Vířníci
Vířníci (Rotifera, Rotatoria) je parafyletická skupina převážně sladkovodních, mikroskopických živočichů s tělem rozčleněným na hlavu, trup a nohu a s nápadným obrveným vířivým aparátem v přední části. Někdy jsou mezi ně zahrnováni vrtejši, kteří jsou z fylogenetického hlediska podskupina vířníků extrémně pozměněná parazitismem. V tomto širším pojetí jsou vířníci monofyletickou skupinou (kmenem), která je charakterizovaná především syncytiální pokožkou obsahující proteinovou ochrannou laminu („vnitřní kutikulu“) a také neobvykle stavěnými spermiemi s předním (tažným) bičíkem. V užším, tradičnějším pojetí, tedy bez vrtejšů, jsou vířníci kromě členěného těla a vířivého aparátu typičtí mastaxem, osvaleným jícnem vybaveným chtinózními lištami, které slouží jako čelisti při drcení potravy. Typicky se živí řasami, prvoky, bakteriemi, organickým detritem apod., potravu si přihánějí s proudem vody za pomoci vířivého aparátu. Někteří se chovají jako predátoři drobných živočichů nebo jako parazité.
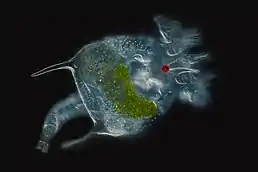 Krunýřenka Brachionus quadridentatus | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Nadoddělení | trojlistí (Triblastica) |
| Oddělení | prvoústí (Protostomia) |
| Kmen | vířníci (Rotifera) |
| třídy | |
| Některá data mohou pocházet z datové položky. | |
Vířníků je známo přes 2000 druhů (přičemž v Česku bylo zaznamenáno necelých 600 druhů). Vyskytují se po celém světě, obývají hlavně sladké, ale i slané vody, častí jsou i ve vodě zachycené v mechových polštářích či v půdě. Jsou schopni vytvářet odolná stádia, ať už jsou to vajíčka (u třídy točivky) nebo vyschlí dospělci ve stavu tzv. anabiózy (třída pijavenky). Vířníci mají silný sklon k partenogenezi. U točivek se partenogenetické generace střídají s generací pohlavní, kdy se z haploidních neoplozených vajec líhnou miniaturní samci. Pijavenky jsou proslulé svou asexualitou – samci se u nich vůbec nevyskytují, z vajec se líhnou vždy jen další partenogenetické samice, klony svých matek. Třetí třída vířníků, žábrovci, je ale striktně sexuální s plně vyvinutými samci i samicemi. Totéž platí i pro vrtejše.
.jpg.webp)
Charakteristika
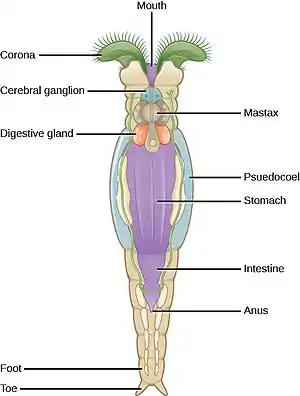
Vířníci v tradičním pojetí zahrnují drobné vodní organismy nápadné tělem rozděleným na tři oddíly (hlava, trup, noha), vířivým aparátem (koronou) tvořeným obrvenými políčky či pruhy v přední části hlavy a svalnatým žvýkacím jícnem (mastax).[1][2] Silně modifikovanou skupinou vířníků jsou však i vrtejši,[3][4] dříve, a někdy i v moderní literatuře,[5][6] řazení do samostatného kmene. Vrtejši se charakteristice svých volně žijících příbuzných značně vymykají. V následujícím textu jsou představeni volně žijící „klasičtí“ vířníci bez vrtejšů.
Stavba těla, vířivý aparát, pohyb
_Figure_1.jpg.webp)
Typičtí vířníci jsou mikroskopičtí tvorové dosahující nejčastěji velikosti jedné až několika desetin milimetru, výjimečně dorůstají až 3 mm (rod Seison). Samci točivek jsou mnohdy naopak menší než 0,1 mm. Vzhledem k drobné velikosti není u vířníků vyvinuta cévní ani dýchací soustava. Název kmene je odvozen od víření bičíků na přídi těla. Protože bičíky se nepohybují synchronně, ale každý bičík je trochu opožděn vůči sousednímu bičíku (vzniká tak tzv. metachronní vlna), budí vířníci s širokými kruhovými obrvenými poli po stranách hlavy dojem, že se vířivý aparát otáčí, rotuje – odsud název kmene Rotifera odvozený z řečtiny (doslova „nosiči koleček“).[5]
U některých vířníků tvoří vířivý aparát dva dobře definované pruhy brv, přední trochus (prototroch) a zadní cingulum (metatroch), mezi nimiž jsou ústa obklopená obrveným polem s kratšími bičíky. Toto uspořádání je některými autory považováno za homologické uspořádání bičíků trochoforové larvy měkkýšů, kroužkovců a dalších. Vířníci jsou pak interpretováni jako neoteničtí živočichové – v podstatě trochofory schopné množení.[3] Ačkoli jde o atraktivní hypotézu, fylogeneze vířníků (jejich příbuznost s kmeny bez trochofory) ani jejich embryogeneze ji nepodporují. Spíše se u trochofor a vířníků jedná o konvergentní dosažení morfologie optimalizované pro pohyb a filtraci potravy z vody.[4] U mnoha druhů vířníků je navíc vířivý aparát oproti výše popsanému stavu silně modifikovaný – některý z pruhů bičíků (někdy oba) např. může chybět,[3] variant uspořádání vířivého aparátu je popsána řada.[2] Vířivý aparát slouží jednak k plavání a jednak k přihánění potravy. Vířníci se totiž mohou za pomoci lepových žláz ústících na konci prstíků jejich nohy dočasně přichytit (přilepit) k podkladu a potravu filtrovat z proudu vody poháněného činností brv. Některé druhy jsou přisedlé trvale (jejich nedospělá stádia jsou ale volně plovoucí) a mnohdy si pak vytvářejí trubicovité ochranné schránky zpevněné někdy cizorodými částečkami typu zrnek písku nebo i fekálními peletami. Existují však i čistě planktonní druhy, které na substráty nepřisedají a nohu včetně lepových žláz mají zakrnělou.[5]
Vířníci jsou vybaveni okružními i podélnými svaly – ty podélné slouží zejména jako zatahovače hlavy (resp. korony) a nohy, okružní způsobují tlakem na tekutinu pseudocoelu protažení těla. Tyto pohyby (zkracování a natahování těla) ve spolupráci s lepovými žlázami na konci těla a dočasným uchycením k substrátu také vysunutým rostrem na předním konci těla umožňují píďalkovitý pohyb typický zejména pro zástupce třídy pijavenky. Jejich noha i tělo jsou rozděleny na zužující se pseudosegmenty (nepravé články vymezené záhyby na epidermis, ale nepromítající se do vnitřní stavby orgánů), které jsou schopné se do sebe teleskopicky zasouvat a tím přispívat ke změnám délky těla při píďalkovitém pohybu. Někteří vířníci mohou být ve volné vodě schopni kromě plynulého, brvami zajišťovaného plavání také rychlého trhavého pohybu díky prudkým, svalům zajišťovaným pohybům různých výrůstků.[5] Svalovina vířníků může být jak hladká (hlavně u pijavenek), tak příčně pruhovaná (spíše u točivek), případně může být pruhovaná i šikmo.[4]
Epidermis, lorika
Pokožka (epidermis) vířníků má neobvyklou stavbu: není tvořena jednotlivými buňkami, ale soubuním (syncytiem), v němž se vytváří intracelulární vrstva označovaná jako lamina nebo vnitřní kutikula (kromě toho je na povrchu těla přítomna i tenoučká extracelulární glykoproteinová kutikula).[1][5] Vzhledem ke své lokalizaci není vnitřní kutikula svlékána. Je tvořena proteiny keratinového typu.[3] Někdy je lamina zesílena a tvoří pevnou schránku zvanou lorika chránící trupovou část těla. Lorika může vybíhat v trny, ostny či jiné útvary. Ty jsou důležité pro určování, zároveň však mohou být v rámci druhu variabilní např. v závislosti na roční době, množství predátorů aj.[5]
Syncytiální epidermis s intracelulární kutikulou je rys, který volně žijící vířníci sdílejí s vrtejši.[5] Obrvené části hlavy vířníků jsou tvořeny velkými několikajadernými mnohobičíkatými buňkami.[3]
Trávení, vylučování
Tělo vířníků je pseudocoelomátní, kolem trávicí trubice je tedy tekutinou vyplněná štěrbinovitá dutina (pseudocoel). V silně osvaleném žvýkacím hltanu (mastaxu) se nacházejí chitinové ozubené lišty (trophi),[1] které pomáhají zpracovávat pohlcenou potravu. Někdy lišty vyčnívají z úst a pomáhají lovit nebo vysávat kořist.[5] Bývají uspořádány symetricky, existují ale i druhy s asymetricky utvářeným mastaxem.[2] Lišty mastaxu jsou považovány za homologické čelistem čelistovek a oknozubek a chitinovým ostnům kolem úst ploutvenek – jde tedy o společný znak zástupců skupiny Gnathifera.[7] Vlastní trávení se odehrává hlavně v žláznatém žaludku za mastaxem, do něhož vyúsťuje párová žaludeční žláza někdy označovaná i jako hepatopankreas. Produkuje trávicí enzymy.[1] Střevní epitel je obrvený – bičíky pomáhají posunovat tráveninu vzad. Střevo je u většiny vířníků průchozí, pak vyúsťuje společně s vylučovací a rozmnožovací soustavou prostřednictvím kloaky, která se navenek otevírá v dorzální oblasti na rozhraní trupu a nohy. U některých pijavenek je trávicí soustava za mastaxem redukovaná na syncytiální masu v oblasti žaludku. Trávení zde probíhá velkou měrou intracelulárně a trávicí soustava v tomto případě končí slepě.
Vylučovací soustava je tvořena párem protonefridií, z nichž každá může být tvořena jednou až 50 plaménkovými buňkami. Odvodné kanálky protonefridií vedou do močového měchýře, který při kontrakci, která nastává až 6× za minutu, vypudí tekutinu do kloaky a ven z těla. Především u sladkovodních vířníků je role protonefridií hlavně osmoregulační (vylučují přebytečnou vodu, která se do těla neustále dostává na základě osmózy).[5]
Nervová soustava

Nervová soustava je tvořena několika ganglii, přičemž největší je hlavové ganglion (mozek) ležící dorzálně od mastaxu.[4][5] Přímo do něj může být integrován miskovitý fotoreceptor, další očka mohou být přítomna v oblasti vířivého aparátu nebo na rostru. Další výrazná ganglia jsou nožní ganglion nebo ganglion mastaxu.[4] Na těle mohou být přítomna tykadla, papily či svazky brv s chemo- a mechanoreceptorickou funkcí.[4][5]
Rozmnožování, asexualita, anabióza, eutelie
Vířníci jsou gonochoristé, jejichž rozmnožování probíhá rozmanitými způsoby od standardního pohlavního rozmnožování (třída žábrovci) přes střídání pohlavního množení s partenogenetickým množením (třída točivky) až po čistě partenogenetické množení pomocí neoplozených vajec u pijavenek – samci se u nich vůbec nevyskytují, potomci jsou klony svých matek. I u točivek má asexuální množení z neoplozených, mitózou vzniklých, tzv. amiktických vajíček během roku výraznou převahu. Doba zdvojení populace vířníků v amiktické fázi může být pouhých 15 hodin, jedna samice vyprodukuje za život několik až několik desítek (cca 5–40) vajíček. Při změně podmínek začnou některé samice točivek produkovat miktická, meiózou vzniklá, a tedy haploidní vajíčka. Zůstanou-li neoplozena, líhnou se z nich haploidní, krátkověcí, trpasličí, čile se pohybující samečci se zakrnělou trávicí soustavou. Kopulace bývá tzv. traumatická, tzn., že samec svým penisem injikuje spermie do těla samice přes tělní stěnu přímo do pseudocoelu. Pokud se samcům svými značně atypickými spermiemi s předním (tažným) bičíkem, jichž nesou omezený počet,[1] podaří oplodnit miktická vajíčka, začnou se tato vyvíjet v tzv. zimní vajíčka obalená silnější stěnou a schopná přestát nepříznivé podmínky. Mohou zůstat životaschopná minimálně kolem sta let.[8] Líhne se z nich další generace amiktických samic.[5] Vajíčka jsou v těle samice vytvářena v párovém (pijavenky) či nepárovém (točivky) germovitelláriu (pohlavně-žloutkové žláze), kde syncytiální vitellárium produkuje žloutkové buňky, které vajíčko zásobují živinami.[1]
Asexualita pijavenek je patrně neobyčejně stará – desítky milionů let. Takto staré asexuální linie jsou mezi živočichy velice vzácné, což je interpretováno tak, že druhy a skupiny, které ztratily schopnost pohlavního množení, a tedy i schopnost produkce variabilního potomstva, brzy vymírají. Za nejvýznamnější faktor způsobující vymírání klonálních populací jsou považovány patogenní organismy, které se mohou populacím geneticky uniformních organismů snadno přizpůsobit. Pijavenky jsou však schopné dlouhodobě přečkávat nepříznivé podmínky ve vyschlém, neaktivním stavu – ve stavu anabiózy (kryptobiózy). Zároveň se mohou v tomto stavu šířit větrem na velmi vzdálené lokality. Zdá se, že díky této schopnosti jsou schopny zbavit se některých svých patogenů (plísní ze skupiny řasovky), kteří tak dlouhé vyschnutí nesnesou. Dormantní vířníci se navíc snadno šíří do nezamořených oblastí. Schopnost anabiózy by tak u pijavenek souvisela s možností dlouhodobě se obejít bez sexu.[9] Během anabiózy jsou v podstatě zastaveny jejich životní pochody do doby, než se opět obnoví příznivé podmínky. V tomto stavu jsou kromě sucha schopny přečkat i další extrémní podmínky (nízké a vysoké teploty, ale i zvýšenou radiaci – v souvislosti se schopností dlouhodobé dormance mají dobře vyvinuté mechanismy oprav DNA).[2] „Oživit“ pijavenky v kryptobióze se podařilo i po 20 letech vyschnutí[5]. Zamrzlé v sibiřském permafrostu dokonce zůstaly životaschopné 24 000 let.[10] Schopnost dobře reparovat poškozenou DNA je dávána do souvislosti také s faktem, že genomy vířníků obsahují neobvykle vysoký podíl (asi do 10 %) genů jiných organismů získaných horizontálním genovým transferem. Jedna z hypotéz tuto skutečnost vysvětluje tak, že při reparaci rozsáhlých poškození DNA, jako jsou např. dvouřetězcové zlomy, je do DNA vířníků jaksi „omylem“ inkorporována i cizorodá DNA. Platnost této hypotézy ale byla zpochybněna a otázka masivního transferu genů do genomu vířníků zůstává otevřená.[4]
Vířníci vykazují tzv. eutelii, konstantní počet buněk (resp. jader v případě syncytiálních tkání) v těle i v jednotlivých orgánech u všech jedinců téhož druhu (a pohlaví).[5] Jader bývá v těle vířníků kolem tisíce.[4][11] U některých druhů se však lze setkat i s variabilním počtem jader, alespoň v některých tkáních.[4] Tak či tak, počet buněk / jader se zvyšuje jen během rané ontogeneze, po vylíhnutí již nepřibývají[3][4] a organismus roste jen díky zvětšování buněk.[1] Eutelie je spojena s neschopností regenerace.[3][1] Vývoj vířníků je přímý. Dospělci se dožívají se jen několika týdnů (pokud neupadnou na delší dobu do stavu anabiózy).[5]
Vrtejši
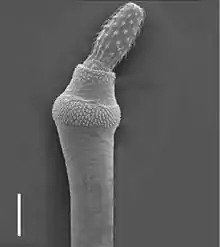
Zvláštní podskupinou vířníků, zde jen stručně zmíněnou, jsou vrtejši považovaní často za samostatný kmen.[3][4] Je to čistě endoparazitická skupina, jejíž zástupci parazitují ve střevě různých obratlovců, hlavně ryb. Jejich mezihostitelem bývá členovec. Měří několik mm až více než půl metru.[1][4] Podobně jako ostatní vířníci mají syncytiální epidermis s intracelulární kutikulou (laminou), spermie s tažným bičíkem[1] a mají konstantní počet buněk (jader), jsou tedy euteličtí. Jinak se od vířníků liší absencí trávicí soustavy, jednodušší stavbou těla (chybí jim vířivý aparát i noha) a dalšími znaky. Nápadným útvarem v přední části jejich těla je otrněný zatažitelný rypák, který slouží k ukotvení ve střevní sliznici. Vrtejši jsou gonochoristé, vyvíjejí se přes několik larválních stádií.[5]
_(19558263630).jpg.webp)
Diverzita, systematika

První vířníci (pijavenky) byli pozorováni A. van Leeuwenhoekem, který je zmiňuje v dopise z roku 1687.[2] Původně byli vzhledem ke své velikosti a obrvení považováni za nálevníky, teprve později byla rozpoznána jejich příslušnost mezi mnohobuněčné živočichy.[11] Vířníci v tradičním pojetí, tedy bez vrtejšů, se dělí na tři třídy: točivky, pijavenky a žábrovce.
Třída točivky (Monogononta) je nejpočetnější (přes 1500 druhů) a z hlediska ekologie a životních strategií nejpestřejší třídou vířníků. Jsou známy čistě planktonní i trvale přisedlé zástupce, někdy žijící v rourkách a někdy vytvářející celé kolonie. Kromě druhů, které využívají ke shánění potravy brvy vířivého aparátu, jsou známí i predátoři, kteří mají kolem úst dlouhá ramena s dlouhými tuhými bičíky fungujícími jako past na drobné organismy. U některých točivek je kutikula zesílena v loriku. Pro točivky je typické haplo-diploidní určení pohlaví a střídání nepohlavní (amiktické) a pohlavní (miktické) fáze v životním cyklu,[3][5] čili tzv. heterogonie. Jejich samičky mají jen jediný vaječník (proto „Monogononta“).[1][3]
Do třídy pijavenky (Bdelloidea) patří výhradně volně žijící a pohybliví (nepřisedlí) zástupci, celkem přes 400 druhů. Zpravidla mají dobře vyvinutý, dvěma laloky tvořený vířivý aparát, jímž si přihánějí vodu s částečkami potravy. Jejich zvláštností je výše zmíněná absence sexuálního rozmnožování.[5] Mají párový vaječník.[3] Vzhledem k tomu, že často obývají vysýchavá nebo zamrzající prostředí (periodické tůňky, mech, vlhké půdy, polární jezírka apod.), jsou schopny upadat do stavu kryptobiózy (anabiózy).[1]
Nejmenší třída žábrovci (Seisonidea) s pouhými čtyřmi druhy řazenými do rodů Seison a Paraseison[12] zahrnuje velké (až 3 mm dlouhé) ektoparazity vyskytující se na mořských korýších rodu Nebalia. Vzhledem ke způsobu života mají silně redukovaný vířivý aparát, místo prstíků mají nohu zakončenou přísavným diskem. Jde o gonochoristy se striktně pohlavním rozmnožováním, samci jsou dobře vyvinuti.[5]
Od 80. let 20. století začalo být zřejmé, že volně žijící vířníci a parazitičtí vrtejši spolu sdílejí některé detailní morfologické znaky[5] jako je zejména stavba pokožky[13] nebo spermií[14] a vrtejši začali být považováni nejen za sesterskou skupinu vířníků, ale přímo za jejich parazitismem modifikovanou podskupinu.[15] Tento názor byl záhy podpořen celou řadou molekulárně-fylogenetických studií.[4] Vířníci jsou pak považováni buď za parafyletickou skupinu a monofyletická skupina zahrnující spolu s nimi i vířníky je označována jako Syndermata,[5][6] nebo jsou vrtejši do kmene vířníků zahrnuti jako podskupina – označení Syndermata je pak synonymem pro rozšířený kmen Rotifera.[3][4] Vzájemné příbuzenské vztahy mezi čtyřmi skupinami vířníků (točivky, pijavenky, žábrovci, vrtejši) zůstávají nejasné, různé analýzy vedou k odlišným závěrům. Bazální postavení mají snad v rámci kmene točivky.[4][5] Každopádně prakticky nikdy není podporována sesterská pozice vrtejšů vůči (ostatním) vířníkům, vrtejši jsou vždy rekonstruováni jako vnitřní skupina vířníků.[2][6][16]
Fosilie vířníků jsou vzácné a jsou relativně mladého data, nejstarší nálezy jsou eocenní.[4] Většinou jde o fosilní pozůstatky pijavenek v jantaru. Subfosilní nálezy v holocenních rašelinách apod. jsou hojnější.[2]
Výskyt
Většina z více než 2000 popsaných druhů vířníků žije v různorodých sladkovodních biotopech od jezer až po vodní mikrobiotopy ve vlhkých mechových polštářích a v půdě. Mohou být planktonní nebo preferovat různé ponořené povrchy, někdy jsou intersticiální (vyskytují se mezi zrnky písku).[5] Menší část druhů (kolem 15 %) se vyskytuje ve slaných a brakických vodách, ať už vnitrozemských nebo mořských. Vyskytují se na všech kontinentech včetně Antarktidy.[2] Točivky obývají spíše rozsáhlejší vodní biotopy, směrem k pólům jejich diverzita klesá. Pijavenky jsou hojné i ve vlhkých mikroprostředích a v teplých oblastech jsou vzácnější.[2] Mezi vířníky jsou ale zastoupeni i ektokomenzálové až ektoparazité nejrůznějších vodních bezobratlých, zejména členovců a kroužkovců, jakož i endoparazité opaskovců a plžů.[5] Zástupci rodu Pourriotia parazitují ve stélkách řas rodu Vaucheria a vyvolávají na nich tvorbu jakýchsi miniaturních hálek. Další vířníci (např. Dicranophorus, Asciaporrecta) mohou parazitovat ve schránkách krytenek (rozlitka, štítenka). Na žábrách ryb žije jako komenzál nebo parazit Encentrum kozminskii.[2] Pokud jsou mezi vířníky započítáni i vrtejši, přispějí k jejich diverzitě více než tisícovkou druhů parazitujících ve střevě obratlovců.[4]
Na území Česka bylo k r. 2019 nalezeno 574 druhů vířníků, předpokládá se však (vzhledem k nálezům z okolních zemí), že se zde může vyskytovat dalších asi 400 druhů, které dosud nebyly zaznamenány.[17]
Význam
Vířníci jsou důležitým článkem potravních řetězců ve vodě: živí se jednobuněčnými organismy (bakterie, řasy, prvoci), drobnými živočichy (např. jinými druhy vířníků) či organickým detritem[1][5] a sami se stávají kořistí větších živočichů, jako jsou drobní korýši, kteří jsou zase složkou potravy mnoha ryb.[18] Rybí potěr se může vířníky živit přímo a v akvakultuře jsou některé druhy vířníků využívány jako první potrava ryb nebo krevet. V mořské akvaristice mají tento význam hlavně brakické a mořské druhy rodu krunýřenka (Brachionus), např. B. plicatilis.[19][20] Dále mohou vířníci sloužit i jako indikátory znečištění vod nebo jako modelové organismy při výzkumu stárnutí, anabiózy i evoluce sexuality.[4][5]
Odkazy
Reference
- SMRŽ, Jaroslav. Základy biologie, ekologie a systému bezobratlých živočichů. Praha: Karolinum, 2013. ISBN 978-80-246-2258-3. S. 47–51.
- FONTANETO, Diego; DE SMET, Willem H. Handbook of Zoology. Příprava vydání Andreas Schmidt-Rhaesa. Svazek Volume 3: Gastrotricha and Gnathifera. [s.l.]: DE GRUYTER, 2015. Dostupné online. ISBN 978-3-11-027381-6. DOI 10.1515/9783110274271.217. Kapitola 4. Rotifera, s. 217–300.
- NIELSEN, Claus. Animal evolution : interrelationships of the living phyla. 3rd ed. vyd. Oxford: [s.n.], 2012. 1 online resource (x, 402 pages) s. Dostupné online. ISBN 978-0-19-162530-5, ISBN 0-19-162530-2. OCLC 771927420 Kapitola 34: Phylum Rotifera, s. 185–194.
- GIRIBET, Gonzalo; EDGECOMBE, Gregory. The invertebrate tree of life. Princeton, New Jersey: [s.n.], 2020. 1 online resource (xvi, 589 pages) s. Dostupné online. ISBN 978-0-691-19706-7, ISBN 0-691-19706-7. OCLC 1129197548 Kapitola 39: Rotifera, s. 293–309.
- PECHENIK, Jan A. Biology of the invertebrates. Seventh edition. vyd. New York, NY: [s.n.], 2015. xv, 606 pages s. Dostupné online. ISBN 978-0-07-352418-4, ISBN 0-07-352418-2. OCLC 863127655 Kapitola 10: The Gnathifera: Rotifers, Acanthocephalans, and Two Smaller Groups, s. 183–203.
- SCHIERWATER, Bernd; DESALLE, Rob. Invertebrate zoology : a tree of life approach. First edition. vyd. Boca Raton: [s.n.], 2021. xvi, 627 pages s. Dostupné online. ISBN 978-1-4822-3581-4, ISBN 1-4822-3581-1. OCLC 1201692976
- MARLÉTAZ, Ferdinand; PEIJNENBURG, Katja T.C.A.; GOTO, Taichiro. A New Spiralian Phylogeny Places the Enigmatic Arrow Worms among Gnathiferans. Current Biology. 2019, roč. 29, čís. 2, s. 312–318.e3. Dostupné online [cit. 2021-12-19]. DOI 10.1016/j.cub.2018.11.042. (anglicky)
- PISCIA, Roberta; GUILIZZONI, Piero; FONTANETO, Diego. Dynamics of rotifer and cladoceran resting stages during copper pollution and recovery in a subalpine lake. Annales de Limnologie - International Journal of Limnology. 2012, roč. 48, čís. 2, s. 151–160. Dostupné online [cit. 2021-12-20]. ISSN 0003-4088. DOI 10.1051/limn/2012006.
- WILSON, Christopher G.; SHERMAN, Paul W. Anciently Asexual Bdelloid Rotifers Escape Lethal Fungal Parasites by Drying Up and Blowing Away. Science. 2010-01-29, roč. 327, čís. 5965, s. 574–576. Dostupné online [cit. 2021-12-20]. ISSN 0036-8075. DOI 10.1126/science.1179252. (anglicky)
- SHMAKOVA, Lyubov; MALAVIN, Stas; IAKOVENKO, Nataliia. A living bdelloid rotifer from 24,000-year-old Arctic permafrost. Current Biology. 2021-06, roč. 31, čís. 11, s. R712–R713. Dostupné online [cit. 2021-12-21]. DOI 10.1016/j.cub.2021.04.077. (anglicky)
- KOMÁREK, Julius. Zoologie bezobratlých. Praha: Přírodovědecké vydavatelství Č. akademie věd a umění, 1952. Kapitola Rotatoria (Rotifera), s. 239–247.
- LEASI, Francesca; ROUSE, Greg W.; SØRENSEN, Martin V. A new species of Paraseison (Rotifera: Seisonacea) from the coast of California, USA. Journal of the Marine Biological Association of the United Kingdom. 2012-08, roč. 92, čís. 5, s. 959–965. Dostupné online [cit. 2021-12-20]. ISSN 0025-3154. DOI 10.1017/S0025315411000129. (anglicky)
- CLÉMENT, Pierre. Phylogenetic relationships of rotifers, as derived from photoreceptor morphology and other ultrastructural analyses. Hydrobiologia. 1980-08, roč. 73, čís. 1–3, s. 93–117. Dostupné online [cit. 2021-12-20]. ISSN 0018-8158. DOI 10.1007/BF00019432. (anglicky)
- FERRAGUTI, M.; MELONE, G. Spermiogenesis in Seison nebaliae (Rotifera, Seisonidea): further evidence of a rotifer-acanthocephalan relationship. Tissue and Cell. 1999-08, roč. 31, čís. 4, s. 428–440. Dostupné online [cit. 2021-12-20]. DOI 10.1054/tice.1999.0012. (anglicky)
- LORENZEN, S. The Origins and Relationships of Lower Invertebrates. Příprava vydání Conway Morris et al.. Oxford: Oxford University Press, 1985. Kapitola Phylogenetic aspects of pseudocoelomate evolution.
- SIELAFF, Malte; SCHMIDT, Hanno; STRUCK, Torsten H. Phylogeny of Syndermata (syn. Rotifera): Mitochondrial gene order verifies epizoic Seisonidea as sister to endoparasitic Acanthocephala within monophyletic Hemirotifera. Molecular Phylogenetics and Evolution. 2016-03, roč. 96, s. 79–92. Dostupné online [cit. 2021-12-20]. DOI 10.1016/j.ympev.2015.11.017. (anglicky)
- DEVETTER, Miloslav, et al. Checklist of rotifers (Phylum: Rotifera) of the Czech Republic (Aktuální seznam vířníků (Kmen: Rotifera) České republiky). Příroda (sborník Západočeského muzea v Plzni). 2020. Dostupné online.
- WALLACE, R. L. Rotifers: Exquisite Metazoans. Integrative and Comparative Biology. 2002-07-01, roč. 42, čís. 3, s. 660–667. Dostupné online [cit. 2021-12-20]. ISSN 1540-7063. DOI 10.1093/icb/42.3.660. (anglicky)
- LUBZENS, E. Raising rotifers for use in aquaculture. Hydrobiologia. 1987-04, roč. 147, čís. 1, s. 245–255. Dostupné online [cit. 2021-12-20]. ISSN 0018-8158. DOI 10.1007/BF00025750. (anglicky)
- MOTYČKA, Vladimír; ROLLER, Zdeněk. Bezobratlí (1). 1. vyd. [s.l.]: Albatros, 2001. (Svět zvířat; sv. X). ISBN 80-00-00884-X. S. 40–42.
Česky
- BUCHAR, Jan, a kol. Klíč k určování bezobratlých. 1. vyd. [s.l.]: Scientia, 1995. ISBN 80-85827-81-6. Kapitola Kmen: vířníci - Rotatoria, s. 57–64.
- SMRŽ, Jaroslav. Základy biologie, ekologie a systému bezobratlých živočichů. Praha: Karolinum, 2013. ISBN 978-80-246-2258-3. S. 47–51.
- BARTOŠ, Emanuel. Klíč zvířeny ČSR. Příprava vydání S. Hrabě a kol.. Svazek 1. Praha: Nakladatelství ČSAV, 1954. Kapitola Vířníci – Rotatoria, s. 248–286.
Anglicky
- FONTANETO, Diego; DE SMET, Willem H. Handbook of Zoology. Příprava vydání Andreas Schmidt-Rhaesa. Svazek Volume 3: Gastrotricha and Gnathifera. [s.l.]: DE GRUYTER, 2015. Dostupné online. ISBN 978-3-11-027381-6. DOI 10.1515/9783110274271.217. Kapitola 4. Rotifera, s. 217–300.
- PECHENIK, Jan A. Biology of the invertebrates. Seventh edition. vyd. New York, NY: [s.n.], 2015. xv, 606 pages s. Dostupné online. ISBN 978-0-07-352418-4, ISBN 0-07-352418-2. OCLC 863127655 Kapitola 10: The Gnathifera: Rotifers, Acanthocephalans, and Two Smaller Groups, s. 183–203.
Externí odkazy
 Obrázky, zvuky či videa k tématu vířníci na Wikimedia Commons
Obrázky, zvuky či videa k tématu vířníci na Wikimedia Commons  Taxon Rotifera ve Wikidruzích
Taxon Rotifera ve Wikidruzích