Interleukin 15
Interleukin 15 (IL-15) byl v roce 1994 objeven a charakterizován jako růstový faktor buněk. IL-15 je 14 - 15 kDa velký glykoprotein, který svoji terciární strukturou patří do rodiny cytokinů obsahujících čtyři alfa-helixové domény.[1] Kromě IL-15 se do této rodiny cytokinů řadí také interleukin-2 (IL-2), interleukin-4 (IL-4), interleukin-7 (IL-7), interleukin-9 (IL-9), granulocyte colony-stimulating factor (G-CSF) a granulocyte-macrophage colony-stimulating factor (GM-CSF).[2]
Interleukin 15 je konstitutivně produkován velkým množstvím buněčných typů, jako jsou monocyty, makrofágy, dendritické buňky (DC), keratinocyty, fibroblasty a nervové buňky. Navíc byla jeho produkce pozorována v mnoha tkáních, jako jsou ledvina, placenta, plíce, srdce nebo kosterní sval.[2] Díky svým pleiotropním působením hraje IL-15 významnou roli jak ve vrozené tak v adaptivní imunitní odpovědi. Navíc se podílí na regulaci imunitního systému v odpovědi na infekci bakteriálního či virového původu. Důležitou roli hraje IL-15 také v protinádorové odpovědi, alergiích a v autoimunitních chorobách.[3]
Exprese IL-15
Interleukin 15 je kódován genem velkým 34 kb, nacházející se u člověka na chromozomu 4q31 a u myší v centrálním regionu chromozomu 8.[4] Gen IL-15 tvoří 9 exonů (1 - 8 a 4A) a 8 intronů. Pouze 4 exony (5 - 8) kódují maturovaný protein (Obr. 2).[1]

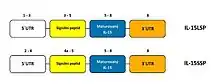
Alternativním sestřihem jsou u člověka a také u myší vytvářeny dvě izoformy IL-15, které se odlišují délkou svého signálního peptidu. Delší izoforma (IL-15 LSP) obsahuje 48 aminokyselin dlouhý leader peptid, na 5´-konci netranslatovaný region (5'UTR) o velikosti 316 bp, 486 bp dlouhou kódující sekvenci a na 3´-konci 400 bp dlouhý UTR region.[1] IL-15 LSP izoforma byla identifikována v Golgiho aparátu, časných endozomech a v endoplasmatickém retikulu. Proto se může vyskytovat jako sekretovaná forma a/nebo vázaná na membránu nejčastěji dendritických buněk. Navíc je IL-15 LSP izoforma exprimována tkáněmi kosterního svalstva, placentou, srdcem, plícemi, játry a ledvinami.[5] Izoforma s krátkým signálním peptidem (IL-15 SSP) o velikosti 21 aminokyselin je kódována exony 4A a 5. Tato krátká izoforma byla identifikována zejména ve tkání testes a thymu. IL-15 SSP je lokalizován v cytoplazmě, nebo je translokována do jádra. IL-15 SSP nevytváří sekreční formu proteinu.[1] Signální peptidy obou izoforem sdílejí sekvenci 11 identických aminokyselin, které jsou kódovány exonem 5. IL-15 LSP a IL-15 SSP kódují stejný maturovaný protein.[6]
U myší, jsou alternativním sestřihem variabilního exonu 5, vytvářeny dvě izoformy interleukinu 15. Izoforma kódována IL-15 mRNA s normálním exonem 5 vytváří sekretovanou formu. Na druhou stranu, izoforma obsahující alternativní exon 5 s jiným 3´-sestřihovým místem, má vyšší translační účinnost a postrádá hydrofobní doménu signální sekvence, a proto je lokalizována intracelulárně.[6]
Produkce IL-15 je kontrolována mnoha faktory na úrovni transkripce, translace a vnitrobuněčného pohybu. Nejdůležitější regulace syntézy IL-15 probíhá na posttranskripční úrovni.[3] Ačkoliv je mRNA interleukinu 15 konstitutivně exprimována v mnoha tkáních a buněčných typech, jako jsou fibroblasty, svalové buňky, keratinocyty, adipocyty či nádorové buňky, představuje IL-15 cytokin, který je produkován zejména dendritickými buňkami, monocyty, makrofágy a stromálními buňkami. Na rozdíl od interleukinu 2, není produkován T buňkami. Rozdíl mezi expresi IL-15 mRNA a produkci proteinu je způsoben posttranskripční regulací, která vede k odpojení translace a udržování netranslatovaného mRNA IL-15 v buňce. Translace mRNA uložené v buňce může být indukována specifickými signály. Základním regulačním mechanismem tohoto procesu představuje 5´-netranslatovaný region IL-15 mRNA, obsahující 12 upstream iniciačních translačních AUG kodonů u člověka a 5 u myší.[7] Navíc důležitou roli v regulaci, hrají také proteinové sekvence signálního peptidu a C-konce maturovaného proteinu.[8] Produkce interleukinu 15 může být stimulovaná pomocí GM-CSF, dvouvláknové mRNA, oligonukleotidy s metylovanými CpG, lipopolysacharidem (LPS), interferonem gama (IFN-γ), nebo také po infekci monocytů herpes virem, Mycobacterium tuberculosis či Candida Albicans.[7] V indukci produkce IL-15 se uplatňují zejména 5´konzervované regulační motivy nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) a IRF response element (IRF-E) (Obr. 3).[4]
Signalizace IL-15
Působení interleukinu 15 zahrnuje juxtakrinní (cell-to-cell contact), intrakrinní (přímo v buňce) a reverzní signalizaci. Zpočátku byl interleukin 15 charakterizován jako solubilní molekula. Později bylo prokázáno, že IL-15 existuje také v membránově vázané formě, která představuje majoritní formu. Na membránu může být vázán přímo nebo v komplexu s IL-15Rα. V obou případech je součástí imunologický synapse.[7]
Hlavním způsobem signalizace interleukinu 15 představuje trans-prezentace, která je zprostředkována receptorem IL-15Rα, vytvářejícím s IL-15 stabilní, na membránu vázaný komplex IL-15Rα/IL-15 (Obr. 6).[9]
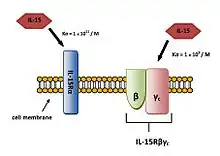
Receptor IL-15Rα váže IL-15 s vysokou afinitou (Ka = 1×1011/M), na rozdíl od receptoru IL-15Rβγc, který váže IL-15 s afinou o něco nižší (Ka = 1×109/M)(Obr. 7).[3]
Juxtakrinní signalizační dráha začíná vazbou IL-15 na IL-15Rα receptor, s následnou prezentací IL-15 v komplexu okolním buňkám nesoucím na svém povrchu IL-15βγc receptor. Následně prostřednictvím β a γc řetězce receptorového komplexu, dochází k aktivaci Janus kinase 1 (JAK1) β řetězcem a Janus kinase 2 (JAK2) γc řetězcem komplexu. Jejich aktivace vede k fosforylaci a aktivaci signal transducer and activator of transcription 3 (STAT3) a STAT5, které podobně jako v případě IL-2 indukují změnu v B-cell lymphoma 2 (Bcl-2), MAP (mitogen-activated protein kinase) kinázové dráhy a fosforylaci Lck (Lymphocyte-specific protein tyrosine kinase) a Syk (spleen tyrosine kinase) kinázy. Výsledkem signální kaskády je proliferace a maturace buněk (Obr. 8).[3][10]
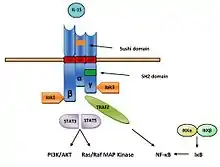
U adipocytů dochází k aktivaci JAK2 a STAT5 místo původní aktivace JAK1/3 a STAT3/5. Fosforylací STAT se formují transkripční faktory, které následně regulují transkripci příslušným genů. β řetězec aktivuje tyrozin kinázy rodiny Src, (Lck, Fyn a Lyn). Navíc může aktivovat také fosfatidylinositol 3 kinázu (PI3K) AKT signální dráhy, čímž reguluje expresi transkripčních faktorů c-Fos, c-Jun, c-Myc a NF-κB. Na druhou stranu, receptor IL-15Rα může přenášet signál prostřednictvím Syk protein kinázy rodiny Syk/ZAP70 a následně aktivuje protein tyrosin kinázu AXL.[7] Interleukin 15 se může vázat na IL-15βγc signální komplex bez přítomnosti IL-15Rα receptoru. Po vazbě, aktivuje IL-15 kinázy z rodiny Src, Lck a Fyn, které následně aktivují PI3K a MAPK signální dráhy.[11]
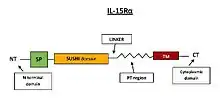
Signalizace interleukinem 15 může pobíhat kromě trans-prezentace také cis-prezentací, kdy dochází k interakci IL-15 se signálním komplexem IL-15Rβγc na povrchu téže buňky, prostřednictvím vysokoafinitního receptoru IL-15Rα. Tento způsob signalizace umožňuje flexibilita C-konce ektodomény IL-15Rα, prostřednictvím linkeru o velikosti 32 aminokyselin, a/nebo 74 aminokyselin dlouhým PT regionem (Obr. 4). Díky tomu leží C-konec IL-15Rα v rovině téměř rovnoběžné s plazmatickou membránou.[9]
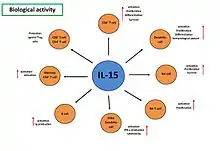
Biologická aktivita IL-15
Role interleukinu 15 ve vrozeném a adaptivním imunitním systému byla poprvé prokázána na IL-15Rα-/- a IL-15-/- myších. U těchto myší bylo pozorováno snížení celkového množství CD8+ T lymfocytů. Navíc bylo pozorováno, že myši byly deficientní v populaci paměťových CD8+ T lymfocytů (CD8+CD44hiCD122hi), NK buněk, NK-T buněk a v populaci intraepiteliálních lymfocytů (EIL). Na druhou stranu, u IL-15 transgenních myší, byla pozorována vysoká hladina IL-15 a také zvýšené množství periferních NK buněk, NK-T buněk, CD44hiCD8+ T lymfocytů a fyziologická hladina CD4+ lymfocytů.[12]
Vliv IL-15 na buňky vrozené imunity
NK buňky (natural killers) představují efektorové buňky vrozené imunity, které hrají klíčovou roli v imunitní odpovědi proti virové nebo bakteriální infekci a v protinádorové imunitě. Neaktivované NK buňky konstitutivně exprimují IL-15Rα receptor a IL-15Rβγc signální komplex, které jsou nezbytné pro rozpoznání, vazbu a působení membránově vázaného IL-15, trans-prezentovaného zejména na dendritických buňkách.[12] Interleukin 15 hraje klíčovou roli ve vývoji, diferenciaci a přežívání NK buněk (Obr. 5). Hlavním zdrojem IL-15 pro NK buňky, představuje trans-prezentace CD11+ dendritickými buňkami, které indukují diferenciaci NK buněk, up-regulaci aktivačních a inhibičních receptorů LY-49. Navíc IL-15 ve spolupráci s IL-2 indukuje vývoj CD56+ NK buněk z CD34+ hematopoetických progenitorových buněk, které jsou významnými producenty interferonu γ (IFN-γ), tumor necrosis factor α (TNF-α) a GM-CSF. Kromě toho, IL-15 s IL-2 a IL-12 ovlivňují cytotoxickou aktivitu NK buněk, a to zejména stimulací CD56dimCD16+ k produkci IIFN-γ, ale také prostřednictvím up-regulace receptorů NKG2D a MICA, expresí efektorových molekul TRAIL.[11]
Monocyty cirkulující v krevním oběhu, jsou po aktivačním signálu transportovány do místa zánětu, kde dávají vznik makrofágům, které spolu s dendritickými buňkami hrají důležitou roli v regulaci adaptivní imunitní odpovědi jako antigen prezentující buňky (APC).[2] Myší monocyty exprimují IL-15 mRNA po indukcí IFN-γ a mikrobiálními stimuly. Naopak u lidí, monocyty exprimují membránově vázanou formu IL-15 která je podobně jako u myší regulována pomocí IFN-γ. Navíc u lidí, IL-15 stimuluje produkci interleukin 8 (IL-8) a monocyte chemotactic protein 1 (MCP-1) a chemokiny, které se podílejí na přitahování neutrofilů do místa zánětu.
Autokrinním působením IL-15 u makrofágů, zvyšuje produkci prozánětlivých cytokinů, jako TNF-α, IL-1 a IL-6. Naopak nízká hladina IL-15 vede ke zvýšené produkci proti-zánětlivého cytokinu IL-10. Výsledkem působení IL-15 u monocytů a makrofágů je zvýšení jejich fagocytární aktivity.[11]
Dendritické buňky produkují IL-15 jako odpověď na IFN typ I, ligaci CD40, dvouvláknovou RNA nebo lipopolysacharid (LPS). Navíc se interleukin 15 podílí na diferenciaci DC buněk, upregulaci kostimulačních molekul a také k produkci IFN-γ. IL-15 se také podílí na regulaci produkce IL-2, který působí jako růstový faktor T lymfocytů. Působení interleukinu 15 na folikulární dendritické buňky v germinálních centrech, vede k proliferaci a zvyšuje sekreci chemokinů.[2][11]
Neutrofily představují subpopulaci buněk, která se jako první nachází v místě zánětu, kde indukuje antimikrobiální mechanizmy. Neutrofily konstitutivně exprimují IL-15Rα receptor a také signální komplex IL-15Rβγc. Po vazbě IL-15, dochází k zvýšení exprese aktinu, který zprostředkovává cytoskeletální přestavbu, a tím zvyšuje fagocytární aktivitu neutrofilů v místě zánětu.[11] Navíc interleukin 15 inhibuje apoptózu u neutrofilů, eozinofilů, Fas-mediovanou apoptózu B lymfocytů a také T buněk. Tento antiapoptický efekt IL-5 na granulocyty a také lymfocyty je zprostředkován up-regulací exprese proteinu rodiny Bcl-2 (Bcl-2, Bcl-x a Mcl-1) a downregulací pro-apoptických proteinů Bax, Bad, Bim, Noxa a Puma.[7] Působení interleukinu 15 se zvyšuje v neutrofilech sekrece IL-8, exprese major histocompatibility komplex II (MHC-II). Tím umožňuje neutrofilům prezentovat antigen jako APC buňka.[11]
Adipocyty exprimují alternativní IL-15 receptor, který je označován jako IL-15RX. Signalizace zprostředkována tímto receptorem v tukových buňkách probíhá nezávisle na signálním komplexu IL-15Rβγc.[3] Signální dráha pokračuje down-stream aktivací JAK2 a STAT5 místo původní aktivace JAK1/3 a STAT3/5, které jsou aktivovány v T lymfocytech. Signální dráha vede k proliferaci adipocytů.[11]
Vliv IL-15 na buňky adaptivní imunity
Účinek interleukinu 15 na CD8+ a CD4+ T lymfocytu závisí na jejich aktivačním a vývojovém stádiu. Signalizace je zprostředkována IL-15Rα receptorem a je klíčovým faktorem pro udržování CD8+ T lymfocytů ve všech vývojových stádiích.[2] Působením IL-15 dochází ke zvýšení exprese řady anti-apoptických molekul, jako Bcl-2 u naivních CD44lowCD8+ T lymfocytů, nebo Bcl-2 a Bcl-xl v paměťových CD44highCD8+ T lymfocytech.[11] Navíc v paměťových CD4+ a CD8+ T lymfocytech indukuje interleukin 15 proliferaci a rezistenci k inhibičnímu působení T regulačních buněk (Treg), (Obr. 5). Spolu s IL-7 hraje IL-15 klíčovou roli v homeostatické proliferaci CD4+ T lymfocytů.[7] Závislost udržování paměťových CD4+ T lymfocytů na signálu IL-15 se liší od paměťových CD8+ T lymfocytů. Pozorovaný efekt koreluje s rozdílnou expresí IL-15Rβ (CD122) řetězce, jehož exprese je regulována dvěma T-box transkripčníma faktory, T-bet a Eomes, které se podílejí na diferenciaci naivních CD8+ T do efektorových a paměťových T lymfocytů.[13] Odpověď CD4+ T lymfocytů na IL-15 závisí zejména na jejich aktivačním stavu. Signalizace prostřednictvím TCR receptoru,za přítomnosti kostimulačního signálu, navozuje IL-15 klidový fenotyp CD4+ T lymfocytů, který je charakterizován sníženou expresí CD25, CD71 a CD95. U aktivovaných CD4+ T lymfocytů zvyšuje expresi CD40L (CD154) a tím i schopnost interagovat s APC buňkami prostřednictvím TCR receptoru bez kostimulačního signálu CD3 a CD28.[11]
U B lymfocytů se interleukin 15 podílí na proliferaci, diferenciaci a produkci imunoglobulinů.[1] Ačkoliv IL-15 nestimuluje B lymfocyty v klidovém stádiu, podílí se na proliferaci jako kostimulační signál a spolu s CD40L stimuluje sekreci polyklonálnich imunoglobulinů IgM, IgG1 a IgA plazmatickými buňkami.[7]
Role IL-15 v nemoci
Porucha exprese a signalizace interleukinu 15 se podílí na manifestaci mnoha autoimunitních onemocnění. Příkladem může být pemphigus vulgaris, revmatoidní artritida, systémový lupus erythematodes, sarkoidóza, roztroušená skleróza a celiakie.[1]
Virové onemocnění
Interleukin 15 hraje důležitou roli v imunitní odpovědi během HIV infekce.[14] Po virovém přenosu dochází k nárůstu virové zátěže v akutní fázi infekce, která vede k zvýšené produkci prozánětlivých cytokinů, produkovaných dendritickými buňkami, makrofágy, monocyty, NK buňkami a T lymfocyty. V další fázi nemoci, HIV-1 infekce ničí CD4+ a CD8+ T lymfocyty, B lymfocyty, NK buňky a také buňky nelymfoidního původu. U pacientu s HIV-1 infekci bylo pozorována snížena hladina IL-15.[11] V důsledku této defektní produkce mají specifické CD8+ T lymfocyty zvýšenou senzitivitu k CD95 /Fas – indukované apoptóze, a proto jsou zabíjeny HIV infikovanými buňkami.[15] Kromě specifických CD8+ T lymfocytů mají klíčovou roli v imunitní odpovědi v HIV infekci, také DC a NK buňky. DC buňky izolované z pacientů produkovali menší množství IL-15 na rozdíl od produkce jiných cytokinů, jako IL-12 nebo IL-18. Snížením hladiny IL-15, vede k redukci počtu NK buněk. Je patrné, že IL-15 představuje efektivní prostředek v imunoterapii infekce HIV, protože se podílí na obnově NK buněk, proliferaci efektorových CD8+ a CD4+ T lymfocytů. Na druhou stranu, právě tyto buňky jsou napadané virovou infekcí, a to by mohlo vést k progresi onemocnění.[1] Kromě pozitivního výsledku, u HIV infekce, byl také prokázán pozitivní vliv IL-15 při imunizací myších modelů herpes simplex virem, virem hepatitidy B či tetanovým toxoidem.[16]
Onemocnění zprostředkované imunitním systémem
Celiakální spurie, neboli celiakie je enteritída zprostředkována imunitním systémem projevující se u citlivých jedinců, po požití proteinů bohatých na prolin a glutamin obsažených například v pšenici (gliadin) nebo ječmeni (secalin). Interleukin 15 je v mukóze produkován epitelovými a dendritickými buňkami, které současně prezentují transglutaminázou (TG) deaminovaný gluten naivním CD4+ T lymfocytům. Následně aktivované Th1 lymfocyty sekretují pro-zánětlivé cytokiny (IFN-γ a IL-21), které se podílejí na destrukci mukózy. Th2 aktivované lymfocyty prostřednictvím cytokinů indukují klonální expanzi B lymfocytů a syntézu anti-gliadin a anti-TG protilátek. Další z mechanismu IL-15 podílejících se na manifestaci onemocnění je inhibice Treg buněk, inhibice apopotózy u intraepiteliálních lymfocytů (EIL) nebo aktivaci perforin/granzym systému.[2]
Nádorová onemocnění
Hlavní efektorové buňky v proti-nádorové imunitní odpovědi jsou cytotoxické CD8+ T lymfocyty a NK buňky, které mají schopnost zabíjet nádorové buňky.[7] IL-15 má potenciál indukovat proliferaci a diferenciaci těchto efektových buněk.[16] Hlavní problém u pacientů s nádory je anergie T lymfocytů na tumor associated antigen (TAA) zprostředkovaná Treg buňkami. IL-15 indukuje v efektorových CD8+ T lymfocytech rezistencí k Treg buňkách a tím potenciálně pozměňuje anergii CD8+ T lymfocytů k TAA.[7] Další využití IL-15 v protinádorové terapii je v kombinaci s IL-6, kdy dochází k aktivaci NK buněk. Tento proces je zprostředkován stimulací aktivačního receptoru NK buněk, NKG2D jehož ligandy, jako MHC class I chain-related protein A (MICA), MHC class I chain-related protein B (MICB) UL-16- binding protein (ULBP) exprimovány na stresových, virem infikovaných či nádorových buňkách.[3]
Reference
- J. C. Steel, et al., "Interleukin-15 biology and its therapeutic implications in cancer," Trends in Pharmacological Sciences, vol. 33, pp. 35-41, 2012. PMID:22032984.
- A. Di Sabatino, et al., "Role of IL-15 in immune-mediated and infectious diseases," Cytokine & Growth Factor Reviews, vol. 22, pp. 19-33, 2011. PMID:21074481.
- J. P. Lodolce, et al., "Regulation of lymphoid homeostasis by interleukin-15," Cytokine & Growth Factor Reviews, vol. 13, pp. 429-439, 2002. PMID:12401478.
- T. A. Waldmann and Y. Tagaya, "The multifaceted regulation of interleukin-15 expression and the role of this cytokine in NK cell differentiation and host response to intracellular pathogens," Annual Review of Immunology, vol. 17, pp. 19-49, 1999. PMID:10358752.
- Y. Tagaya, et al., "Generation of secretable and nonsecretable interleukin 15 isoforms through alternate usage of signal peptides," Proceedings of the National Academy of Sciences of the United States of America, vol. 94, pp. 14444-14449, Dec 1997. PMID:9405632.
- H. Nishimura, et al., "Differential roles of interleukin 15 mRNA isoforms generated by alternative splicing in immune responses in vivo," Journal of Experimental Medicine, vol. 191, pp. 157-169, Jan 2000. PMID:10620614.
- M. Jakobisiak, et al., "Interleukin 15 as a promising candidate for tumor immunotherapy," Cytokine & Growth Factor Reviews, vol. 22, pp. 99-108, 2011. PMID:21531164.
- R. N. Bamford, et al., "The 5 ' untranslated region, signal peptide, and the coding sequence of the carboxyl terminus of IL-15 participate in its multifaceted translational control," Journal of Immunology, vol. 160, pp. 4418-4426, May 1998. PMID:9574546.
- S. K. Olsen, et al., "Crystal Structure of the Interleukin-15 Interleukin-15 Receptor Complex: INSIGHTS INTO TRANS AND CIS PRESENTATION," Journal of Biological Chemistry, vol. 282, pp. 37191-37204, 2007. PMID:17947230.
- K. S. Schluns, et al., "The roles of interleukin-15 receptor α: Trans-presentation, receptor component, or both?," The International Journal of Biochemistry & Cell Biology, vol. 37, pp. 1567-1571, 2005. PMID:15896666.
- P.-Y. Perera, et al., "The role of interleukin-15 in inflammation and immune responses to infection: implications for its therapeutic use," Microbes and Infection, vol. 14, pp. 247-261, 2012. PMID:22064066.
- K. S. Schluns, "Distinct cell types control lymphoid subset development by means of IL-15 and IL-15 receptor expression," Proceedings of the National Academy of Sciences, vol. 101, pp. 5616-5621, 2004. PMID:15060278.
- P. R. Burkett, et al., "IL-15R expression on CD8+ T cells is dispensable for T cell memory," Proceedings of the National Academy of Sciences, vol. 100, pp. 4724-4729, 2003. PMID:12671073.
- M. Amicosante, et al., "Levels of interleukin-15 in plasma may predict a favorable outcome of structured treatment interruption in patients with chronic human immunodeficiency virus infection," Journal of Infectious Diseases, vol. 188, pp. 661-665, Sep 2003. PMID:12934181.
- Y. M. Mueller, et al., "Increased CD95/Fas-induced apoptosis of HIV-specific CD8(+) T cells," Immunity, vol. 15, pp. 871-882, Dec 2001.PMID:11754810.
- A. Diab, et al., "IL-15: targeting CD8+T cells for immunotherapy," Cytotherapy, vol. 7, pp. 23-35, 2005.PMID:16040381.