Fyziologie psa domácího
Fyziologie psa domácího popisuje základní funkce organismu psa domácího a fungování jeho jednotlivých orgánových soustav. Pes domácí je nejstarší domestikované zvíře a je též zvířetem laboratorním, jeho fyziologie je proto velmi dobře prozkoumána. Znalost fyziologických pochodů je dále uplatňována v řízení chovu psů. Mezi jednotlivými plemeny, přestože ta mohou být vzhledově značně odlišná, nejsou ve fungování samotného organismu významnější rozdíly.
Pes domácí je živočich, teplokrevný savec. S člověkem sdílí 80 % genů[1], tyto geny řídí fungování jednotlivých buněk a základní uspořádání orgánů a orgánových soustav. Fyziologie buňky psa a člověka je shodná, stejně jako mnoho dalších fyziologických pochodů.
Fyziologie dýchání

Stejně jako člověk i pes potřebuje ke svému životu kyslík. Při dýchání dochází k výměně plynů mezi krví a vzduchem, pes ze vzduchu získává kyslík a zbavuje se oxidu uhličitého. Vzduch se do organismu dostává nozdrami, umístěnými na nosní houbě psa. Nosní houba je většinou vlhká díky sekretům nosní žlázy (gl. lateralis nasi), ale není to pravidlem a suchá nosní houba není sama o sobě příznakem žádného onemocnění. Nozdra psa má víceméně oválný tvar, na boku ale pokračuje hlubokým žlábkem (sulcus alaris), který vede nahoru a do strany. Toto utváření nozdry společně s uspořádáním slizničních řas v nosní dutině způsobuje, že při nádechu vzduch proudí skrze oválnou část nozdry a při výdechu odchází žlábkem na straně. Pes si tak při dýchání nevíří pachovou stopu.[2][3]. Vdechovaný vzduch prochází nosní dutinou, kde je zvlhčován, filtrován a předehříván na teplotu 30-35 °C.[4] V zadní části nosní dutiny je čichové bludiště s čichovými receptory, kam pes může cíleně vhánět vzduch při čenichání, při normálním dýchání je tato část nosní dutiny mimo činnost.[5]
Vdechovaný vzduch dále prochází do nosohltanu. Uspořádání měkkého patra umožňuje psovi dýchat i ústní dutinou, což je obvyklé po fyzické námaze, při stresu a při zvýšené teplotě vnějšího prostředí.[3] Vzduch pokračuje přes hrtan do průdušnice, která se v hrudníku větví na dvě průdušky. Ty vstupují do párových plic.
Plíce jsou samotným dýchacím orgánem psa. Uvnitř plic je průdušky dále větví, posledním dělením vznikají průdušinky, které přivádějí vzduch do plicních sklípků. Objem plic je přímo úměrný velikosti psa a u psa o hmotnosti 23 kg se průměrně rovná 1500 ml[6] a celková plocha respiračního povrchu, kde probíhá výměna plynů, je 90 m2[6]
Pes dýchá při negativním tlaku, jeho nitrohrudní tlak je vždy nižší než tlak atmosférický, tento rozdíl rozpíná plíce a brání jejich kolabování. Při nádechu se kontrakcí bránice zvětšuje objem hrudní dutiny, plíce se rozpínají, tlak uvnitř klesá a to žene do plic čerstvý vzduch. Na roztahování hrudníku se kromě bránice podílejí také další svaly, a to zevní mezižeberní a také svaly šikmé. V klidu je výdech pasivní děj, kdy se objem hrudníku a plic zmenšuje působením elastických sil. Aktivního výdechu se účastní vnitřní mezižeberní svaly a svaly břišního lisu a u psa je projevem dušnosti.
Dechový objem psa činí 10-20 ml/kg.[7] Počet dechů za minutu se u malého psa se pohybuje v rozmezí 15 až 30, u velkého psa 10 až 20/min.
Fyziologie oběhové soustavy
Oběhová soustava psa je tvořena uzavřenou soustavou cév a komorovým čerpadlem, srdcem. V těle psa, stejně jako u ostatních savců, je oběhová soustava dvojitá, rozdělená na dvě části, systémový a plicní krevní oběh. Uvnitř cév se nachází transportní medium, krev.
Srdce psa má tvar kuželu a je k poměru k tělu velké, tvoří 0,7 %[8] tělesné hmotnosti. Srdce je orgán tvořený srdeční svalovinou, uvnitř se nachází čtyři dutiny, pravá síň a pravá komora, které pumpují krev do nízkotlakého plicního oběhu, a levá síň a tlustostěnná levá komora, která žene krev do velkého oběhu tělního. Mezi síní a komorou a mezi srdcem a plicnicí a srdcem a aortou se nacházejí chlopně, které zajišťují směr pohybu krve. Nárazem krve na zavřené srdeční chlopně vznikají srdeční ozvy, které jsou slyšitelné pomocí fonendoskopu. Místa na hrudníku psa, kde jsou nejlépe slyšet srdeční ozvy na jednotlivých chlopních, se nazývají puncta maxima.
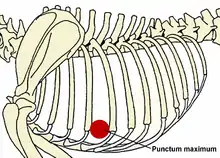
Puncta maxima psa (dle Černého)[9]
| Srdeční chlopeň | Puncta maxima na těle psa |
|---|---|
| Mitrální chlopeň | Vlevo 5. mezižebří v úrovni přechodu žebra v chrupavku |
| Aortální chlopeň | Vlevo 4. mezižebří pod linií vedenou ramenním kloubem |
| Trojcípá chlopeň | Vpravo 4. mezižebří v úrovni přechodu žebra v chrupavku |
| Poloměsíčítá chlopeň | Vlevo 3 mezižebří pod punct. max. aortální chlopně |
Srdeční činnost psa je stejně jako u člověka řízena převodním systémem, který generuje vzruchy, které se dále po převodních drahách šíří srdeční svalovinou a jsou příčinou koordinované srdeční činnosti. Impuls k srdeční činnosti u zdravého psa vychází ze sinoatriálního uzlu, výsledný rytmus se nazývá sinusový. U psa je normální srdeční frekvence v klidu je 60-160/min,[10] přičemž u malých psů je srdeční frekvence vyšší než u psů většího vzrůstu. U plodů a štěňat je srdeční frekvence vyšší, 180-200/min,[11] a pokles srdeční frekvence pod 170/min je již příznakem tísně plodu.[11] Běžná je přitom mírně nepravidelná srdeční akce, při nádechu se srdeční akce mírně zrychluje a při výdechu naopak zpomaluje. Tento typ arytmie se nazývá sinusová respirační arytmie, je u psa v klidu naprosto fyziologickým stavem a při zátěži a výslednému zvýšení srdeční frekvence zmizí.[10][12] Podstatou jevu je Bainbridgeův reflex a aktivace sympatiku v závislosti na plnění velkých žil, na které má mimo jiné vliv i rozdílný tlak v hrudníku během nádechu a výdechu.
Tepový objem srdce společně s poddajností stěny velkých tepen ovlivňuje hodnotu systolického krevního tlaku, diastolický krevní tlak je ovlivňovaný více minutovým objemem krve přečerpané srdcem a periferním odporem krevního oběhu. Průměrný krevní tlak psa je 120/70 mmHg,[13] ale fyziologická hodnota velice závisí na plemeni vyšetřovaného psa, obří plemena nebo zlatí retrívři mají krevní tlak nižší, naopak chrti vyšší než je průměrná hodnota.[14] Rutinně se krevní tlak psa neměří, není ho možné změřit rtuťovým tonometrem, používají se sonografické metody založené na Dopplerově jevu a oscilometr.[13]
Stejně jako u člověka je i u psa tlak krve řízen centrálně ovlivňováním průsvitu cév, především arteriol. Rychlý účinek na zvýšení krevního tlaku má adrenalin a noradrenalin, pomaleji ale dlouhodoběji je krevní tlak řízen renin-angiotenzinovým systémem, na kterém se podílejí ledviny a také sympatikus produkcí hormonu aldosteronu. Konečným důsledkem je kromě vazokonstrikce také omezení tvorby moči, zadržování tekutiny a zvýšení cirkulujícího objemu.
Místní autoregulační mechanismy dále určují distribuci krve do jednotlivých částí těla. Při snížení obsahu kyslíku v nějaké části těla se místní cévy rozšíří, umožní přítok většího množství krve a situace se normalizuje. V klidu je každá část těla zásobována poměrně stálým procentuálním podílem srdečního výdeje, to se ale může změnit v závislosti na situaci: v klidu přitéká do kosterních svalů 20-25% srdečního výdeje, ale během fyzické aktivity to může být až 75%.[13]
Tělní tekutiny
Hlavní extracelulární tekutinou je krev. Psí krev se složením ani funkcí významně neodlišuje od krve ostatních savců: skládá se z tekuté složky, krevní plazmy, a krevních buněk. Krev je transportní medium, zajišťuje transport plynů, metabolitů, hormonů, iontů i tepla a vytváří a udržuje stálé vnitřní prostředí v organismu. Dospělý pes má asi 80 ml/kg krve[15] a krevní buňky tvoří 37-55% jejího objemu.[16]
Červené krvinky psa jsou bezjaderné buňky bikonkávního tvaru. Jejich průměr je okolo 7 μm, jsou tedy podobně velké jako krvinky člověka a mírně větší než červené krvinky kočky.[17] Pod světelným mikroskopem je dobře patrné centrální projasnění, které zabírá 1/3 buňky[17] a běžná je mírná anizocytóza - ne všechny krvinky jsou přesně stejně velké. V jednom litru krve jich je 5,5 - 8,5×1012,[16] jejich průměrný objem (MCV) je 65-75 fl a každá obsahuje 22-25 pg hemoglobinu.[16] Hlavní funkcí červených krvinek je transport dýchacích plynů - kyslíku a oxidu uhličitého.
Bílé krvinky jsou buňky, které se podílejí na fungování imunitního systému organismu. V krvi dospělého zdravého psa je nachází 6 - 17×109 bílých krvinek v jednom litru.[16] Existuje několik druhů bílých krvinek, lišících se velikostí, uspořádáním buňky i funkcí. Nejpočetnější bílou krvinkou v krvi psa je neutrofilní granulocyt. Tyto buňky slouží jako pěšáci imunitního systému, hrají úlohu v nespecifické imunitě a při akutních zánětech. Zralých neutrofilních granulocytů se segmentovaným jádrem je v krvi 3,3 - 10,5×109/l. Dalším početněji zastoupenou bílou krvinkou jsou lymfocyty, jejich množství se pohybuje mezi 1,0 - 3,6×109/l. Lymfocyty jsou obecně buňky zajišťující specifickou imunitu s tvorbou protilátek.
Zástava krvácení je společně se srážecími faktory přítomnými v krevní plazmě zprostředkována krevními destičkami. Jsou to malé, bezjaderné buňky oválného nebo tyčinkovitého tvaru, velikosti 1/4 - 2/3 červené krvinky.[17]. Obsahují granula, která se uvolňují při srážení krve. Aktivované krevní destičky mají tenké panožky a shlukují se za vzniku bílého trombu, což je první, prozatímní uzávěr porušené cévy, který brání další ztrátě krve.
Fyziologie trávení

Trávicí soustava psa je složená z dutiny ústní, hltanu, jícnu, žaludku a střev. Její hlavní funkce je zajistit přívod živin.
Dutina ústní je uzpůsobena především ke získávání potravy, méně již k jejímu zpracování. Řezáky slouží jen k samotnému uchopení potravy (kořisti), ne k jejímu dělení na jednotlivá sousta. Vzhledem ke tvaru čelistí, zubů i samotné dutiny ústní pes přijatou potravu nemůže drtit, ale jeho třenové stoličky a stoličky do sebe zapadají jako nůžky a umožňují stříhání potravy včetně tvrdých kostí na kusy, které mohou být dále spolknuty vcelku.[18] Pes má mohutné žvýkací svaly a silné čelisti, což mu umožňuje dosáhnout síly skusu potřebné k drcení kostí. Přesné určení síly skusu psa je ale poměrně obtížné a ke studiu bylo použito několik přístupů – u živých psů v anestezii se žvýkací svaly stimulovaly elektrickým proudem a měřila se takto získaná výsledná síla skusu,[19][20] síla skusu se určovala měřením psích lebek a regresní analýzou takto získaných údajů,[21] nebo psi kousali na rukáv opatřený senzory, které měřily sílu skusu.[22] Vyvinutý tlak záleží na tom, kde v čelisti je síla skusu měřena, změřená síla skusu na úrovni špičáku se u pokusných psů (kříženců různých velikostí) pohybovala od 147 do 946 N,[19] na úrovni druhé stoličky činil změřený tlak již 524 – 3417 N.[19] U jednotlivých psů síla skusu koreluje s velikostí psa, čím větší pes, tím silnější skus,[21] a u středních a velkých psů má díky působení páky na výslednou sílu skusu vliv také poměr délky a šířky čelistí.[21] Při měření síly zákusu na rukáv u různě velkých psů byl změřený tlak průměrně 256 N, přičemž medián byl 163 N.[22]
Při příjmu potravy se pes orientuje především čichem,[18] má sice chuťové pohárky, ale ve srovnání s člověkem jen asi 1/6 celkového množství.[23] Jazyk samotný také nemá při příjmu potravy takový význam jako u člověka, potravu jen přidržuje, ale v podstatě neposunuje směrem dozadu.[18] Jazyk se uplatňuje při pití: pes nemá pysky schopné pití v doušcích, vodu vmetává rychlými pohyby jazyka směrem k hltanu, kde ji po nahromadění většího množství spolkne.
Sliny slouží při příjmu potravy především ke zvlhčování soust. Produkce slin začíná už jakmile pes vidí nebo cítí potravu[24] a dá se vyvolat i vytvořeným podmíněným reflexem. Množství slin vyloučených za den se pohybuje mezi 0,1 - 0,15 l,[25], pH psích slin je 7,34 - 7,8 a osmolalita je stejná nebo nižší než v krevní plazmě.[24]. Psí sliny jsou tvořeny především vodou, dále sodnými, draselnými, vápenatými a chloridovými ionty, bikarbonátem, hlenem a enzymy, jako je lysozym s antibakteriálním účinkem. Psí sliny neobsahují žádnou α-amylázu.[24] Pes krmený masem vylučuje sliny spíše hlenovité, zatímco pes přijímající suché granulované krmivo tvoří sliny řídké a vodnaté.[25]
Zatímco při příjmu potravy mají sliny jen lubrikační funkci, jsou velmi důležité při termoregulaci psa při tzv. evaporativním ochlazování. Je-li to nutné, může pes vylučovat sliny 10x rychleji než člověk (na gram tkáně slinné žlázy),[26] tvořené sliny jsou vodnaté a izotonické a jejich vypařování z povrchu jazyka a sliznice dutiny ústní slouží k ochlazování těla psa stejně, jako vypařování potu z povrchu kůže ochlazuje tělo lidské. Parasympatikem ovlivňovaná produkce slin je příčinou zvýšeného slinění při nadměrném stresu nebo nevolnosti.
Příjem potravy je hltavý a velice rychlý. Jedná se o atavismus zděděný po vlčím předkovi. Při polykání dochází ke koordinovaným stahům svalů jazyka, jazylky, hltanu, jícnu i bránice. Jícen psa je snadno roztažitelný, jeho svalová vrstva je tvořena jen příčně pruhovanou svalovinou a průchod perilstatických vln je proto velmi rychlý,[26] sousto se dostane do žaludku během přibližně 5 sekund.[27]
Žaludek psa je velký a roztažitelný vakovitý orgán. Tvoří 62% celkového objemu trávicího traktu.[27] V žaludku začíná vlastní proces trávení, zvláště bílkovin a tuků, žaludek slouží též jako rezervoár potravy. Pes je přizpůsobený k příjmu většího množství potravy v delších časových intervalech a velikost žaludku tomu odpovídá.
Žlázy v žaludeční sliznici vylučují pepsin, proteolytický enzym, který má největší aktivitu při pH 2, a žaludeční lipázu, enzym štěpící triacylglyceroly při pH 4.[26] Celkový podíl žaludeční lipázy na trávení tuků je ale malý.[26] K zajištění nízkého pH je do žaludeční šťávy vylučována kyselina chlorovodíková. Pes má jen velmi nízkou bazální sekreci kyseliny, proto celkové pH v psím žaludku kolísá od 1 - 6 ve vztahu k příjmu potravy, průměrné pH je 3 - 4.[28] U nenakrmeného psa je žaludeční pH poměrně stálé a může odpovídat pH ve dvanáctníku.[29] Pokles pH po příjmu potravy závisí na typu přijatého krmení a jeho pufrační kapacitě - po příjmu tekuté kompletní stravy pokleslo pH žaludečního obsahu pod 4 již za 10 - 20 minut, po nakrmení masem kleslo za hodinu na méně než 6.[26] Během trávení potravy klesá pH až na hodnotu 2,1.[29] a maximální sekrece HCl dosahuje 39 mEq/h, přičemž u člověka je to maximálně 23 mEq/h.[29]
Celková produkce žaludečních šťáv je ovlivňována množstvím přijaté potravy a jejím obsahem bílkovin,[26] a je hormonálně řízená - gastrin a ACTH zvyšují produkci kyseliny solné v žaludku, sekretin naopak vede ke snížení produkce (potlačením působení gastrinu). 24 kg vážící pes je schopný za 3,5 hodiny vyloučit 1 litr žaludeční šťávy.[30]
Perilstatika žaludku napomáhá trávení promícháváním a drcením jeho obsahu. Žaludek se vyprazdňuje postupně a doba, za kterou se trávenina přesune do dvanáctníku, záleží na typu přijaté potravy - nejrychlejší je u syrové stravy, pomalejší u konzervované stravy a nejpomalejší po příjmu granulí,[31] tekutiny z žaludku odcházejí nejrychleji. Vliv má též složení krmiva - to s obsahem masa se vyprazdňuje pomaleji než krmení na rostlinné bázi - obsah vlákniny, tuku, tryptofanu, jeho acidita a osmolalita, přístup psa k vodě a individuální rozdíly mezi zvířaty. Poločas vyprazdňování žaludku se proto průměrně pohybuje mezi 72 - 240 minutami.[32]
Naprostá většina přijatých živin se vstřebává v tenkém střevě.
Do dvanáctníku ústí vývody slinivky břišní, která do střeva vylučuje pankreatickou lipázu k trávení tuků, chymotrypsin, což je proteáza štěpící bílkoviny, a pankreatickou amylázu, která štěpí škroby. Poměr vylučovaných trávicích enzymů se může přizpůsobit přijímané potravě, krmení bohaté na bílkoviny vede k většímu vylučování chymotripsinu, krmiva s přídavkem pšeničných otrub zvyšují vylučování amylázy.[31] Pankreatická šťáva má pH pH je 7,1 - 8,2.[33], kromě enzymů obsahuje také velké množství bikarbonátu, který neutralizuje kyselý obsah žaludku, a antibakteriálně působící protein, který potlačuje růst bakterií E.coli, Salmonella, Shigella a Klebsiella.[33]
Další žlázou, která se podílí na trávení střevního obsahu, jsou játra. Žluč se v játrech tvoří neustále, ale u hladového psa je 29 to 53%[31] ukládáno ve žlučníku, kde se dále koncentruje a do dvanáctníku je vypuzována až po nakrmení. Žluč obsahuje bilirubin a žlučové kyseliny, které jsou u psa z 99 % konjugovány, a to s taurinem ve formě kyseliny tautocholové, kyseliny taurodeoxycholové a kyseliny taurochenodeoxycholové.[31] Žlučové kyseliny slouží jako emulgátory tuků, rozbíjejí tukové kapénky na menší, dostupnější trávení a vstřebávání střevní sliznicí.
Vylučování pankreatické šťávy a žluče je spuštěno sekretinem, který tvoří buňky sliznice dvanáctníku při detekci kyselého obsahu uvnitř střeva, a cholecystokininem, který je produkovaný při přítomnosti nestrávené potravy, zvláště tuků a peptidů.[33]
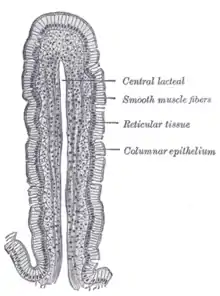
Promícháním žaludečního obsahu s pankreatickou šťávou a žlučí vznikne trávenina, s průměrným pH ve dvanáctníku kolem 6,2[32] a průměrným pH ve střevě celkem 7,3[31] Střevní výstelka tvoří prstovité výběžky, klky, které zvětšují absorpční povrch střeva. Cytoplazmatická membrána střevních buněk, která směřuje do dutiny střeva, je též zprohýbaná a vytváří mikroklky, neboli kartáčový lem. Na kartáčovém lemu pokračuje trávení, nachází se zde další enzymy, jako je glykosidáza,[29] která štěpí oligosacharidy na jednoduché cukry, několik druhů peptidáz (např. aminopeptidázy, enteropeptidázu, endopeptidázu)[29] a fosfatázy, mimo jiné i alkalická fosfatáza. Po rozložení živin na monosacharidy, aminokyseliny a malé kapénky tuků jsou tyto absorbovány skrze střevní sliznici do krve (v případě tuků do lymfy jako chylomikrony). Ze střeva se aktivně vstřebávají také soli, v případě sodíku a draslíku se vstřebává více než 90 % jejich obsahu v trávenině,[31] a voda - v tenkém střevě se voda vstřebává v množství odpovídající 15-65 ml/kg živé hmotnosti,[31] takže pes o hmotnosti 20 kg absorbuje za den přes 3 l tekutiny, z toho je 50 % vstřebáno v lačníku a 40% v kyčelníku.[32] Doba tranzitu střevem je přímo úměrná době vyprazdňování žaludku[31], je ovlivňována složením potravy, množstvím vlákniny v trávenině i působením nervového systému. Peristaltické vlny posouvají tráveninu lačníkem průměrnou rychlostí 5 cm/min a kyčelníkem 15 cm/min.[34]; délka tenkého střeva u psa o hmotnosti 20 kg se rovná 340 cm, doba tranzitu tenkým střevem je proto rovna přibližně 1 hodině.[34] 95% procent tráveniny dosáhne terminálního konce kyčelníku po 12 hodinách.[34] U psů, jejichž doba tranzitu tenkým střevem byla měřena pomocí orálně podaného barya, prošla veškerá kontrastní látka z dvanáctníku do tlustého střeva během 214 +/- 25 minut.[31] Tekutiny samotné naproti tomu procházejí tenkým střevem velice rychle, značený fyziologický roztok se dostal do slepého střeva už za 37 minut.[34]

V tlustém střevě sice dochází k dalšímu trávení živin, ale pro psa samotného to má již malý význam. Slepé střevo psa je velmi malé, tvoří jen 1 % objemu trávicího traktu, tlusté střevo 13 %.[29] V tlustém střevě se vstřebává poslední přebytečná voda a elektrolyty a bohatá bakteriální populace zajišťuje fermentaci nestravitelných živin, jako je vláknina či složité škroby. Při krmení dobře stravitelnou potravou činí podíl trávení v tlustém střevě jen 1 - 4 %[35] z celkové stravitelnosti, při krmení syrovým bramborovým škrobem nebo tapiokou i 24 %.[34] Nejdůležitějšími bakteriálními rody v tlustém střevě jsou Streptococcus, Lactobacillus, Bacteroides a druhy Clostridium perfringens a E.coli.[35] Produktem fermentace vlákniny jsou těkavé mastné kyseliny, které slouží jako důležitý zdroj energie pro buňky střevní výstelky tlustého střeva,[35] ale psí organismus jako celek je využít neumí (na rozdíl od některých býložravců). Dalšími produkty bakteriálního rozkladu je sulfan, metan, amoniak nebo indol,[35] bakterie rozkládají také bilirubin, za vzniku hnědě zbarveného urobilinogenu. Zahušťováním se trávenina postupně přeměňuje ve formované tuhé výkaly, které během defekace definitivně opouštějí řitním otvorem trávicí soustavu.
Celková doba od pozření potravy po vyloučení stolice, která z této potravy vznikla, byla měřena pomocí bezdrátové kapslové endoskopie a pohybovala se mezi 1294 a 3443 minutami.[36] Velké rozdíly jsou způsobené především dobou setrvávání výkalů v tlustém střevě, které slouží i jako jejich rezervoár.
Fyziologie vylučování a osmoregulace
K zajištění stálého vnitřního prostředí v organismu musí pes mít způsob, jak regulovat pH tělních tekutin, koncentraci solí, vápníku a fosforu a kde se zbavovat konečných produktů metabolismu, jako je močovina nebo kreatinin, a přijatých cizorodých látek.

Stejně jako člověk a ostatní savci má pes párovou ledvinu. Uvnitř ledvin se nacházejí nefrony, které tvoří její funkční jednotky. Každý nefron je tvořen klubíčkem vlásečnic, glomerulem, v Bowmanově pouzdře, kde se přitékající krev filtruje a vzniká primární moč, a kanálkem, kudy primární moč protéká a její složení je buňkami kanálku aktivně měněno, dochází k absorpci glukózy, výměně a sekreci iontů, sekreci močoviny a zahušťování moči. Definitivní moč přechází do sběracích kanálků, které ústí do ledvinné pánvičky. Odtud prochází močovodem do močového měchýře, kde je skladována a pak vymočena a odstraněna z těla. V jedné psí ledvině se nachází přes 400 000 nefronů[37] a teprve po zničení 75% z nich se začne projevovat ledvinné selhávání.[38] Zatímco u člověka tvoří nefrony s dlouhými Henleovými kličkami, které na principu protiproudového mechanismu účinně zahušťují vznikající moč, jen asi 15% všech nefronů, v psí ledvině mají dlouhou kličku všechny nefrony.[39] Psi proto mají větší schopnost koncentrovat moč než člověk: maximální osmolalita psí moči je 2,000 mOsm/l,[40], u člověka je maximum na 1200 mOsm/l moči.[41]
K udržení vodní bilance zdravý pes denně vypije 45 - 80 ml/kg vody.[42] Široké rozmezí je způsobeno velkými individuálními rozdíly v potřebě vody - spíš než na hmotnosti je spotřeba závislá na povrchu těla, malí psi pijí na svou hmotnost více než psi velcí,[42] zvířata krmená vlhkou potravou pijí méně než ta, která dostávají granule,[43] vliv má i fyzická námaha a teplota vnějšího prostředí. Bezdůvodný[p 1] příjem vody nad 90 ml/kg/24 hodin[43] je již považován za patologický.
Psí moč je čirá tekutina, různé intenzity žluté barvy. Obvykle platí, že čím je koncentrovanější, tím je tmavší. Její specifická hmotnost je pohybuje od 1001 do více než 1075 kg/m3,[44]. Specifická hmotnost pod 1008 kg/m3[45] znamená, že moč je hypostenurická, více zředěná než krevní plazma - to může být normální v případě nadměrného pití způsobeného například stresem (tzv. primární polydipsie), nebo při infuzní terapii, hypostenurická moč je produkována v případě, že nějaký patologický proces blokuje působení hormonu ADH na nefrony: může se jednat o zánět, jako v případě pyometry, selhání jater, nebo Cushingův syndrom, či podání kortikosteroidů. Centrální diabetes insipidus, kdy ADH není vůbec přítomen, podání diuretik nebo přítomnost glukózy v moči (glukosurie) je projevuje též produkcí velmi málo zředěné moči.
Isostenurická moč je taková, která má specifickou hmotnost odpovídající krevní plazmě, u psa je to 1008 – 1012 kg/m3.[45] Jednorázové naměření této hodnoty je prakticky bezvýznamné, může znamenat, že pes ztratil schopnost koncentrovat moč, ale stejně tak se může jednat o momentální normální stav, kdy vodní bilance organismu koncentraci moči nevyžaduje. Nenormální je ale isostenurická moč v kombinaci s dehydratací.
Nejčastěji zdravý pes vytváří koncentrovanou moč se specifickou hmotností 1015 – 1045 kg/m3.[44] Extrémně koncentrovaná moč se specifickou hmotností nad 1050 kg/m3.[44] už značí zhoršenou perfuzi ledvin, která nastává při hypovolémii nebo při srdečním selhání.
pH psí moči se pohybuje mezi 5,5 - 7, při vysokoproteinové dietě, při námaze či během horečky je spíše kyselá,[45] po nakrmení, při krmení potravou s velkým obsahem vlákniny a malým obsahem bílkovin bývá spíše zásaditější.[45] Moč zdravého psa je sterilní.
Obvyklá produkce moči se pohybuje mezi 24 - 41 ml/kg za den,[42] nad 45 ml/kg/den[43] je to již považováno za nadměrné počení (polyurii).
Odkazy
Poznámky
- Bez vlivu velké fyzické námahy, horka, horečky, některých léků apod.
Reference
- PETSKO, Gregory A. Sleeping dogs [online]. Genome Biology, 23.10.2003 [cit. 2012-01-11]. Dostupné online. (anglicky)
- Randy Kidd. The Canine Sense of Smell [online]. [cit. 2011-12-25]. Dostupné online. (anglicky)
- How do dogs breathe? [online]. [cit. 2012-01-14]. Dostupné v archivu pořízeném dne 2012-06-27. (anglicky)
- MILAN, Dudek. Základy veterinářství služebních psů Policie ČR [online]. Cz-pes.cz [cit. 2012-01-14]. Kapitola Základy fyziologie psa. Dostupné online. (česky)
- PROCHÁZKA, Zdeněk. Chov psů. Praha - Litomyšl: Paseka, 2005. Biologické základy chovu psů s. ISBN 80-7185-768-8. Kapitola 76, s. 320. (česky)
- SIEGWART, Benno; GEHR, Peter; GIL, Joan, et al. Morphometric estimation of pulmonary diffusion capacity: IV. The normal dog lung. S. 141–159. Respiration Physiology [online]. 11/1971 [cit. 14.1.2012]. Roč. 13, čís. 2, s. 141–159. Dostupné online. ISSN 0034-5687.
- HUNYADY, Kristen. Anesthesia Cheat Sheet [online]. University of Wisconsin - Madison: School of Veterinary Medicine [cit. 2012-01-14]. Dostupné v archivu pořízeném dne 2012-03-04. (anglicky)
- ČERNÝ, Hugo. Veterinární anatomie pro studium a praxi. Brno: Noviko a.s., 2004. 528 s. ISBN 80-86542-05-X. Kapitola kapitola 4.7.3. srdce, osrdečník, s. 166–167. (česky)
- Černý, Veterinární anatomie pro studium a praxi, kapitola 4.7.3.1 Puncta maxima, s.170
- Cardiovascular System Introduction: Introduction [online]. Merck veterinary manual [cit. 2012-01-17]. Dostupné v archivu pořízeném dne 2012-03-04. (anglicky)
- YIN, Sophia. The Small Animal Veterinary Nerdbook. third edition. vyd. Davis: CattleDog Publishing, 2010. ISBN 978-0-9641518-5-7. Kapitola Reproduction:Pregnancy, s. 19.12.. (anglicky) Dále Sophia Yin, The Small Animal Veterinary Nerdbook.
- Sophia Yin, The Small Animal Veterinary Nerdbook, kapitola Cardiology: Arrhythmias, s.3.18
- REECE, William O. Fyziologie a funkční anatomie domácích zvířat. druhé, rozšířené. vyd. Praha: Grada, 2010. 480 s. ISBN 978-80-247-3282-4. S. 227. (anglicky)
- EGNER, Beate; CARR, Anthony; BROWN, Scott. Essential facts of blood pressure in dogs and cats. UK: Lifelearn, Inc, 2003. 216 s. ISBN 3-00-011096-8. S. 11. (anglicky)
- FINSTERER, U.; PRUCKSUNAND, P.; BRECHTELSBAUER, H. Critical evaluation of methods for determination of blood volume in the dog. S. 63–72. European Journal of Physiology [online]. 27.2.1973 [cit. 21.1.2012]. Roč. 341, čís. 1, s. 63–72. Dostupné online. ISSN 1432-2013.[nedostupný zdroj]
- SVOBODA, Miroslav; SENIOR, David F.; DOUBEK, Jaroslav, a kol. Nemoci psa a kočky 1.díl. 2.. vyd. Brno: Noviko a.s., 2008. 1152 s. ISBN 978-80-86542-18-8. Kapitola Základní laboratorní diagnostika, s. 70. (česky)
- COWELL, Rick L.; TYLER, Ronald D.; MEINKOTH, James H., a kol. Diagnostic Cytology and Hematology of the Dog and Cat. 3.. vyd. Kanada: Mosby Elsevier, 2008. 475 s. ISBN 978-0-323-03422-7. Kapitola Peripheral blood smears, s. 395. (anglicky)
- Procházka, Chov psů, kapitola biologické základy chovu psů, s. 67
- ELLIS , THOMASON JJ, KEBREAB E, JL; THOMASON, JJ; KEBREAB, E, et al. Calibration of estimated biting forces in domestic canids: comparison of post-mortem and in vivo measurements. S. 769–80. Journal of Anatomy [online]. 6/2008 [cit. 1.2.2012]. Roč. 6, čís. 212, s. 769–80. Dostupné online.
- COREN, Stanley. Dog Bite Force: Myths, Misinterpretations and Realities [online]. 1469-7580: Psychology Today, 17.5.2010 [cit. 2012-02-01]. Dostupné online. (anglicky)
- ELLIS, JL; THOMASON, J.; KEBREAB, E., et al. Cranial dimensions and forces of biting in the domestic dog. S. 362–373. Journal of anatomy [online]. 3/2009 [cit. 1.2.2012]. Roč. 3, čís. 214, s. 362–373. Dostupné online. ISSN 1469-7580.
- LINDNER, DL; MARRETTA, SM; PIJANOWSKI, GJ, et al. Measurement of bite force in dogs: a pilot study. S. 49–52. Journal of Veterinary dentistry [online]. 6/1995 [cit. 1.2.2012]. Roč. 2, čís. 12, s. 49–52. Dostupné online. ISSN 0898-7564.
- General Physiology of Dogs [online]. [cit. 2011-12-22]. Dostupné v archivu pořízeném dne 03-01-2012. (anglicky)
- Subcommittee on Dog and Cat Nutrition. Nutrient requirements of dogs and cats. [s.l.]: National Academies Press, 2003. 424 s. ISBN 978-0309086288. Kapitola Comparative digestive physiology of Dogs and Cats, s. 6. (anglicky)
- HEGEWALD - KAWICH, Horst. Psí desatero. Praha: Knižní klub, 1996. 174 s. ISBN 80-7176-395-0. Kapitola Výživa, s. 112. (česky)
- Nutrient requirements of dogs and cats, s. 7
- CHIBA, Lee I. Animal Nutrition Handbook - Digestive Physiology [online]. Auburn University - College of Agriculture, 2009 [cit. 2012-02-04]. Dostupné online. (anglicky)
- BROWN, Scott Anthony. Veterinary Pharmacology and Therapeutics. 8.. vyd. Iowa: Wiley-Blackwell, 2001. 1201 s. Dostupné online. ISBN 0-8138-1743-9. Kapitola chapter 3: Pharmacokinetics: Disposition and fate of Drugs in the Body, s. 23. (anglicky)
- KARARLI, Turgul T. COMPARISON O F THE GASTROINTESTINAL ANATOMY, PHYSIOLOGY, AND BIOCHEMISTRY OF HUMANS AND COMMONLY USED LABORATORY ANIMALS. Biopharmaceutics & Drug Disposition [online]. 11.1.2006 [cit. 5.2.2012]. Roč. 6, čís. 5. PDF online.
- Procházka, Chov psů, s. 68
- SMEETS-PEETERS, TIM WATSON3, MANS MINEKUS', Marianne; WATSON, Tim; MINEKUS, Mans, et al. A review of the physiology of the canine digestive tract related to the development of in vitro systems. S. 45–69. Nutrition Research Reviews [online]. 1998 [cit. 5.2.2012]. Čís. 11, s. 45–69. Dostupné online.
- Nutrient requirements of dogs and cats, s. 8
- Nutrient requirements of dogs and cats, s. 9
- Nutrient requirements of dogs and cats, s. 10
- Nutrient requirements of dogs and cats, s. 11
- BOILLAT, Carol S.; GASCHEN, Frédéric P.; GISELLE, L. Assessment of the relationship between body weight and gastrointestinal transit times measured by use of a wireless motility capsule system in dogs. S. 898–902. American Journal of Veterinary Research [online]. 8/2010 [cit. 6.2.2012]. Roč. 71, čís. 8, s. 898–902. Dostupné online. ISSN 0002-9645.
- EISENBRANDT, David L.; PHEMISTER, Robert D. Postnatal development of the canine kidney: Quantitative and qualitative morphology. S. 179–193. American Journal of Anatomy [online]. 2/1979 [cit. 6.2.2012]. Roč. 154, čís. 2, s. 179–193. Dostupné online. ISSN 1553-0795.
- FIALA, Tomáš. Chronické selhání ledvin [online]. Praha: AA-vet, 30.6.2005 [cit. 2012-02-06]. Dostupné v archivu pořízeném dne 2011-11-28. (česky)
- J.Charles Jennette, Jean L. Olson, Melvin M. Schwartz and Fred G. Silva. Heptinstall's Pathology Of The Kidney. 6. vyd. Svazek 1. Filadelfie: Lippincott Williams and Wilkins, 2006. 1600 s. ISBN 0-7817-4750-3. Kapitola Renal anatomy and histology, s. 20. (anglicky)
- LEAF, Alexander; MAMBY, Audley R. THE NORMAL ANTIDIURETIC MECHANISM IN MAN AND DOG;ITS REGULATION BY EXTRACELLULAR FLUID TONICITY. The journal of Clinical investigation [online]. 9.8.1951 [cit. 22.5.2012]. Dostupné online.
- SHERWOOD, Lauralee. Fundamentals of Human Physiology. [s.l.]: Brooks/Cole, 2010. 710 s. ISBN 0840062257. Kapitola 14, s. 539. (anglicky)
- FENNER, William R. Quick Reference to Veterinary Medicine. 3.. vyd. USA: Wiley-Blackwell, 2000. 732 s. Dostupné online. ISBN 0-397-51608-8. Kapitola 10 Polyuria and polydipsia, s. 118. (anglicky)
- Proceeding of the NAVC North American Veterinary Conference [online]. Orlando, Florida: IVIS.org, 8.-12.1.2005 [cit. 2012-02-06]. PDF online. (anglicky)
- WATSON, ADJ. Using urine specific gravity [online]. IRIS, rev. 2010 [cit. 2012-02-06]. Dostupné v archivu pořízeném dne 2012-02-26. (anglicky)
- YIN, Sophia. The Small Animal Veterinary Nerdbook, Kapitola: Urinanalysis, s 4.27
Související články
- Anatomie psa domácího
- Exteriér psa
- Genetika zbarvení psů
- Pes domácí
Literatura
- PROCHÁZKA, Zdeněk. Chov psů. Praha - Litomyšl: Paseka, 2005. 314 s. ISBN 80-7185-768-8.
- DUDEK, Milan, a kolektiv. Základy veterinářství služebních psů Policie ČR. Praha: Ministerstvo vnitra, 1996. Dostupné online.
- TROJAN, Stanislav, a kolektiv. Lékařská fyziologie. 4., přepracované a doplněné vyd. Praha: Grada Publishing, 2003. 772 s. ISBN 80-247-0512-5.
- REECE, William O. Fyziologie a funkční anatomie domácích zvířat. 2., rozšířené vyd. Praha: Grada, 2010. 480 s. ISBN 978-80-247-3282-4.
- KUČERA, Jaroslav, a kolektiv. Nefrologie a urologie psa a kočky. Brno: Noviko, 2007. 337 s. ISBN 978-80-86542-17-1.
- SVOBODA, Miroslav; SENIOR, David F.; DOUBEK, Jaroslav, a kol. Nemoci psa a kočky 1.díl. 2. vyd. Brno: Noviko, 2008. 1152 s. ISBN 978-80-86542-18-8.