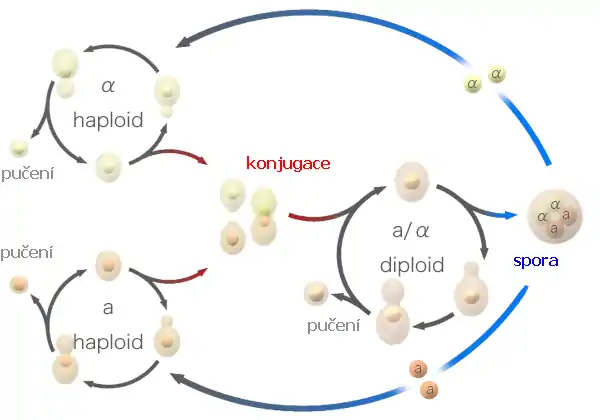
V průběhu normálního životního cyklu jsou kvasinky haploidní, tj. mají po jedné kopii od každého chromozomu, a tedy jedinou kopii od každého genu. Haploidní kvasinky jsou používány ve výzkumech, protože jak recesivní tak dominantní mutace se projeví ve výsledném mutantním fenotypu.
Haploidní buňky se rozmnožují asexuálně prostřednictvím mitózy. Dceřiná buňka vyrůstá na pólu. Když doroste do dospělce, přestane růst a vytváří přehrádku v prostředku buňky. Přehrádka dělí matečnou buňku do dvou ekvivalentních dceřiných buněk. Za příhodných podmínek proběhne celý dělicí cyklus za 3 hodiny. V přírodě jsou kvasinky často nutričně deprivovány. Protože je S. pombe dimorfní kvasinkou, je schopna se „přepnout“ z kvasinkové morfologie do pseudohyfálního stavu, v němž zůstanou dceřiné buňky spojeny. Pseudohyfální růst umožňuje buňkám se efektivněji šířit a shánět se po nových nutričních zdrojích. S. pombe má dva partnerské typy, jmenovitě „+“ a „-“. Pokud je bohatá etapa následována nutriční deficiencí, tyto partnerské typy konjugují a fúzují na koncích Následně fúzují jádra, aby vytvořila diploidní buňku, zvanou zygota.
Obyčejně podstupují zygoty meiózu okamžitě, následuje sporulace a formování 4-sporového zygotického konglomerátu. Stěna toho útvaru autolyzuje, uvolňuje haploidní spory, které jsou schopny přežít delší období stresu.
Když se životní podmínky zlepší, spory začnou růst a životní cyklus haploidních buněk znovu začne.
Diploidní buňky se středově štěpí, jako haploidní buňky, ale jsou delší a širší než haploidní.
Haploidní buňky měří od 7–8 (nově narozené) do 12–15 (při dělení) mikrometrů na délku a 3–4 mikrometry do šířky, zatímco diploidní buňky měří od 11–14 (nově narozené) do 20–25 (při dělení) mikrometrů na délku a 4–5 mikrometrů do šířky.
Diploidní buňky pokračují v mitóze do vyčerpání živin
Rozmnožování kvasinek
Kvasinky se mohou rozmnožovat nepohlavně a to většinou pučením, krom několika málo druhů tzv. poltivých kvasinek, resp. pohlavně za vzniku pohlavních spor.
Vegetativní rozmnožování
Během pučení (nepohlavní reprodukce) se na mateřské buňce vytvoří pupen, který se postupně zvětšuje. Při dosažení dostatečné velikosti dojde k oddělení od mateřské buňky. Podle místa, kde pupen na povrchu kvasinky vzniká, se rozlišuje pučení monopolární, bipolární a multipolární, případně mohou pupeny vznikat i zcela náhodně.
Pohlavní rozmnožování
U většiny kvasinek lze kromě nepohlavního rozmnožování pozorovat i rozmnožování pohlavní, při kterém vznikají spory. Často kvasinky (schopné sexuálního rozmnožování) sporulují při nedostatečném přísunu živin.
Při pohlavním rozmnožování spolu splynou (konjugují) dvě buňky a posléze se spojí i jejich jádra – dojde ke karyogamii. Občas k ní dojde později, takže mezitím existuje dikaryontní fáze, při které se buňky se dvěma jádry dělí a dávají vznik dikaryotickému myceliu. Toto lze pozorovat především u skupiny tzv. bazidiomycet. Konjugací dvou kvasinkových buněk vzniká zygota, která později sporuluje za vzniku asku nebo bazidia, v nichž jsou uloženy spory. Ke sporulaci je nezbytné meiotické dělení, během něhož vzniknou 4 spory, které se však mohou dále dělit, takže někdy je v asku 8, 16 i více spor. Protože ale u kvasinek se sleduje silný sklon k redukci pohlavního rozmnožování, u mnohých druhů spolu splývají dvě somatické (diploidní) buňky, nebo k žádné konjugaci vůbec nedochází (jedná se potom o tzv. imperfektní kvasinky).
Genetika kvasinek
Genetické mapy kvasinek
Pro konstrukci genetické mapy kvasinek byly vypracovány různé metody založené na využití procesu meiózy nebo mitózy, Jednou z nich je často používaná tetrádová analýza, která je základní mapovací metodou. Při použití této metody se předpokládá tvorba vřecek se 4 sporami, které se mikromanipulační technikou (po natrávení stěny vřecka) izolují, a tím se získají 4 haploidní meitoické „klony“.
DNA
Většina genů je soustředěna v chromosomech. Kvasinkový chromosom je tvořen lineární molekulu dsDNA, jejíž velikost se pohybuje kolem 1,107 Mbp. Značnou část tvoří repetice (rozptýlené a krátké tandemové), pravděpodobně se vyskytují i genové repetice.
V kvasinkách se nalézá cca 20–100 molekul v závislosti na kultivačních podmínkách, růstové fázi aj. mitochondriální DNA (mtDNA). Molekuly mtDNA se výrazně liší svou velikostí (17–108 kb), jsou kružnicové v malém počtu i lineární.
Plazmidová DNA obvykle není pro buňky důležitá, protože nekóduje esenciální životní funkce. Ve většině laboratorních kmenů kvasinky S. cerevisiae je možné však v jádře nalézt kruhový 2 μm plazmid. Mimo to kvasinka obsahuje plazmidovou DNA asi v 60 kopiích v haploidním a dvojnásobný počet ve stádiu diploidním. Existence plazmidu v jádře je pro kvasinky značena jako cir+ resp. cir−. 2 μm plazmid se využívá coby základ vektorů používaných v genových manipulacích u kvasinek.
Metabolismus
Dýchání představuje velice účinný způsob využití cukru kvasinkou.[11] Z molekuly glukózy (C6H12O6) kvasinka získá energii postačující k syntéze až 38 molekul ATP.

Vysoký energetický zisk však vyžaduje rozsáhlou počáteční energetickou investici do syntézy a údržby komplexního enzymového aparátu, a nevyplatí se, pokud je glukóza v nadbytku a přísun kyslíku je omezen, což odpovídá situaci v kynoucím těstě.
V takové situaci se kvasinka uchyluje k lihovému kvašení (etanolové fermentaci).
Taxonomické členění

Kvasinky se řadí do říše houby (ačkoliv jsou to mikroskopické jednobuněčné organizmy), superskupiny Opisthokonta, domény (nadříše) Eukaryota. Netvoří však žádnou přirozenou taxonomickou skupinu[12], a proto je nemožné je jednotně definovat. Jako takové jsou roztroušeny ve dvou odděleních hub, buď jako houby vřeckovýtrusné, nebo stopkovýtrusné. Níže uvedený pokus o vyjmenování základních čeledí kvasinek vychází z knihy The yeasts, a taxonomic study z roku 1998,[13] ačkoliv se od té doby objevila řada nových poznatků.
Vřeckovýtrusné kvasinky se nachází v těchto skupinách:
- Primitivní archiaskomycety: čeledi Schizosaccharomycetaceae, Taphrinaceae, Protomycetaceae, Pneumocystidaceae
- Pravé vřeckovýtrusné: rody Endomyces a Oosporidium
- Hemiaskomycety: čeledi Ascoidaceae, Cepaloascaceae, Dipodascaceae, Endomycetaceae, Eremotheciaceae, Lipomycetaceae, Metschnikowiaceae, Saccharomycetaceae, Saccharomycodaceae, Saccharomycopsidaceae, Candidaceae
Stopkovýtrusné kvasinky se do určité míry nachází zejména v čeledích Spolobolomycetaceae, Cryptococcaceae, Filobasidiaceae, Teliosporaceae a v řádu Tremellales.
Významní zástupci kvasinek
Průmyslově významné kvasinky
- Některé kvasinky, např. z rodů Saccharomyces, Kloeckera, Torulopsis způsobují kvašení (alkoholové, octové, mléčné, máselné, propionové) a rozdělují se podle rozložení kvasinkové kultury a vývinu plynu na svrchní a spodní.
- Saccharomyces cerevisiae je všeobecně využívaná kvasinka. Je nezbytná při vaření piva, výrobě vína, pečení chleba aj. Průmyslově využívané kmeny jsou většinou staletí šlechtěné a polyploidní. Mimo to je tato kvasinka jako jednoduchý eukaryotický organismus také zajímavým modelem v molekulární biologii. Ke svému rozmnožování potřebují cukr.
- Kvasinka Rhodotorula glutinis je řazena k významným průmyslovým kmenům. V potravinářství je používána zejména k obohacování krmných směsí vzhledem ke značnému obsahu karotenoidů, které je schopna produkovat i ve stresových podmínkách.
- Kvasinka Phaffia rhodozyma je používána díky vysokému obsahu karotenoidního barviva astaxanthinu jako dietní doplněk výživy lososů a drůbeže. Pokud jsou hydrolyzované buňky přidány ke krmivu, astaxanthin je snadno absorbován ve střevech a mění barvu lososího masa z růžové na oranžovou, nebo barvu žloutku a masa u drůbeže.
- Yarrowia lipolytica (zástupce poltivých kvasinek) je schopna metabolizovat ropné produkty, používá se k výrobě kyseliny citronové.
Patogenní kvasinky
Mezi patogenní druhy kvasinek patří např. zástupci rodů Candida, Cryptococcus, Trichosporon, kteří vyvolávají různá kožní, slizniční aj. onemocnění.
- Candida albicans, zástupce imperfektních kvasinek, je patogen urogenitálního traktu. Zde žije zcela běžně, ale je-li potlačena přirozená poševní mikroflóra (např. léčba antibiotiky) může se začít množit a působit značné obtíže.
- Cryptococcus neoformans je nebezpečný patogen napadající nervovou soustavu. Hlavními přenašeči jsou holubi.
- Trichosporon cutaneum je kožní patogen (zástupce poltivých kvasinek) žijící na vousech a ve vlasech.
Léky, používané proti kvasinkám, kvasinkovým mikroorganismům a plísním (mykózy) se nazývají antimykotika. Desinfekční látky chemicky hubící kvasinky se nazývají levurocidní (levure - kvasinka, droždí, z francouzštiny).
Odkazy
Reference
- http://phys.org/news/2016-03-diverse-yeasts-coffee-chocolate.html - You can thank diverse yeasts for that coffee and chocolate
- MCGOVERN, P. E. Fermented beverages of pre- and proto-historic China. Proceedings of the National Academy of Sciences. 2004-12-15, roč. 101, čís. 51, s. 17593–17598. DOI 10.1073/pnas.0407921102.
- World's Earliest Wine [online]. Archaeological Institute of America [cit. 2014-02-08]. Dostupné online.
- 7,000 Year-old Wine Jar [online]. University of Pennsylvania Museum of Archaeology and Anthropology [cit. 2014-02-08]. Dostupné online.
- Co se o sobě dovídáme z naší genetické informace[nedostupný zdroj]
- Kvasinky člověku věrnější než pes. www.vesmir.cz [online]. [cit. 29-09-2007]. Dostupné v archivu pořízeném dne 29-09-2007.
- Kvasná chemie aneb vznik piva, vína u nás z historického hlediska
- Populární článek (na Idnes) o studii odborníků filadelfské univerzity, která byla zveřejněna v časopisu Nature Chemical Biology.
- CHEN, Ines. Yeast as budding stem cells?. Nat Struct Mol Biol. 2009-04, roč. 16, čís. 4, s. 351. Dostupné online. ISSN 1545-9993. DOI 10.1038/nsmb0409-351.
- LAUN, Peter, Carlo V. Bruschi, J. Richard Dickinson, Mark Rinnerthaler, Gino Heeren, Richard Schwimbersky, Raphaela Rid, Michael Breitenbach. Yeast mother cell-specific ageing, genetic (in)stability, and the somatic mutation theory of ageing. Nucl. Acids Res.. 2007-12-03, roč. 35, čís. 22, s. 7514–7526. Dostupné online. DOI 10.1093/nar/gkm919.
- ODPOVĚĎ NA KAŽDOU OTÁZKU. www.vesmir.cz [online]. [cit. 29-09-2007]. Dostupné v archivu pořízeném dne 29-09-2007.
- Kocková-Kratochvílová A.: Kvasinky a kvasinkovité mikroorganizmy, Alfa, Bratislava, 1982, str. 17
- KURTZMAN, C. P.; FELL, Jack W. The yeasts, a taxonomic study. [s.l.]: Elsevier, 1998. Dostupné online.
Literatura
- BARNETT J. A., PAYNE R. W., YARRPW D. Yeasts charakteristics and identification. Cambridge: Cambridge University Press, 2000. Dostupné online. ISBN 0-521-57396-3. (anglicky)
- BURKE DAN, DAWSON DEAN, STEARNS TIM,. Methods in yeast genetics: a Cold Spring Harbor Laboratory course manual. New York: Cold Spring Harbor Laboratory Press, 2000. ISBN 0-87969-588-9. (anglicky)
- JANDEROVÁ, B., BENDOVÁ, O. Úvod do biologie kvasinek. Univerzita Karlova v Praze: Nakladatelství Karolinum, 1999.
- KOCKOVÁ–KRATOCHVÍLOVÁ A. Kvasinky a kvasinkovité mikroorganizmy. Bratislava: Alfa, 1982.
- KOCKOVÁ–KRATOCHVÍLOVÁ A. Kvasinky ve výzkumu a praxi. Praha: Academia S. 1986.
- KURTZMAN CLETUS P., FELL JACK W. The Yeast: a taxonomic study. Amsterdam: Elsevier, 1998. ISBN 0-444-81312-8.
- RYCHTERA M., PÁCA J. Bioinženýrství kvasných procesů. Praha: VŠCHT, 1987.
- ŠIPICKÝ, M., ŠUBÍK, J. Genetika kvasinek. Bratislava: Veda, 1992.
- VODRÁŽKA Z. Biotechnologie. Praha: [s.n.] ISBN 80-7080-121-2. S. 1991.
Externí odkazy
 Obrázky, zvuky či videa k tématu kvasinky na Wikimedia Commons
Obrázky, zvuky či videa k tématu kvasinky na Wikimedia Commons  Slovníkové heslo kvasinka ve Wikislovníku
Slovníkové heslo kvasinka ve Wikislovníku- Miniatlas mikroorganismů [online]. [cit. 2013-10-20]. Dostupné v archivu pořízeném dne 2007-09-29.
- Vše o droždí [online]. [cit. 2013-10-20]. Dostupné online.