TLR4
Toll-like receptor 4 je protein, u člověka kódovaný genem TLR4. TLR4 je transmembránový protein, patřící do rodiny toll-like receptorů, které patří mezi pattern-recognition receptory (PRR). Jeho aktivace vede ke spuštění intracelulární signální dráhy NF-κB, produkci pro-zánětlivých cytokinů a aktivaci odpovědi vrozené imunity.[1] Jeho nejznámějším ligandem je lipopolysacharid (LPS), složka přítomná na vnější membráně gramnegativních bakterií, ale i na některých grampozitivních bakteriích. Mezi další ligandy patří některé virové proteiny, polysacharidy a různé endogenní proteiny, jako například nízkodenzitní lipoprotein (LDL), beta-defensiny a proteiny teplotního šoku (HSP).[2]
TLR4 byl zároveň označen jako CD284 (diferenciační skupina 284).[3] Molekulární váha TLR4 je přibližně 95kDa.[4]
Funkce
TLR4 je patří do rodiny toll-like receptorů (TLR), která zastává zásadní roli při rozpoznání patogenů a aktivaci vrozené imunity. Rozeznávají s patogeny asociované molekulární vzory (PAMPs), které jsou exprimovány na infekčních agens, a zprostředkovávají produkci cytokinů, potřebných pro vývoj účinné imunity. TLR jsou vysoce konzervované od Drosophily až po člověka a vyznačují se funkční a strukturní podobností. Různé TLR vykazují rozdílné vzory exprese. Tento receptor je nejvíce exprimován na placentě, ve slezině a v myelomonocytické subpopulaci leukocytů.[3]
Je známě, že TLR4 kooperuje s proteiny LY96 (rovněž známým jako MD2) a CD14, aby došlo ke zprostředkování signálních transdukčních událostí vyvolaných lipopolysacharidem (LPS), který se nachází u většiny gamnegativních bakterií.[4] Mutace v tomto genu jsou spojovány s rozdíly v odpovědi vůči LPS. Signalizace u TLR4 probíhá tvorbou komplexu mezi extracelulární na leucinové repetice bohatou doménou a intracelulárním toll/interleukin-1 receptorovou doménou (TIR). Stimulace prostřednictvím LPS indukuje řadu interakcí s několika pomocnými proteiny, které tvoří TLR4 komplex na buněčném povrchu. Nejprve je LPS rozeznán vazbou na lipopolysacharid vázající protein (LBP). LPS-LBP komplex přenáší LPS dále na CD14. CD14 je protein s glykofosfatidylinositolovou (GPI) kotvou, zanořenou v membráně, který váže LPS-LBP komplex a usnadňuje přenos LPS na protein MD2, asociovaný s extracelulární doménou TLR4. Vazba LPS spouští dimerizaci MD2 s TLR4 a konformační změna TLR4 je pak rozhodující pro nábor adaptorových proteinů, nesoucích TIR domény, nezbytných pro přenos signálu uvnitř buňky.[5]
Bylo nalezeno několik transkripčních variant TLR4, ale jejich protein kódující potenciál zůstává u většiny neznámý.[6]
Signalizace
Po rozpoznání LPS vedou konformační změny u TLR4 k navázání intracelulárních TIR domén, které obsahují adaptorové proteiny. Tyto adaptory jsou spojeny s TLR4 prostřednictvím homofilních interakcí mezi doménami TIR. Existují čtyři adaptorové proteiny, zapojené do dvou hlavních intracelulárních signálních drah.[7]
MyD88 závislá dráha
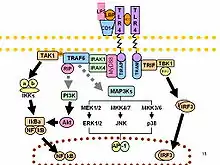
MyD88 závislá dráha je regulována prostřednictvím dvou proteinů asociovaných s adaptory: proteinem primární odpovědi myeloidní diferenciace 88 (MyD88) a proteinovým adaptérem obsahujícím IR doménu (TIRAP). TIRAP-MyD88 reguluje fázi časné aktivace NF-kB a produkci prozánětlivých cytokinů.[1] MyD88 signalizace zahrnuje aktivaci kináz asociovaných s IL-1 receptorem (IRAK) a adaptorového proteinů TRAF6. TRAF6 idukuje aktivaci TAK1 kinázy, která dále vede k aktivaci kaskád mitogenem aktivované proteinkinázy (MAP kinázy) a IκB kinázy (IKK). Signální dráha IKK vede k aktivaci transkripčního faktoru NF-kB, zatímco MAPK kaskády aktivují jiný transkripční faktor: AP-1. Avšak obě hrají roli v expresi prozánětlivých cytokinů.[5] Aktivace NF-kB prostřednictvím TAK-1 probíhá v proteinovém komplexu (signalosomu), tvořeném skafoldovým proteinem zvaným NEMO. Součástí signalosomu jsou dvě různé kB kinázy, které se nazývají IKKα a IKKβ. IKK-β fosforyluje IκB, což způsobí jeho degradaci a umožní jadernou translokaci transkripčního faktoru NF-kB.[8]
MyD88 nezávislá dráha
MyD88 nezávislá dráha je regulovaná prostřednictvím adaptorových proteinů TRIF, obsahujících TIR doménu, a TRAM. TRAM-TRIF prostřednictvím proteinu TRAF3 aktivují transkripční faktor IRF3, který indukuje produkci interferonů typu I.[7]
SARM: negativní regulátor TRIF zprostředkované dráhy
Pátým adaptorovým proteinem obsahujícím doménu TIR je Sterile α and HEAT (SARM). Jedná se však o inhibitor TLR4 signální dráhy. Aktivace SARM prostřednictvím vazby LPS inhibuje TRIF zprostředkovanou dráhu, avšak neinhibuje MyD88 závislou dráhu. Tento mechanismus slouží k regulaci nadměrné aktivace v reakci na LPS, která může vést k poškození vyvolanému zánětem, například k sepsi.[5]
Evoluční historie
Ke vzniku TLR4 došlo divergencí TLR2 a TLR4 před asi 500 miliony lety, v době těsně před začátkem evoluce obratlovců.[9] Porovnání sekvencí exonů TLR4 člověka a lidoopů prokázalo, že lidská varianta TLR4 se od našeho posledního společného předka s šimpanzi takřka nezměnila. Lidské a šimpanzí TLR4 exony se liší pouze třemi substitucemi, zatímco lidské a paviání jsou si v extracelulární doméně podobné z 93,5%.[10] Nicméně, lidé mají v TLR4 větší počet předčasných stop kodonů, než ostatní hominidé.Ve studii porovnávající 158 lidí z různých částí světa mělo 0,6% jedinců nonsense mutaci.[11][12] To naznačuje, že na lidský TLR4 působí slabší evoluční tlak než na nám příbuzné primáty. Distribuce lidských polymorfismů TLR4 odpovídá migraci mimo Afriku a je pravděpodobné, že tyto polymorfismy vznikly v Africe ještě před migrací na jiné kontinenty.[12][13]
Interakce
Bylo zjištěno, že TLR4 interaguje s:
- LY96 (MD2) - protein vázající lipopolysacharid[14]
- MyD88 - adaptorový protein[15]
- TOLLIP - inhibiční adaptorový protein[16]
- Niklem[17]
Intracelulární transport TLR4 je závislý na GTPáze Rab-11a. Knockdown Rab-11a má za následek omezený přesun TLR4 do fagosomů obsahujících E. coli a následné snížení transdukce signálu MyD88 nezávislou dráhou.[18]
Klinický význam
U lidské varianty TLR4 byla identifikována řada jednonukleotidových polymorfismů (SNP)[19] a u některých z nich byla zjištěna souvislost se zvýšenou citlivostí na gramnegativní bakteriální infekce[20] nebo s rychlejší progresí a závažnějším průběhem sepse u vážně nemocných pacientů.[21]
Inzulinová rezistence
Protein Fetuin-A, který patří mezi endogenní ligandy TLR4, usnadňuje vazbu lipidů na receptory, čímž vyvolává rezistenci na inzulin.[22]
Nádorová onemocnění
Exprese TLR4 může být detekována u mnoha nádorových buněk a buněčných linií. TLR4 aktivuje NF-kB a MAP kinázovou dráhu, což implikuje možnou přímou roli buněčně autonomní signalizace TLR4 při regulaci karcinogeneze, zejména prostřednictvím zvýšené proliferace nádorových buněk, inhibice apoptózy a metastázování. Signalizace TLR4 může také přispívat k rezistenci vůči paclitaxelu, který se používá jako cytostatikum v léčbě karcinomu prsu, a rezistenci vůči siRNA terapii, využívané v terapii karcinomu prostaty. U 63% pacientek trpících karcinonem prsu, byla detekována exprese TLR4 na buňkách karcinomu a úroveň exprese nepřímo korelovala s přežitím. Navíc, nízká exprese MyD88 korelovala se sníženou metastázou do plic a sníženou expresí chemokinů CCL2 a CCL5. Nejvyšší exprese TLR4 v porovnání s ostatními TLR byla zaznamenána u linie lidského karcinomu prsu MDA-MB-231 a knockdown TLR4 vedl ke snížení proliferace i snížení hladin IL-6 a IL-8. Ovšem TLR4 signalizace v imunitních buňkách nádorového mikroprostředí může vést jak k produkci prozánětlivých cytokinů (TNF, IL-1β, IL-6, IL-18, atd.), tak protizánětlivých cytokinů (IL-10, TGF-β, atd.). Zároveň může podporovat produkci angiogenních mediátorů (VEGF, EGF, TGF-β, atd.). Toto může působit na polarizaci makrofágů asociovaných s nádory, přeměnu fibroblastů na fibroblasty podporující nádor, také přeměnu dendritických buněk v pro-nádorové a v neposlední řadě pro-tumorigenních aktivaci nezralých myeloidních buněk a jejich přeměnu na myeloidní supresorové buňky (MDSC). TLR signalizace je spojována a akumulací a funkcí MDSC v místě nádoru a také se schopností mezenchymálních stromálních buněk čelit protinádorové imunitě zprostředkované NK buňkami. U HepG2 hepatoblastomových buněk LPS zvýšil hladiny TLR4, buněčnou proliferaci a rezistenci vůči chemoterapii. Tyto jevy bylo možné zvrátit knockdownem genu pro TLR4. LPS stimulace podobně působila i na buněčnou linii lidské rakoviny jater H7402, kde bylo možné projevy zvrátit inhibicí STAT3. Kromě úspěšného využití BCG pro terapii rakoviny močového měchýře, byly v léčbě skvamocelulárního karcinomu, karcinomu žaludku a karcinomu děložního hrdla zaznamenány úspěchy i při orální aplikaci streptokoků připravených lyofilizací se současným využitím ligandů TLR4 / TLR2 - derivátů muramyldipeptidu.[23]
Těhotenství
Aktivace TLR4 při intrauterinních infekcích vede k deregulaci syntézy prostaglandinů, což vede ke kontrakci hladkého svalstva dělohy.
Asp299Gly polymorfismus
Přestože je TLR4 tradičně známý jako receptor pro LPS, váže i řadu jiných lipidů. Původce nejčastější a nejzávažnější formy malárie převážně v Africe - Plasmodium falciparum, produkuje glykofosfatidylinositol, který může aktivovat TRL4.[24] Dva objevené SNP u TLR4 jsou koexprimovány s vysokou penetrací v afrických populacích: TLR-4-Asp299Gly and TLR-4-Thr399Ile. Tyto polymorfismy jsou spojeny se zvýšenou produkcí imunomodulačního IL-10 prostřednictvím TLR4 signalizace a následným snížením produkce prozánětlivých cytokinů.[25] Bodová mutace u TLR-4-Asp299Gly silně koreluje se zvýšenou infekcí Plasmodium falciparum, což naznačuje, že mutace brání TLR4 v silné odpovědi proti alespoň některým plasmodiálním infekcím. Míra infekce malárie a související morbidita jsou sice v populaci s TLR-4-Asp299Gly vyšší, zdá se však, že úmrtnost klesá. To může naznačovat, že malárie pro svou patogenezi alespoň částečně využívá produkce cytokinů. Díky snížení produkce cytokinů vlivem mutace sice může docházet ke zvýšené vnímavosti k infekci, ovšem zdá se, že úmrtnost se snižuje.[24]
Kromě toho je TLR4-D299G spojován s agresivním kolorektálním karcinomem u člověka. Ukázalo se, že lidské adenokarcinomy tlustého střeva pacientů s TLR4-D299G byly častěji v pokročilém stadiu s metastázami, než pacienti s jinou variantou. Stejná studie prokázala, že buňky střevního epitelu (Caco-2) exprimující TLR4-D299G prošly epiteliálně-mezenchymální tranzicí a morfologickými změnami spojenými s progresí nádoru, zatímco buňky střevního epitelu s obvyklou variantou TLR4 nikoliv.[26]
Studie se zvířaty
Během výzkumu možné souvislosti mezi TLR4 receptorem a nadměrnou konzumací alkoholu, bylo zjištěno, že po manipulaci genů zodpovědných za expresi TLR4 a GABA receptorů u hlodavců navyklých k nadměrnému pití, došlo u zvířat k výraznému snížení tohoto chování.[27] Navíc je známo, že ethanol může i bez přítomnosti LPS aktivovat signální dráhy TLR4.[28]
Vysoké hladiny TLR4 a M2 makrofágů asociovaných s nádory jsou spojeny s náchylností ke zvýšenému růstu nádorů u myší po spánkové deprivaci. Geneticky modifikované myši neprodukující TLR4 naopak vykazovaly normální růst rakoviny.[29]
Léčiva zacílená na TLR4
Bylo prokázáno, že TLR4 má významný podíl na dlouhodobých vedlejších účincích opioidních analgetik. Byly testovány různé ligandy μ-opioidního receptoru a bylo zjištěno, že působí zároveň jako agonisté i antagonisté TLR4, přičemž opioidní agonisté, jako je morfin, jsou také agonisté TLR4, zatímco opioidní antagonisté, jako je naloxon, byli shledáni antagonisty TLR4. Aktivace TLR4 vede k uvolňování prozánětlivých cytokinů včetně TNF-α a interleukinu 1 a předpokládá se, že konstantní uvolňování těchto mediátorů v nízké hladině po čase snižuje účinnost léčby opioidy a má podíl na vývoji tolerance na opioidní analgetika.[30][31] Vlivem dlouhodobého užívání opiodních léčiv dochází ke spuštění vedlejších účinků jako je hyperalgezie a alodynie.[32][33] Léčiva, která působí proti účinkům TNF-α a IL-1, zvyšují analgetické účinky opioidů a zároveň snižují rozvoj tolerance i ostatních vedlejších účinků. Stejné účinky byly zaznamenány při podání léčiv blokujících samotný TLR.[34][35]
Dále bylo zjištěno, že odpověď TLR4 na opioidní léčiva je nezávislá na enantiomeru, takže „nepřirozené“ enantiomery opioidních léčiv, jako je morfin a naloxon, které postrádají afinitu k opioidním receptorům, stále vykazují afinitu k TLR4.[36][37] Z toho vyplývá, že nepřirozené enantiomery opioidních antagonistů, jako je (+)-naloxon lze využít k blokování aktivity TLR4, zatímco analgetická aktivita zprostředkovaná μ-opioidním receptorem zůstává zachována.[37][38] To může být také důvodem příznivého účinku velmi nízké dávky naltrexonu na opioidní analgezii.[39]
Morfin působí prozánětlivě vazbou na protein MD2, který se následně vážně na TLR4.[40] Morfinem indukovaná aktivace TLR4 působí proti potlačování bolesti opioidy a zvyšuje vývoj tolerance a závislosti na opioidech, má vliv na zneužívání drog a podílí se na dalších negativních účincích, jako je respirační deprese a hyperalgezie. Léčiva zacílená na TLR4 by mohla léčbu bolesti na bázi opioidů zlepšit.[41]
Agonisté
Antagonisté
- Eritoran - analog lipidu A
Od prosince 2009 byl vyvíjen jako lék proti těžké sepsi.[47] Avšak později bylo zjištěno, že jeho účinky jsou nedostačující a bylo navrženo jiné využití - v léčbě některých závažných případů chřipky. Ačkoli neléčí samotný virus, lze jej použít proti masivní imunitní reakci zvané cytokinová bouře, která je hlavní příčinou úmrtnosti na těžkou chřipku.[48]
Reference
V tomto článku byl použit překlad textu z článku TLR4 na anglické Wikipedii.
- VAURE, Céline; LIU, Yuanqing. A Comparative Review of Toll-Like Receptor 4 Expression and Functionality in Different Animal Species. Frontiers in Immunology. 2014-07-10, roč. 5. Dostupné online [cit. 2020-01-19]. ISSN 1664-3224. DOI 10.3389/fimmu.2014.00316. PMID 25071777.
- BRUBAKER, Sky W.; BONHAM, Kevin S.; ZANONI, Ivan. Innate Immune Pattern Recognition: A Cell Biological Perspective. Annual Review of Immunology. 2015-03-21, roč. 33, čís. 1, s. 257–290. Dostupné online [cit. 2020-01-19]. ISSN 0732-0582. DOI 10.1146/annurev-immunol-032414-112240. PMID 25581309. (anglicky)
- TLR4 toll like receptor 4 [Homo sapiens (human)] - Gene - NCBI. www.ncbi.nlm.nih.gov [online]. [cit. 2020-01-19]. Dostupné online.
- PARK, B.S.; SONG, D.H.; KIM, H.M. Crystal structure of the human TLR4-human MD-2-E.coli LPS Ra complex. dx.doi.org [online]. 2009-03-03 [cit. 2020-01-19]. Dostupné online.
- LU, Yong-Chen; YEH, Wen-Chen; OHASHI, Pamela S. LPS/TLR4 signal transduction pathway. Cytokine. 2008-05, roč. 42, čís. 2, s. 145–151. Dostupné online [cit. 2020-01-19]. DOI 10.1016/j.cyto.2008.01.006. (anglicky)
- JULIANA, Bortolatto. Toll-like receptor 4 (TLR4) na modulação da imunidade do tipo 2.. , . Universidade de Sao Paulo Sistema Integrado de Bibliotecas - SIBiUSP. . Dostupné online.
- O'NEILL, Luke A. J.; GOLENBOCK, Douglas; BOWIE, Andrew G. The history of Toll-like receptors — redefining innate immunity. Nature Reviews Immunology. 2013-06, roč. 13, čís. 6, s. 453–460. Dostupné online [cit. 2020-01-20]. ISSN 1474-1733. DOI 10.1038/nri3446. (anglicky)
- PALSSON-MCDERMOTT, Eva M.; O'NEILL, Luke A. J. Signal transduction by the lipopolysaccharide receptor, Toll-like receptor-4. Immunology. 2004-10, roč. 113, čís. 2, s. 153–162. Dostupné online [cit. 2020-01-20]. ISSN 0019-2805. DOI 10.1111/j.1365-2567.2004.01976.x. PMID 15379975. (anglicky)
- BEUTLER, B.; REHLI, M. Evolution of the TIR, Tolls and TLRs: Functional Inferences from Computational Biology. Příprava vydání Bruce Beutler, Hermann Wagner. Svazek 270. Berlin, Heidelberg: Springer Berlin Heidelberg Dostupné online. ISBN 978-3-642-63975-3, ISBN 978-3-642-59430-4. DOI 10.1007/978-3-642-59430-4_1. S. 1–21. DOI: 10.1007/978-3-642-59430-4_1.
- SMIRNOVA, Irina; POLTORAK, Alexander; CHAN, Edward KL. [No title found]. Genome Biology. 2000, roč. 1, čís. 1, s. research002.1. Dostupné online [cit. 2020-01-20]. DOI 10.1186/gb-2000-1-1-research002. PMID 11104518.
- QUACH, Hélène; WILSON, Daniel; LAVAL, Guillaume. Different selective pressures shape the evolution of Toll-like receptors in human and African great ape populations. Human Molecular Genetics. 2013-12-01, roč. 22, čís. 23, s. 4829–4840. Dostupné online [cit. 2020-01-20]. ISSN 1460-2083. DOI 10.1093/hmg/ddt335. PMID 23851028. (anglicky)
- BARREIRO, Luis B.; BEN-ALI, Meriem; QUACH, Hélène. Evolutionary Dynamics of Human Toll-Like Receptors and Their Different Contributions to Host Defense. PLoS Genetics. 2009-07-17, roč. 5, čís. 7, s. e1000562. Dostupné online [cit. 2020-01-20]. ISSN 1553-7404. DOI 10.1371/journal.pgen.1000562. PMID 19609346. (anglicky)
- PLANTINGA, Theo S.; IOANA, Mihai; ALONSO, Santos. The Evolutionary History of TLR4 Polymorphisms in Europe. Journal of Innate Immunity. 2012, roč. 4, čís. 2, s. 168–175. Dostupné online [cit. 2020-01-20]. ISSN 1662-811X. DOI 10.1159/000329492. PMID 21968286. (anglicky)
- RE, Fabio; STROMINGER, Jack L. Monomeric Recombinant MD-2 Binds Toll-like Receptor 4 Tightly and Confers Lipopolysaccharide Responsiveness. Journal of Biological Chemistry. 2002-06-28, roč. 277, čís. 26, s. 23427–23432. Dostupné online [cit. 2020-01-20]. ISSN 0021-9258. DOI 10.1074/jbc.M202554200. (anglicky)
- FITZGERALD, Katherine A.; PALSSON-MCDERMOTT, Eva M.; BOWIE, Andrew G. Mal (MyD88-adapter-like) is required for Toll-like receptor-4 signal transduction. Nature. 2001-09, roč. 413, čís. 6851, s. 78–83. Dostupné online [cit. 2020-01-20]. ISSN 0028-0836. DOI 10.1038/35092578. (anglicky)
- ZHANG, Guolong; GHOSH, Sankar. Negative Regulation of Toll-like Receptor-mediated Signaling by Tollip. Journal of Biological Chemistry. 2002-03-01, roč. 277, čís. 9, s. 7059–7065. Dostupné online [cit. 2020-01-20]. ISSN 0021-9258. DOI 10.1074/jbc.M109537200. (anglicky)
- PEANA, Massimiliano; ZDYB, Karolina; MEDICI, Serenella. Ni(II) interaction with a peptide model of the human TLR4 ectodomain. Journal of Trace Elements in Medicine and Biology. 2017-12, roč. 44, s. 151–160. Dostupné online [cit. 2020-01-20]. DOI 10.1016/j.jtemb.2017.07.006. (anglicky)
- HUSEBYE, Harald; AUNE, Marie Hjelmseth; STENVIK, Jørgen. The Rab11a GTPase Controls Toll-like Receptor 4-Induced Activation of Interferon Regulatory Factor-3 on Phagosomes. Immunity. 2010-10, roč. 33, čís. 4, s. 583–596. Dostupné online [cit. 2020-01-20]. DOI 10.1016/j.immuni.2010.09.010. (anglicky)
- SCHRÖDER, Nicolas WJ; SCHUMANN, Ralf R. Single nucleotide polymorphisms of Toll-like receptors and susceptibility to infectious disease. The Lancet Infectious Diseases. 2005-03, roč. 5, čís. 3, s. 156–164. Dostupné online [cit. 2020-01-20]. DOI 10.1016/S1473-3099(05)01308-3. (anglicky)
- LORENZ, Eva; MIRA, Jean Paul; FREES, Kathy L. Relevance of Mutations in the TLR4 Receptor in Patients With Gram-Negative Septic Shock. Archives of Internal Medicine. 2002-05-13, roč. 162, čís. 9, s. 1028. Dostupné online [cit. 2020-01-20]. ISSN 0003-9926. DOI 10.1001/archinte.162.9.1028. (anglicky)
- NACHTIGALL, Irit; TAMARKIN, Andrey; TAFELSKI, Sascha. Polymorphisms of the toll-like receptor 2 and 4 genes are associated with faster progression and a more severe course of sepsis in critically ill patients. Journal of International Medical Research. 2014-02, roč. 42, čís. 1, s. 93–110. Dostupné online [cit. 2020-01-20]. ISSN 0300-0605. DOI 10.1177/0300060513504358. (anglicky)
- PAL, Durba; DASGUPTA, Suman; KUNDU, Rakesh. Fetuin-A acts as an endogenous ligand of TLR4 to promote lipid-induced insulin resistance. Nature Medicine. 2012-08, roč. 18, čís. 8, s. 1279–1285. Dostupné online [cit. 2020-01-20]. ISSN 1078-8956. DOI 10.1038/nm.2851. (anglicky)
- KORNEEV, Kirill V.; ATRETKHANY, Kamar-Sulu N.; DRUTSKAYA, Marina S. TLR-signaling and proinflammatory cytokines as drivers of tumorigenesis. Cytokine. 2017-01, roč. 89, s. 127–135. Dostupné online [cit. 2020-01-21]. DOI 10.1016/j.cyto.2016.01.021. (anglicky)
- MOCKENHAUPT, F. P.; CRAMER, J. P.; HAMANN, L. Toll-like receptor (TLR) polymorphisms in African children: Common TLR-4 variants predispose to severe malaria. Proceedings of the National Academy of Sciences. 2006-01-03, roč. 103, čís. 1, s. 177–182. Dostupné online [cit. 2020-01-21]. ISSN 0027-8424. DOI 10.1073/pnas.0506803102. PMID 16371473. (anglicky)
- VAN DER GRAAF, Chantal A. A.; NETEA, Mihai G.; MORRÉ, Servaas A. Toll-like receptor 4 Asp299Gly/Thr399Ile polymorphisms are a risk factor for Candida bloodstream infection. European Cytokine Network. 2006-03, roč. 17, čís. 1, s. 29–34. PMID: 16613760. Dostupné online [cit. 2020-01-21]. ISSN 1148-5493. PMID 16613760.
- EYKING, Annette; EY, Birgit; RÜNZI, Michael. Toll-like Receptor 4 Variant D299G Induces Features of Neoplastic Progression in Caco-2 Intestinal Cells and Is Associated With Advanced Human Colon Cancer. Gastroenterology. 2011-12, roč. 141, čís. 6, s. 2154–2165. Dostupné online [cit. 2020-01-22]. DOI 10.1053/j.gastro.2011.08.043. PMID 21920464. (anglicky)
- LIU, J.; YANG, A. R.; KELLY, T. Binge alcohol drinking is associated with GABAA 2-regulated Toll-like receptor 4 (TLR4) expression in the central amygdala. Proceedings of the National Academy of Sciences. 2011-03-15, roč. 108, čís. 11, s. 4465–4470. Dostupné online [cit. 2020-01-20]. ISSN 0027-8424. DOI 10.1073/pnas.1019020108. PMID 21368176. (anglicky)
- BLANCO, Ana M.; VALLÉS, Soraya L.; PASCUAL, Maria. Involvement of TLR4/Type I IL-1 Receptor Signaling in the Induction of Inflammatory Mediators and Cell Death Induced by Ethanol in Cultured Astrocytes. The Journal of Immunology. 2005-11-15, roč. 175, čís. 10, s. 6893–6899. Dostupné online [cit. 2020-01-20]. ISSN 0022-1767. DOI 10.4049/jimmunol.175.10.6893. (anglicky)
- HAKIM, F.; WANG, Y.; ZHANG, S. X. L. Fragmented Sleep Accelerates Tumor Growth and Progression through Recruitment of Tumor-Associated Macrophages and TLR4 Signaling. Cancer Research. 2014-03-01, roč. 74, čís. 5, s. 1329–1337. Dostupné online [cit. 2020-01-20]. ISSN 0008-5472. DOI 10.1158/0008-5472.CAN-13-3014. PMID 24448240. (anglicky)
- SHAVIT, Yehuda; WOLF, Gilly; GOSHEN, Inbal. Interleukin-1 antagonizes morphine analgesia and underlies morphine tolerance:. Pain. 2005-05, roč. 115, čís. 1, s. 50–59. Dostupné online [cit. 2020-01-22]. ISSN 0304-3959. DOI 10.1016/j.pain.2005.02.003. (anglicky)
- MOHAN, Shekher; DAVIS, Randall L.; DESILVA, Udaya. Dual regulation of mu opioid receptors in SK-N-SH neuroblastoma cells by morphine and interleukin-1β: Evidence for opioid-immune crosstalk. Journal of Neuroimmunology. 2010-10, roč. 227, čís. 1–2, s. 26–34. Dostupné online [cit. 2020-01-22]. DOI 10.1016/j.jneuroim.2010.06.007. PMID 20615556. (anglicky)
- KOMATSU, Takaaki; SAKURADA, Shinobu; KATSUYAMA, Sou. Chapter 16 Mechanism of Allodynia Evoked by Intrathecal Morphine‐3‐Glucuronide in Mice. Svazek 85. [s.l.]: Elsevier Dostupné online. ISBN 978-0-12-374893-5. DOI 10.1016/s0074-7742(09)85016-2. S. 207–219. (anglicky) DOI: 10.1016/S0074-7742(09)85016-2.
- LEWIS, S.S.; HUTCHINSON, M.R.; REZVANI, N. Evidence that intrathecal morphine-3-glucuronide may cause pain enhancement via toll-like receptor 4/MD-2 and interleukin-1β. Neuroscience. 2010-01, roč. 165, čís. 2, s. 569–583. Dostupné online [cit. 2020-01-22]. DOI 10.1016/j.neuroscience.2009.10.011. PMID 19833175. (anglicky)
- SHEN, Ching-Hui; TSAI, Ru-Yin; SHIH, Meng-Shen. Etanercept Restores the Antinociceptive Effect of Morphine and Suppresses Spinal Neuroinflammation in Morphine-Tolerant Rats:. Anesthesia & Analgesia. 2011-02, roč. 112, čís. 2, s. 454–459. Dostupné online [cit. 2020-01-22]. ISSN 0003-2999. DOI 10.1213/ANE.0b013e3182025b15. (anglicky)
- HOOK, Michelle A.; WASHBURN, Stephanie N.; MORENO, Georgina. An IL-1 receptor antagonist blocks a morphine-induced attenuation of locomotor recovery after spinal cord injury. Brain, Behavior, and Immunity. 2011-02, roč. 25, čís. 2, s. 349–359. Dostupné online [cit. 2020-01-22]. DOI 10.1016/j.bbi.2010.10.018. PMID 20974246. (anglicky)
- WATKINS, Linda R.; HUTCHINSON, Mark R.; RICE, Kenner C. The “Toll” of Opioid-Induced Glial Activation: Improving the Clinical Efficacy of Opioids by Targeting Glia. Trends in Pharmacological Sciences. 2009-11, roč. 30, čís. 11, s. 581–591. Dostupné online [cit. 2020-01-22]. DOI 10.1016/j.tips.2009.08.002. PMID 19762094. (anglicky)
- HUTCHINSON, Mark R.; ZHANG, Yingning; BROWN, Kimberley. Non-stereoselective reversal of neuropathic pain by naloxone and naltrexone: involvement of toll-like receptor 4 (TLR4): Naloxone reverses neuropathic pain via TLR4. European Journal of Neuroscience. 2008-07, roč. 28, čís. 1, s. 20–29. Dostupné online [cit. 2020-01-22]. DOI 10.1111/j.1460-9568.2008.06321.x. PMID 18662331. (anglicky)
- HUTCHINSON, Mark R.; COATS, Benjamen D.; LEWIS, Susannah S. Proinflammatory cytokines oppose opioid-induced acute and chronic analgesia. Brain, Behavior, and Immunity. 2008-11, roč. 22, čís. 8, s. 1178–1189. Dostupné online [cit. 2020-01-22]. DOI 10.1016/j.bbi.2008.05.004. PMID 18599265. (anglicky)
- LIN, Shinn-Long; TSAI, Ru-Yin; TAI, Yueh-Hua. Ultra-low dose naloxone upregulates interleukin-10 expression and suppresses neuroinflammation in morphine-tolerant rat spinal cords. Behavioural Brain Research. 2010-02-11, roč. 207, čís. 1, s. 30–36. Dostupné online [cit. 2020-01-22]. DOI 10.1016/j.bbr.2009.09.034. (anglicky)
- Making morphine work better. Nature. 2012-04, roč. 484, čís. 7395, s. 419–419. Dostupné online [cit. 2020-01-22]. ISSN 0028-0836. DOI 10.1038/484419a. (anglicky)
- CARMEN DRAHL. Small Molecules Target Toll-Like Receptors | Chemical & Engineering News. cen.acs.org [online]. [cit. 2020-01-22]. Dostupné online.
- HUTCHINSON, Mark R.; ZHANG, Yingning; SHRIDHAR, Mitesh. Evidence that opioids may have toll-like receptor 4 and MD-2 effects. Brain, Behavior, and Immunity. 2010-01, roč. 24, čís. 1, s. 83–95. Dostupné online [cit. 2020-01-22]. DOI 10.1016/j.bbi.2009.08.004. PMID 19679181. (anglicky)
- PASCUAL, María; BALIÑO, Pablo; ALFONSO-LOECHES, Silvia. Impact of TLR4 on behavioral and cognitive dysfunctions associated with alcohol-induced neuroinflammatory damage. Brain, Behavior, and Immunity. 2011-06, roč. 25, s. S80–S91. Dostupné online [cit. 2020-01-22]. DOI 10.1016/j.bbi.2011.02.012. (anglicky)
- HARRIS, Shelley A.; SOLOMON, Keith R. Percutaneous penetration of 2,4‐dichlorophenoxyacetlc acid and 2,4‐d dimethylamine salt in human volunteers. Journal of Toxicology and Environmental Health. 1992-07, roč. 36, čís. 3, s. 233–240. Dostupné online [cit. 2020-01-22]. ISSN 0098-4108. DOI 10.1080/15287399209531634. (anglicky)
- HUTCHINSON, M.R.; LORAM, L.C.; ZHANG, Y. Evidence that tricyclic small molecules may possess toll-like receptor and myeloid differentiation protein 2 activity. Neuroscience. 2010-06, roč. 168, čís. 2, s. 551–563. Dostupné online [cit. 2020-01-22]. DOI 10.1016/j.neuroscience.2010.03.067. PMID 20381591. (anglicky)
- KELLEY, Keith W.; DANTZER, Robert. Alcoholism and inflammation: Neuroimmunology of behavioral and mood disorders. Brain, Behavior, and Immunity. 2011-06, roč. 25, s. S13–S20. Dostupné online [cit. 2020-01-22]. DOI 10.1016/j.bbi.2010.12.013. PMID 21193024. (anglicky)
- TIDSWELL, Mark; TILLIS, William; LAROSA, Steven P. Phase 2 trial of eritoran tetrasodium (E5564), a Toll-like receptor 4 antagonist, in patients with severe sepsis*:. Critical Care Medicine. 2010-01, roč. 38, čís. 1, s. 72–83. Dostupné online [cit. 2020-01-22]. ISSN 0090-3493. DOI 10.1097/CCM.0b013e3181b07b78. (anglicky)
- New single-dose drug shortens flu symptoms. AccessScience [online]. [cit. 2020-01-22]. Dostupné online.
- JIA, Ze-jun; WU, Fei-xiang; HUANG, Qing-hai. [Toll-like receptor 4: the potential therapeutic target for neuropathic pain]. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. Acta Academiae Medicinae Sinicae. 2012-04, roč. 34, čís. 2, s. 168–173. PMID: 22776604. Dostupné online [cit. 2020-01-22]. ISSN 1000-503X. DOI 10.3881/j.issn.1000-503X.2012.02.013. PMID 22776604.
- LAN, Xi; HAN, Xiaoning; LI, Qian. Pinocembrin protects hemorrhagic brain primarily by inhibiting toll-like receptor 4 and reducing M1 phenotype microglia. Brain, Behavior, and Immunity. 2017-03, roč. 61, s. 326–339. Dostupné online [cit. 2020-01-22]. DOI 10.1016/j.bbi.2016.12.012. PMID 28007523. (anglicky)
- IMPELLIZZERI, D; CAMPOLO, M; DI PAOLA, R. Ultramicronized palmitoylethanolamide reduces inflammation an a Th1-mediated model of colitis. European Journal of Inflammation. 2015-04, roč. 13, čís. 1, s. 14–31. Dostupné online [cit. 2020-01-22]. ISSN 1721-727X. DOI 10.1177/1721727X15575869. (anglicky)