Zamrzlá evoluce
Zamrzlá evoluce je teoretickým konceptem českého evolučního biologa prof. Jaroslava Flegra. Tento koncept v sobě zahrnuje i dříve formulovanou teorii zamrzlé plasticity[1].

Teorie zamrzlé plasticity popisuje mikroevoluci pohlavních druhů[1][2][3][4][5]. Od hlavního proudu neodarwinismu se liší především tím, že předpokládá dlouhodobou stabilitu ve vlastnostech těchto druhů a silná omezení při jejich nevratném přizpůsobování prostřednictvím přirozeného výběru. K výraznějším přizpůsobením podle ní může docházet jen po určitých typech speciací (např. peripatrické speciaci) na počátku existence druhů. Po většinu doby své existence jsou podle teorie zamrzlé plasticity druhy evolučně zamrzlé a na přírodní výběr reagují jen změnami v zastoupení již přítomné genetické variability.
Teorie zamrzlé evoluce popisuje makroevoluční důsledky evolučního zamrzání pohlavních druhů postulovaného teorií zamrzlé plasticity[1][2]. Jednotlivé znaky podle této teorie nerozmrzají při vzniku nového druhu stejně snadno. V delším časovém měřítku se tak podle ní v jednotlivých evolučních liniích hromadí principem třídění z hlediska stability znaky, které nemohou rozmrznout za žádných představitelných podmínek. Evolvabilita druhů odštěpených v rámci těchto linií se podle teorie zamrzlé evoluce postupně snižuje, což může vést až k drastické ztrátě schopnosti odpovídat na selekční tlaky prostředí a tlaku na vznik symbióz, koloniality, mnohobuněčnosti či sofistikované genetické architektury znaků[1].
Zamrzlá evoluce je i název Flegrovy knihy na dané téma, která dostala v roce 2007 cenu Magnesia Litera v kategorii naučná literatura[6]. Byla přeložena do anglického jazyka a vyšla pod názvem Frozen Evolution[2].
Teorie zamrzlé plasticity
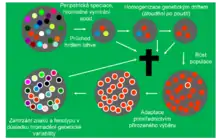
Omezená dědičnost fenotypu pohlavních druhů
Na rozdíl od nepohlavních druhů představuje fungování přirozeného výběru u pohlavních druhů teoretický problém, který podle J. Flegra nedokáže plně vysvětlit ani dnešní genocentrický pohled na evoluci reprezentovaný teorií sobeckého genu[1][2][3][4][5]. Genotyp potomka se u pohlavních organismů skládá z alel namíchaných náhodně po jedné polovině od otce a od matky. Jedna alela může ovlivňovat mnoho znaků (pleiotropie) a jeden znak je téměř vždy spoluvytvářený činností mnoha alel (epistáze). Vliv alel na fenotyp navíc není stálý, ale kontextově závislý – závisí na tom, se kterými alelami stejného genu i jiných genů se daná alela nachází v genomu jedince. Vliv jednotlivých alel na výslednou biologickou zdatnost jedince je ještě komplikovanější, často totiž navíc závisí i na frekvenci dané alely i frekvencích dalších alel v populaci. Vliv alel na biologickou zdatnost tak lze nejlépe popsat pomocí aparátu evolučně stabilních strategií. Kontextová a frekvenční závislost se navíc netýká jen vztahu mezi alelami a celkovou biologickou zdatností organismu, ale i vztahu mezi znaky jedince a jeho fitness. Dědičnost fenotypu a potažmo biologické zdatnosti je tedy u pohlavních organismů velmi malá a přirozený výběr, a to i na úrovni jednotlivých alel, u nich za normálních podmínek může fungovat jen velmi omezeně.
Přizpůsobování pohlavních druhů
Podle teorie zamrzlé plasticity nejsou pohlavní druhy za normálních podmínek schopné nevratného přizpůsobení selekčním tlakům prostředí[4][5]. Jakákoli nová alela, ať už vzniklá v důsledku mutace, nebo introdukovaná migrantem z jiné populace, se podle ní zachytí do složité sítě vzájemně provázaných vlivů alel na biologickou zdatnost. Nemůže být fixována ani eliminována (zcela vymizí zřejmě jen alely s výrazně negativním vlivem na fitness) a v populaci se udržuje v určitém rovnovážném zastoupení. Díky stejným procesům si podle ní pohlavní druhy udržují dlouhodobě vysoký genetický polymorfismus. Na momentální selekční tlaky prostředí tak podle Flegra dokáží zprvu odpovídat velmi rychle, a to pouhým posunem zastoupení alel již přítomných v populaci. Se zvětšujícím se vychýlením oproti původnímu stavu však podle něj systém klade vlivem negativních pleiotropických účinků selektovaných alel čím dál větší odpor a přizpůsobení se zpomaluje. Klíčovou roli v tomto procesu podle autora zřejmě hrají alely udržované v polymorfním stavu frekvenčně závislou selekcí a jejich funkční provázání s ostatními alelami. Po skončení selekčního tlaku se podle teorie zamrzlé plasticity frekvence alel i fenotyp druhu vrací více méně k původním hodnotám. Pohlavní druhy se tudíž podle teorie zamrzlé plasticity po většinu času své existence přizpůsobují tlakům prostředí elasticky – zpočátku snadno, se zvětšující se odchylkou od původního stavu hůře a hůře, až nakonec vůbec.
K nevratnému, plastickému, přizpůsobení pohlavních druhů může podle teorie zamrzlé plasticity docházet jen v určitých úsecích jejich existence[4][5]. Základním předpokladem je redukce genetického polymorfismu prostřednictvím fixací a eliminací alel, která vede k tomu, že se nová alela v každé generaci nachází v sousedství stejných alel. Takováto redukce nastává zejména při peripatrické speciaci, třeba při kolonizaci ostrova malým počtem jedinců, ale podobný vliv by podle Flegra mohlo mít např. i rychlé, náhodné a drastické vymření zástupců daného druhu na většině původního areálu. Obě situace doprovází tzv. efekt zakladatele, v jehož důsledku přejde do nové populace jen málo alel z původního druhu a v důsledku náhody mohou vymizet i některé alely původně velmi hojné. Zbylý genetický polymorfismus, zejména alely udržované frekvenčně závislým výběrem, ale podle teorie zamrzlé plasticity může vymizet jen působením náhody, tj. genetického driftu a příbuzenského křížení, při tzv. bloudění po poušti – mnoha generacích přežívání v malém počtu jedinců, ve kterém nerozhoduje o přežití jedince žádný typ přirozeného výběru, ale pouze náhoda. Pokud nový druh ve stavu vzniku při těchto dramatických událostech nevymře, může dojít souhrou náhodných příznivých okolností ke zvětšení jeho populace. V početné populaci se začne uplatňovat přirozený výběr. Nové alely mají na jednotném genetickém pozadí vždy stejný vliv na zdatnost jedince a druh se podle teorie zamrzlé plasticity může začít plasticky přizpůsobovat novým podmínkám. Ovšem jen do doby, než se znovu nahromadí genetický polymorfismus, zejména alely, jejichž pozitivní vliv na fitness závisí negativně na jejich frekvenci v populaci, a nový, selekcí pozměněný, druh tak přejde do elastické fáze své existence.
Doklady a testování teorie
J. Flegr uvádí na podporu teorie řadu dokladů z oborů genetiky, populační genetiky, ekologie, šlechtitelství, paleontologie a paleobiologie[1][2][3][5][7][8]. Některé ze zmíněných fenoménů je podle autorových slov možné s většími či menšími obtížemi vysvětlit i dílčími hypotézami v rámci hlavního proudu neodarwinismu. Teorie zamrzlé plasticity však všechna uvedená pozorování umožňuje organicky spojit a vysvětlit prostřednictvím jednoho teoretického konceptu, nikoli mnoha jednotlivých ad hoc hypotéz[1]. Model odvozený z teorie zamrzlé plasticity nabízí jedno z vysvětlení značného rozšíření altruistického chování[9].
O přímé testování ekologických implikací teorie zamrzlé plasticity se pokusil J. Toman[10][11]. Výsledky jeho srovnání ekologie starobyle nepohlavních a blízce příbuzných pohlavních linií eukaryot jsou ve vztahu k teorii zamrzlé plasticity ambivalentní. Spíše v její prospěch mluví preference studovaných nepohlavních linií pro abioticky stabilní prostorově homogenní podmínky s malým množstvím konkurentů, predátorů a parazitů včetně podmínek extrémních; srovnávané pohlavní linie naopak preferovaly prostředí bioticky a abioticky heterogennější, zvýhodňující rychlé evoluční reakce na úkor pomalejšího nevratného přizpůsobení. Nepotvrdila se však teorií zamrzlé plasticity předpovídaná tendence nepohlavních linií k větší přizpůsobivosti, v tomto případě tendence celých linií k obývání širšího rozmezí podmínek (konkrétně teplot) v porovnání s příbuznými pohlavními liniemi; srovnávané skupiny se v tomto ohledu statisticky významně nelišily.
Další možná uplatnění konceptu zamrzlé evoluce v ochraně přírody, technice, politologii a etice zmínil J. Flegr v knize Evoluční tání[1].
Podobné koncepty
Autor uvádí jako hlavní inspirační zdroje teorie zamrzlé plasticity teorii přerušovaných rovnováh N. Eldredge a S. J. Goulda, teorii evolučně stabilních strategií J. Maynarda Smithe a teorii mezialelické selekce W. Hamiltona rozpracovanou následně R. Dawkinsem jako teorie sobeckého genu[1][2][3].
Teorie zamrzlé plasticity předpokládá plynulý, gradualistický, průběh evoluce nepohlavních druhů a nerovnoměrný, punktuacionalistický, průběh evoluce druhů pohlavních. Punktuacionalistický charakter evoluce mnohobuněčných druhů popsali v 70. letech 20. století coby model přerušovaných rovnováh N. Eldredge a S. J. Gould a navrhli pro něj postupně řadu alternativních vysvětlení[12]. Další modely vysvětlující punktuacionalistický průběh evoluce představují teorie posunujících se rovnováh S. Wrighta, Founder-flush model H. L. Carsona, model genetické transilience A. Templetona a model genetické revoluce E. Mayra[8]. Teorie zamrzlé plasticity předpokládá, že příčinou punktuacionalistického průběhu evoluce u pohlavních druhů je střídání krátké evolučně rozmrzlé plastické fáze a dlouhé evolučně zamrzlé elastické fáze během doby existence každého pohlavního druhu, které se ve fosilním záznamu nezbytně projeví jako nerovnoměrná rychlost evoluce. Teorii zamrzlé evoluce se nejvíce blíží Mayrův model genetické revoluce, který rovněž zdůrazňuje roli efektu zakladatele[1]. Tento efekt často vede k náhodné a mnohdy drastické změně ve frekvenci jednotlivých alel, které jsou v původní populaci udržovány ve stabilní frekvenci v důsledku existence husté sítě vzájemných interakcí. E. Mayr tvrdí, že ve velké polymorfní populaci nemůže probíhat efektivní adaptivní evoluce proto, že tu nevítězí nejlepší alela, ale alela, která je schopná obstojně fungovat v nejširším spektru genotypů[1]. Model založený na Mayrově genetické revoluci původně preferovali jakožto vysvětlení punktuacionalistického průběhu evoluce i autoři konceptu přerušovaných rovnováh[12]. E. Mayr, který ještě neměl k dispozici teorii evolučně stabilních strategií, ovšem neuvažuje ústřední roli alel s frekvenčně závislým vlivem na fitness a pleiotropních efektů těchto alel na zamrzání (zelastičtění) pohlavních druhů, klíčovou úlohu snížení efektivity selekce během fáze bloudění po poušti ani úlohu následné expanze populace pro rozmrznutí (zplastičtění) nově odštěpeného druhu[1]. Rovněž neuvažuje možnost nevratného zamrzání znaků a jejich skupin postulovanou teorií zamrzlé evoluce[1].
Teorie zamrzlé evoluce
Nevratné zamrzání znaků
Jednotlivé znaky se podle teorie zamrzlé evoluce liší v tom, jak snadno při procesech popisovaných teorií zamrzlé plasticity přecházejí z elastického do plastického stavu[1][2][3][5][8]. Zatímco některým stačí k tomu, aby mohly začít nevratně odpovídat na selekční tlaky, jen malá redukce genetického polymorfismu, jiné stejného stavu dosáhnou až po velmi drastické redukci genetického polymorfismu spojené s efektem zakladatele a dlouhým přežíváním ve velmi malém počtu jedinců. Výjimečně se podle teorie zamrzlé evoluce mohou utvořit i znaky či jejich funkčně provázané skupiny (moduly), které by k úspěšnému rozmrznutí vyžadovaly přežívání v extrémně malém počtu jedinců po nerealisticky dlouhou dobu, tj. podmínky, které by vedly zcela jistě či s velmi vysokou pravděpodobností k vymření celé populace. Takové znaky či jejich skupiny jsou podle ní evolučně zamrzlé v podstatě nevratně. Znaky, které za realistických podmínek nemohou odpovídat na selekci, navíc podle Flegra mohou zřejmě vznikat i jinak. Může se jednat např. o znaky kódované mnoha redundantními geny se zastupitelnou funkcí[5][8]. Podle teorie zamrzlé evoluce se průběhu existence dané evoluční linie pohlavních organismů tyto znaky a jejich skupiny nevratně hromadí, a to prostřednictvím třídění z hlediska stability (tj. velmi obecným procesem, který lze shrnout tak, že nestabilní systémy přicházejí a odcházejí, zatímco stabilní přetrvávají a hromadí se). Evolvabilita dané linie se tak ireverzibilně snižuje[1].
Důsledky snižování evolvability pohlavních linií
Hlavním důsledkem snižování evolvability pohlavních linií předpovězeným teorií zamrzlé evoluce je podle ní ubývání jejich disparity (tvarové a funkční mezidruhové rozmanitosti)[1]. V průběhu času náhodně vymírají v rámci dané linie skupiny odvětvené od společného předka dříve i později. Nové druhy sice stále vznikají a rozrůzňují se, ale podle teorie zamrzlé evoluce se později odštěpené skupiny mohou výrazněji lišit jen ve stále omezenějším okruhu znaků. Výjimečně podle ní sice může dojít k rozmrznutí některého silně zamrzlého znaku či skupiny znaků, statisticky se ale zřejmě uplatňuje trend k nevratné ztrátě disparity.
V dlouhých časových měřítkách může postupná ztráta evolvability vytvářet sílící tlak na alternativní řešení problému s přizpůsobováním. Těmito alternativními řešeními mohou být podle J. Flegra[1] a J. Tomana[13] symbiózy a symbiogeneze, přechody ke koloniálnímu způsobu života až mnohobuněčnosti či vznik modulární tělní a genetické architektury. Společným motivem těchto řešení je postup na novou úroveň hierarchické komplexity, spojení dvou či více entit nižší úrovně stejného či různého typu a jejich společná regulace. Na nové úrovni však podle teorie zamrzlé evoluce začne hromadění nevratně zamrzlých znaků a jejich skupin nanovo a celý cyklus se opakuje. Tlak ze strany nevratného zamrzání znaků tak podle teorie zamrzlé evoluce sehrál esenciální roli při nárůstu hierarchické komplexity pozemského života.
Doklady a testování teorie
J. Flegr uvádí na podporu teorie řadu dokladů z oborů paleontologie, paleobiologie a fylogeneze[1]. Předpovědím teorie, tj. průběžnému snižování evolvability znaků, explicitně odpovídají závěry některých konkrétních paleontologických výzkumů[14].
Další možná uplatnění konceptu zamrzlé evoluce v ochraně přírody, technice, politologii a etice zmínil J. Flegr v knize Evoluční tání[1].
Podobné koncepty
Teorie zamrzlé evoluce předpokládá, že v dlouhodobém, makroevolučním, měřítku, dochází k zamrzání jednotlivých znaků a v důsledku toho i celých evolučních větví. Příčinou tohoto makroevolučního zamrzání druhů je podle J. Flegra[1] třídění z hlediska stability – znaky, které se mohou u nově vznikajících druhů měnit, přicházejí a odcházejí (resp. se mění v něco jiného), znaky, které se měnit nemohou, zůstávají a postupně se hromadí. Fenomén zamrzání evolučních větví, doprovázený snižováním disparity, popsala řada paleontologů. Postupné snižování disparity ve fosilním záznamu mnohobuněčných živočichů a dalších taxonomických skupin popsal a prostřednictvím konceptu historické kontingence vysvětloval např. S. J. Gould[15]. Přímou souvislost s pohlavním rozmnožováním a konkrétní mechanismus, v daném případě založený na hypotetické roli meiozy a semi-meiózy, pro něj však zřejmě navrhl pouze J. A. Davison[16]. Ten ve své poněkud excentrické hypotéze považuje meiózu za odvozený proces zabraňující výraznějším adaptivním změnám a limitující evoluci na generování drobných změn. Klíčovou důležitost v adaptivní evoluci přisuzuje hypotetické semi-meióze, z níž podle jeho názoru byla meióza několikrát nezávisle na sobě odvozena[16].
Bylo opakovaně navrženo, že pohlavní rozmnožování může sloužit spíše k omezení generování nežádoucí variability než k vytváření variability žádoucí[17][18][19]. Radikálnější myšlenka pohlavního rozmnožování jakožto procesu vedoucího ve svém důsledku k silnému omezení adaptivní evoluce je však daleko vzácnější a dříve se vyskytla pravděpodobně jen v již zmíněné hypotéze J. A. Davisona[16].
Přijetí a kritika
Přijetí teorie zamrzlé plasticity sahá od razantního odmítnutí, přes kritiku kvůli nedostatečnému podložení některých předpokladů, neoriginalitě či nadbytečnosti teorie až k pozitivní recepci konceptu jakožto slibné a silné hypotézy.
Pozitivního přijetí se teorie zamrzlé plasticity dočkala zejména v paleontologické a paleobiologické komunitě[20][21]. Několik autorů z oboru biosémiotiky upozornilo na zajímavé analogie mezi principy a predikcemi teorie zamrzlé plasticity a evolucí znakových systémů[22][23][24]. Kladně vyznívají i recenze anglického překladu knihy Zamrzlá evoluce (Frozen Evolution)[2] od B. Halla[25], J. Zimy[26] a C. S. Reeveové[27]. Teorii zamrzlé plasticity pokládají tito autoři za slibnou a knihu za kvalitní a hodnou pozornosti, C. S. Reeveová se vymezuje pouze proti formální stránce výkladu[27].
Publikované recenze autorových odborných článků k tématu teorie zamrzlé plasticity v anglickém jazyce[5][8] sahají od zásadní kritiky až k častějšímu pozitivnímu přijetí s několika konstruktivními návrhy. Český evoluční biolog J. Zrzavý kritizoval zejména přemrštěné ambice teorie zamrzlé plasticity. Nevyhranil se ani tak proti možné faktické správnosti konceptu, jako spíše proti jeho nedostatečné empirické podpoře a významu, když jej glosoval jako "poznámku pod čarou v učebnici evoluční biologie"[28]. L. Kratochvíl označil ve své přednášce[29] teorii za do jisté míry nadbytečnou, protože k přizpůsobování pohlavních druhů může z jeho pohledu docházet i řadou jiných způsobů. Teorie bývá vlažně kritizována i ze strany příznivců evolučně vývojové biologie (evo-devo), a to naopak pro svou přílišnou závislost na neodarwinistickém rozvrhu evoluce[30].
Zjednodušenou verzi teorie představenou v popularizační knize Zamrzlá evoluce[3] silně kritizovali z faktického, formálního i metodologického hlediska T. Grim a V. Remeš v knižní recenzi v časopisu Vesmír[31]. Jejich hlavní argumenty směřovaly na zpochybnění samotného punktuacionalistického průběhu evoluce, přičemž se opřeli o celkovou nedostatečnost fosilního záznamu pro vyvozování dalekosáhlých evolučních závěrů. Dále operovali omezeností frekvenčně závislého vlivu alel na fitness, vysokou dědivostí mnoha znaků, neoriginalitou teorie v porovnání s ostatními koncepty punktuacionalistického průběhu evoluce, nedostatečným podložením některých předpokladů, přespříliš vágními a zjednodušenými formulacemi, špatnou prací s důkazním aparátem, spekulativností, nevhodným způsobem prezentace a údajnými nepřijatelnými argumentačními triky autora. Problémy s přizpůsobivostí pohlavních druhů označili za iluzorní a neviděli nutnost formulovat novou teorii alternativní neodarwinismu. Flegrova obhajoba byla otisknuta v témže čísle Vesmíru[32]. Kritická recenze vyvolala bouřlivou diskuzi mezi českými biology a paleontology v důsledku přesahující problematiku teorie zamrzlé plasticity[20][21][31][32][33]. Výrazně kritický pohled na teorii zamrzlé plasticity z pozice neodarwinismu[zdroj?!][pozn. 1] a scientismu[zdroj?!][pozn. 2] představil v recenzi knihy Zamrzlá evoluce[3] J. Heřt[34]. Po kladném hodnocení první, úvodní, poloviny knihy silně kritizuje Flegrovu metodologii objevu a obhajoby teorie zamrzlé plasticity, novou teorii samotnou i způsob jejího představení[zdroj?!][pozn. 3]. Teorii za nesmyslnou a knihu Zamrzlá evoluce v anglickém překladu (Frozen evolution)[2] za brak označil ve své recenzi D. Graur[35].
S faktickými námitkami svých oponentů se autor pokusil vyrovnat v knize Evoluční tání[1], kde rovněž projasnil některé body, které u odborné veřejnosti vyvolávaly nepochopení, a ve větším detailu představil makroevoluční implikace teorie zamrzlé plasticity, tj. teorii zamrzlé evoluce. Kniha samotná se dočkala pozitivních recenzí v časopisech Vesmír[36] a Respekt[37]. Na témata blízká konceptu zamrzlé evoluce včetně teorie zamrzlé plasticity autor přednášel i na několika mezinárodních konferencích[38][39][40]. Rozpracování a popularizaci myšlenek teorií zamrzlé plasticity a zamrzlé evoluce se věnuje také student J. Flegra J. Toman[13][36][41][42][43][44].
Českou laickou veřejností byla teorie zamrzlé plasticity vřele přijata a zejména v souvislosti s vydáním knihy Zamrzlá evoluce[3] se dočkala značného mediálního pokrytí[45][46][47][48][49][50]. To platí v menší míře[37] také o teorii zamrzlé evoluce detailně představené v knize Evoluční tání[1]. Kniha Zamrzlá evoluce[3] získala v roce 2006 cenu Magnesia Litera v kategorii naučná literatura[6]. K tématu proběhlo i několik veřejných přednášek[43][44][51][52][53] a diskuzí[28][33][54].
Poznámky
- Viz např.:
- "Paleontologická fakta však takovou dvoufázovost evoluce jako obecně platný jev neprokazují. Bylo už uskutečněno množství studií na toto téma s různým výsledkem. Některé nacházejí plynulý, postupný vývoj druhu, jiné naznačují, že se některé druhy skutečně objevují náhle a pak zůstávají prakticky trvale neměnné. Jenže to nelze tvrdit s jistotou. Neměnná je kostra, nic víc paleontolog nemá zpravidla k dispozici. Výrazné evoluční změny ve stavbě a vzhledu měkkých tkání zastihnout nelze."
- "Stejně tak jako nefungují fakta, tak nefungují ani spekulativní Flegrovy úvahy o nemožnosti prosazení nových alel v rámci složitých mezigenových vazeb. Jestliže vznikne nová dominantní alela, prosadí se stejně dobře ve velké i malé populaci. Jistěže se ve velké populaci šíří pomaleji, ale vzniká zde zase úměrně větší množství nových mutací a rychleji se obohacuje genofond druhu. A to, že se vliv nové alely nemůže prosadit a že dokonce její vliv na vznik znaku může být podle podmínek různý až i opačný, bylo sice v ojedinělých případech pozorováno, ale jako obecný jev to lze vyloučit."
- "Gould později své výroky o jejich nové „obecné teorii, která měla nahradit darwinismus“ odvolal a prohlásil se za přesvědčeného darwinistu."
- "Základním mechanismem evoluce (kromě série dalších mechanismů, které jsou součástí neodarwinismu) je přirozený výběr účelných znaku a alel jejich genů, a jestli probíhá rychleji nebo pomaleji, nebo jen v určitém období, to je věc vedlejší."
- Viz např.:
- "Útoky na darwinismus spolu s propagováním nových protidarwinistických postmoderních evolučních teorií, jako byl např. strukturalismus, se objevovaly i na stránkách časopisu AV ČR Vesmír."
- "A patří vůbec do popularizační knihy diskuse nebo spíše přesvědčování laické veřejnosti o správnosti zcela hypotetické koncepce? A uvědomil si Flegr, že svou knihou, resp. svými výroky nahrává kreacionistům? Svědci Jehovovi, „vědečtí kreacionisté“ i dnešní množící se zastánci „inteligentního designu“ mezi našimi postmoderními intelektuály, ti všichni se na jeho knihu budou s velkým halasem odvolávat. „Vidíte, i velký evoluční biolog....atd."
- "A někteří „filosofové“ Flegrovy názory ještě vylepšují. Třeba St. Komárek, který k „Zamrzlé evoluci“ napsal nadšenou předmluvu, přidává k Flegrově „revoluční teorii“ další zajímavý doplněk. Dnešní evolucionisté, rozuměj na rozdíl od Flegra, jsou podle Komárka přesvědčeni, „že hmota je cosi nadaného jen inercií, pasivní výplň prázdnoty, něco bez vlastní aktivity, neřku-li něčeho, jako je „intence“, či snad dokonce „záměr“ či „vůle“. Zřejmě je to jinak. Nemluví-li Komárek o Bohu, pak se postmoderně vrací k dávno překonané Drieschově entelechii a konečnému bodu Omega Teilharda de Chardin."
- Viz např.:
- "Podtitul knihy je tedy chybný, nepravdivý, matoucí a křivdící Darwinovi, nehledě k tomu, že ani není příliš slušný, že, pane Flegr."
- "A uvědomil si Flegr, že svou knihou, resp. svými výroky nahrává kreacionistům? Svědci Jehovovi, „vědečtí kreacionisté“ i dnešní množící se zastánci „inteligentního designu“ mezi našimi postmoderními intelektuály, ti všichni se na jeho knihu budou s velkým halasem odvolávat. „Vidíte, i velký evoluční biolog....atd."
- "Kniha je skvělou učebnicí evoluční biologie (darwinismu). Každému ji doporučuji, protože jde o téma opomíjené a protože se biologie dnes dostává stále více a rychleji do středu nejen vědeckého výzkumu, ale i do praktických aplikací. Čtěte ji však jen do poloviny, pak ji s klidem odložte, dáte byste nerozuměli nebo byste byli zmateni."
- "Kniha by si zasloužila i další ocenění, nejen cenu „Magnesia litera“. (...) To za první část knihy. Za tu druhou bych Vám, pane Flegr, rád předal stejně významnou cenu, a to za bludné myšlení a matení veřejnosti, cenu „Bludný Balvan“."
Reference
- FLEGR, Jaroslav. Evoluční tání: aneb O původu rodů. 1. vyd. Praha: Academia, 2015. 404 s. ISBN 978-80-200-2481-7.
- FLEGR, Jaroslav. Frozen Evolution: Or, that's not the way it is, Mr. Darwin. A Farewell to Selfish Gene.. 1. vyd. [s.l.]: Univerzita Karlova, Prirodovedecka Fakulta, 2008. 226 s.
- FLEGR, Jaroslav. Zamrzlá evoluce: aneb Je to jinak, pane Darwin. 1. vyd. Praha: Academia, 2006. 326 s. ISBN 978-80-200-1526-6.
- FLEGR, Jaroslav. On the “origin” of natural selection by means of speciation. Rivista Di Biologia-Biology Forum. Roč. 1998, čís. 91(2), s. 291–304. Dostupné online.
- FLEGR, Jaroslav. Elastic, not plastic species: frozen plasticity theory and the origin of adaptive evolution in sexually reproducing organisms. Biology Direct. 2010, čís. 5. Dostupné online.
- Archivovaná kopie. www.magnesia-litera.cz [online]. [cit. 2010-01-25]. Dostupné v archivu pořízeném dne 2009-05-07.
- FLEGR, Jaroslav. Was Lysenko (partly) right? Michurinist biology in the view of modern plant physiology and genetics. Rivista Di Biologia-Biology Forum. 2002, čís. 95(2), s. 259–271. Dostupné online.
- FLEGR, Jaroslav. Microevolutionary, macroevolutionary, ecological and taxonomical implications of punctuational theories of adaptive evolution. Biology Direct. 2013, čís. 8. Dostupné online.
- FLEGR, Jaroslav, KULICH, Tomáš. Positive effects of multiple gene control on the spread of altruism by group selection. Jornal of theoretical biology. 2011, čís. 284(1), s. 1–6.
- TOMAN, Jan. Pohlavní rozmnožování optikou evoluce: Vznik, vývoj a paradoxy největší evoluční záhady. 1. vyd. Praha: Academia, 2015. 416 s. ISBN 978-80-200-2306-3.
- TOMAN, Jan. Diplomová práce: Role ekologických faktorů při udržování sexuality [online]. Praha: 2013 [cit. 2015-12-05]. Dostupné online.
- ELDREDGE, Niles, GOULD, Stephen (1972): "Punctuated equilibria: an alternative to phyletic gradualism" In T.J.M. Schopf, ed., Models in Paleobiology. San Francisco: Freeman Cooper. pp. 82-115. Reprinted in N. Eldredge Time frames. Princeton: Princeton Univ. Press, 1985, pp. 193-223 Archivováno 22. 4. 2009 na Wayback Machine.
- TOMAN, Jan. Teorie zamrzlé plasticity a zamrzlé evoluce: Paradoxy neadaptability. Sborník abstraktů a příspěvků z 16. výjezdního interdisciplinárního semináře v Nečtinech. [online]. Plzeň: Meziborové aktivity Výzkumného centra Nové technologie Západočeské univerzity v Plzni, 19. - 22. 1. 2015 [cit. 2015-12-06]. Dostupné online.
- MIKULÁŠ, Radek. The principle of "frozen evolution" and its manifestation in the fossil record: The Brachiopod genus Aegiromena Havlíček, 1961 (upper Ordovican, Czech Republic) [online]. Plzeň: Palaeontological workshop held in honour of Doc. RNDr. Jaroslav Kraft, CSc. (9. 4. 1940–10. 1. 2007), 2008 [cit. 2015-12-05]. Dostupné online.
- GOULD, Stephen. Wonderful Life: The Burgess Shale and the Nature of History. 1. vyd. New York, London: W. W. Norton & Company, 1989. 346 s. Dostupné online. ISBN 0-393-02705-8.
- DAVISON, John. Evolution as a Self-Limiting Process. Rivista Di Biologia-Biology Forum. 1998, čís. 91(2), s. 199–220.
- HENG, Henry. The genome-centric concept: resynthesis of evolutionary theory. Bioessays. 2009, čís. 31(5), s. 512–525.
- GORELICK, Root, HENG, Henry. Sex reduces genetic variation: a multidisciplinary review. Evolution. 2011, čís. 65(4), s. 1088–1098.
- WILKINS, Adam, HOLLIDAY, Robert. The evolution of meiosis from mitosis. Genetics. 2009, čís. 181(1), s. 3–12.
- ŠPINKA, Marek. Potřebuje se evoluční biologie vyvíjet?: Flegrova Zamrzlá evoluce je zajímavá, důležitá a dobře napsaná kniha. Vesmír. 2007/5, roč. 86, s. 325–326. Dostupné online.
- MIKULÁŠ, Radek. Není všechno pták, co zdechlo: Ad Tomáš Grim a Vladimír Remeš: Potřebujeme skutečně nový model evoluce?. Vesmír. 2007/7, roč. 86, s. 462–463. Dostupné online.
- OSTDIEK, Gerald. Cast in Plastic: Semiotic Plasticity and the Pragmatic Reading of Darwin. Biosemiotics. 2011, čís. 4(1), s. 69–82.
- SHCHERBAKOV, Victor. Stasis is an Inevitable Consequence of Every Successful Evolution. Biosemiotics. 2012, čís. 5(2), s. 227–245.
- MARKOŠ, Anton. Biosphere as semiosphere: Variations on Lotman. Sign System Studies. 2014, čís. 42(4), s. 487–498. Dostupné online.
- HALL, Brian. A review of Frozen Evolution: Or That’s Not the Way It is, Mr. Darwin, edited by Jaroslav Flegr. Evolution & Development. 2009, čís. 11(1), s. 126–129. Dostupné online.
- ZIMA, Jan. Book Review. Flegr J., 2008: Frozen evolution. Or, that’s not the way it is, Mr. Darwin.. Folia Zoologica. 2009, čís. 58(2), s. 240. Dostupné online.
- REEVE, Catherine. Book reviews. Frozen Evolution: Or that’s not the way it is, Mr. Darwin. Human Ethology Bulletin. 2010, čís. 25(4), s. 5–8. Dostupné online.
- Záznam diskuse v Městské knihovně v Praze z roku 2008 mezi profesory Flegrem, Zrzavým a docentem Storchem. http://www.rozhlas.cz/leonardo/priroda/_zprava/415794
- KRATOCHVÍL, Lukáš: Plastická evoluce - je to jinak, pane Baldwin - Úloha fenotypové plasticity v makroevoluci. Přednáška v cyklu Biologické čtvrtky ve Viničné 22. 10. 2009. http://bio.natur.cuni.cz/~leb/ctvrtky.php Archivováno 11. 12. 2015 na Wayback Machine
- ČERNÝ, Robert: Od moderní k postmoderní evoluční biologii - Evo-devo, embrya, genové regulační sítě a makroevoluce. Přednáška v cyklu Biologické čtvrtky ve Viničné 12. 11. 2009. http://bio.natur.cuni.cz/~leb/ctvrtky.php Archivováno 11. 12. 2015 na Wayback Machine
- GRIM, Tomáš, REMEŠ, Vladimír. Potřebujeme skutečně nový model evoluce?. Vesmír. 2007/5, roč. 86, s. 320–326. Dostupné online.
- FLEGR, Jaroslav. Zamrzlá evoluční biologie?. Vesmír. 2007/5, roč. 86, s. 322–325. Dostupné online.
- FLEGR, Jaroslav, GRIM, Tomáš: Zamrzla evoluce? Je to jinak pane Flegr? - Hádání o dědivosti, fosíliích a účinnosti selekce. Přednáška v cyklu Biologické čtvrtky ve Viničné 10. 10. 2007. http://bio.natur.cuni.cz/~leb/ctvrtky.php Archivováno 11. 12. 2015 na Wayback Machine
- HEŘT, Jiří. FLEGR versus Darwin [online]. SISYFOS, 2007/5 [cit. 2015-12-05]. Dostupné online.
- GRAUR, Dan. New biological books. Frozen Evolution: Or, That’s not the Way It Is, Mr. Darwin: A Farewell to Selfish Gene.. The quarterly review of biology. 2010, čís. 85, s. 501–502. Dostupné online.
- TOMAN, Jan. Evoluce už není, co bývala: Jaroslav Flegr: Evoluční tání aneb O původu rodů. Vesmír. 2015/6, roč. 94, s. 366–369. Dostupné online.
- TUREČEK, Petr. Evoluční tání v Praze. Respekt. 27. 9. 2015. [http://www.respekt.cz/tydenik/2015/40/evolucni-tani-v-praze https://www.natur.cuni.cz/fakulta/pro-media/aktuality/evolucni-tani-v-praze-respekt-29.9.2015-rubrika-civilizace-strana-61 Dostupné online].
- FLEGR, Jaroslav: Postneodarwinistic theories of evolution. Rethinking evolutionary theory from the perspective of cultural studies. Center for Literary and Cultural Research, Berlin 30. 10. - 2. 11. 2008.
- FLEGR, Jaroslav: Why is evolution frozen and what does it mean for life sciences - Evolutionary and ecological implications of punctuational theories of adaptive evolution. Rethinking evolutionary theory from the perspective of cultural studies. 3rd ISHE Summer Institute, Prague 5. - 9. 8. 2011.
- FLEGR, Jaroslav: From selfish gene to frozen evolution. 12th VBC PhD Symposium, Vienna 6. - 7. 11. 2014.
- TOMAN, Jan. Když je neadaptabilita adaptivní: aneb Co nám mohou povědět dnešní modely evoluce o vzniku sexuality. Vesmír. 2010/7, roč. 89, s. 414–415. Dostupné online.
- TOMAN, Jan. Evoluce jako starobinec: aneb Stárnou evoluční linie, nebo nestárnou?. Vesmír. 2015/3, roč. 89, s. 149–151. Dostupné online.
- TOMAN, Jan: Poslední zhasne - Zamrzlá plasticita a konec evoluce. Přednáška v cyklu Biologické čtvrtky ve Viničné 12. 3. 2015. http://bio.natur.cuni.cz/~leb/ctvrtky.php Archivováno 11. 12. 2015 na Wayback Machine Nahrávky přednášky a diskuse jsou dostupné na youtube.
- TOMAN, Jan: Kam kráčí evoluční biologie? Přednáška v cyklu čtvrtečních seminářů v CTS 28. 5. 2015. http://www.cts.cuni.cz/ctvrtecni-seminare/Jan-Toman-Kam-kraci-evolucni-biologie.html
- ŠPINKA, Marek. Nezastydlá evoluční teorie. Respekt. 29. 4. 2007, čís. 2007/18. Dostupné online.
- FLEGR, Jaroslav: Příroda není zase tak krutá. Rozhovor v časopisu Respekt 2007/19, 6. 5. 2007. http://www.respekt.cz/tydenik/2007/19/priroda-neni-zase-tak-kruta
- Rozhovor v MF DNES 5. 5. 2007
- Rozhovor v Lidových novinách 3. 2. 2007
- MATYÁŠ, Josef, FLEGR, Jaroslav: Zájem o nové směry je cesta do pekel, říká evoluční biolog. Rozhovor v Lidových novinách 22. 6. 2010. http://relax.lidovky.cz/zajem-o-nove-smery-je-cesta-do-pekel-rika-evolucni-biolog-pkc-/veda.aspx?c=A100621_164005_ln_veda_ev
- MRÁZEK, Daniel, FLEGR, Jaroslav. Zamrzlá plasticita aneb Evoluční teorie po revizi. Rozhovor v ČRo Leonardo, vysíláno 28. 2. 2013. http://www.rozhlas.cz/priroda/expedice_vyzkum/_zprava/zamrzla-plasticita-aneb-evolucni-teorie-po-revizi--1029222
- FLEGR, Jaroslav: Dědivost není! - O vzniku přírodního výběru prostřednictvím speciace. Přednáška v cyklu Biologické čtvrtky ve Viničné 16. 10. 1997. http://bio.natur.cuni.cz/~leb/ctvrtky.php Archivováno 11. 12. 2015 na Wayback Machine
- ZRZAVÝ, Jan: Pravděpodobnost a fylogeneze - Anti-Flegr podruhé. Přednáška v cyklu Biologické čtvrtky ve Viničné 2. 4. 1998. http://bio.natur.cuni.cz/~leb/ctvrtky.php Archivováno 11. 12. 2015 na Wayback Machine
- FLEGR, Jaroslav: Evoluce skončila, zapomeňte - Kam kráčí teorie zamrzlé plasticity. Přednáška v cyklu Biologické čtvrtky ve Viničné 21. 4. 2011. http://bio.natur.cuni.cz/~leb/ctvrtky.php Archivováno 11. 12. 2015 na Wayback Machine
- Záznam přednášky Jaroslava Flegra na AFO 2015 o teorii sobeckého genu a teorii zamrzlé evoluce + diskuse s Danielem Fryntou. Nahrávka je dostupná na youtube.
Související články
Externí odkazy
- Záznam diskuse v Městské knihovně v Praze z roku 2008 mezi profesory Flegrem, Zrzavým a docentem Storchem.
- Záznam diskuse v rámci Biologických čtvrtků mezi profesorem Flegrem a Tomášem Grimem