Protein teplotního šoku
Protein teplotního šoku (hsp – heat shock protein) je označení pro skupinu proteinů, které zajišťují univerzální stresovou odpověď buněk, například při vystavení buňky vysoké teplotě, extrémnímu pH nebo třeba těžkým kovům. Váží se na nesbalené proteiny, chrání je či pomáhají jejich sbalení (chaperonová aktivita).[1]
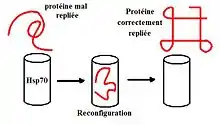
Hsp se vyskytují jak u prokaryot, tak u eukaryot (včetně člověka). U eukaryot se obvykle dělí do tří skupin, Hsp60, Hsp70 a Hsp90, a to podle jejich velikosti.[2] V ER se vyskytuje třeba BiP, v mitochondriích je Grp75 a v cytosolu bakterií DnaK.
Funkce HSP v imunitě
Význam HSP pro imunitní systém je založen na jejich schopnosti vázat nejen proteiny, ale také peptidy. Mohou tak přenášet peptidy určené k antigenní prezentaci. Vazba peptidu na HSP je typicky nízkoafinní a s nízkou specifitou [3]. Vazebné místo pro peptidy bylo detekováno u hsp70, hsp90, gp96 a kalretikulinu[3].
HSP dále stimulují receptory imunitního systému a podílejí se na správném sbalení celé řady imunologicky významných proteinů[4][5]. Například gp96 se podílí na sbalování TLR a integrinů[3].
Význam v antigenní prezentaci
HSP jsou nedílnou součástí procesu antigenní prezentace - ať už klasické MHCI či MHCII dráhy[3][6][7], zkřížené prezentace[4], nebo autofagie[7].
Klasická MHCI prezentace
Ve zkratce klasická MHCI prezentace je dráha, kdy antigenní peptidy vzniklé v proteazomu jsou transportovány skrz TAP protein do endoplasmatického retikula a zde naloženy na MHCI molekuly. Ty následně putují i s peptidy sekreční drahou na plasmatickou membránu. Při takto zjednodušeném popisu klasické MHCI prezentace jsou HSP obvykle zanedbávány, hrají ale významnou roli v účinnosti celého procesu[3].
HSP vážou poškozené buněčné proteiny a navádí je do proteasomu. Následně vážou vzniklé peptidy a přenáší je k membráně endoplasmatického retikula k TAP transportéru. Hsp90 a hsp70 si peptidy pravděpodobně předávají, kdy hsp90 asociuje s proteazomem a čeká na vzniklý peptid, hsp70 asociuje s TAP transportérem a navzájem mohou hsp90 a hsp70 interagovat spolu navzájem[3]. Uvnitř ER jsou pak další chaperony, tvoří společně s TAP a MHCI takzvaný "peptide loading complex" (v překladu komplex nakládající peptidy na MHCI). Peptidy v ER váže hlavně kalretikulin, gp96 více napomáhá nasednutí vysokoafinního peptidu do žlábku MHCI molekuly[3].
Funkce HSP při antigenní prezentaci má své opodstatnění. Vznikající peptidy mohou obsahovat hydrofobní aminokyseliny, které je třeba ve vodném prostředí cytosolu skrýt. Zároveň pouhá difuze peptidů by byla velmi neefektivní a snižovala by účinnost celé prezentace[3].
Klasická MHCII prezentace
Též v rámci dráhy MHCII prezentace se HSP uplatňují. Podílejí se na průběhu klatrinové endocytózy a maturaci váčků[7]. Pokud se HSP dostanou do extracelulárního prostředí, mohou při pohlcení antigen-prezentující buňkou navádět vázané peptidy do MHCII dráhy. Zde však není vůbec zřejmé, jakým způsobem je voleno mezi dráhou MHCII a zkříženou prezentací (viz níže)[4].
Autofagie
Autofagická dráha umožňuje prezentaci intracelulárních antigenních peptidů na MHCII molekule. HSP se podílejí na klasické makroautofagii, kdy proteinové agregáty či celé organely jsou obklopeny dvojitou membránou a následně degradovány při fůzi s lyzozomem[7]. Dále se HSP účastní speciálního typu autofagie, bývá označován jako chaperony mediovaná autofagie (CMA), kdy HSP napomáhá transportu cytosolických proteinů přímo do lyzozomů[7].
Zkřížená prezentace
Zkřížená prezentace je dráha umožňující prezentaci extracelulárních antigenů na MHCI. Pokud se HSP dostanou do extracelulárního prostředí, mohou se vázat na povrchové receptory dendritických buněk a indukovat pohlcení a zkříženou prezentaci nesených peptidů. Pro nasměrování do zkřížené prezentace jsou zásadní scavenger receptory LOX-1 a SRECI[4]. Dříve byl za hlavní HSP receptor považován CD91, jeho úloha však byla v poslední době zpochybněna, neboť není výrazně exprimován na většině dendritických buněk a u řady HSP se nepodařilo prokázat vazbu[4].
LOX-1 váže hlavně hsp60 a hsp70, SRECI váže prakticky všechny imunitně významné HSP[4].
Uplatnění HSP ve zkřížené prezentaci významně zvyšuje její účinnost[4][3]. HSP chrání navázané peptidy při transportu buněčnými kompartmenty před degradací. Zároveň jsou peptidy v komplexu s HSP efektivněji pohlcovány, než kdyby byly solubilní. Vysoká účinnost zkřížené prezentace je důležitá zejména při protinádorové imunitě. Množství nádorových antigenů, vůči nimž může být namířena imunitní odpověď je totiž omezené. Pokud má být imunitní systém dostatečně aktivován, dendritické buňky musí prezentovat co největší množství peptidů. Samozřejmě je třeba aktivovat nejen cytotoxické T lymfocyty, ale také pomocné Th1 lymfocyty a extracelulární HSP mohou navádět vázané peptidy jak na MHCI, tak na MHCII. Jakým způsobem však dochází k rozhodnutí mezi těmito dvěma drahami není zatím jasné[3].
HSP hrají roli i intracelulárně při zkřížené prezentaci. Cytosolická cesta zkřížené prezentace zahrnuje transport antigenních peptidů z endosomu do cytosolu skrze membránový pór. Na tomto transportu se podílí i hsp70 a hsp90[3].
HSP jako DAMPy
Pokud se HSP octnou v extracelulárním protstředí, mohou být naším imunitním systémem vnímany jako tzv. danger associated molecular patterns (DAMP)[4]. Jedná se o struktury, které stimulují PRR podobně jako různé bakteriální produkty, ale DAMP jsou odvozené z našich vlastních, endogenních struktur.
HSP mohou interagovat s TLR2 a TLR4 a aktivovat tak antigen prezentující buňky, tzn. indukovat u nich expresi kostimulačních molekul, prozánětlivých a polarizačních cytokinů a MHC molekul[6][3].
HSP mohou signalizovat také přes scavenger receptory. Některé asociují s TLR, jiné přímo spouští prozánětivé dráhy v buňce (MAPK, NF-kB). Výjimkou je v tomto případě scavenger receptor SRA, který po stimulaci působí imunosupresivně[3].
Jak se HSP dostávají do extracelulárního prostředí
HSP mohou být sekretovány z nádorových a imunitních buněk za stresových podmínek skrze nekanonické sekreční dráhy, podobně jako IL1b, neboť nemají signální peptid, který by je naváděl do klasické sekreční dráhy[4].
Další možností je vylití HSP při buněčné nekróze, nebo sekrece prostřednictvím exosomů[4]. Také při některých speciálních typech apoptotické smrti (např. indukované chemoterapeutiky) dochází k vystavení HSP na extracelulární straně plasmatické membrány[6].
Jak dlouhou dokážou HSP asociovat s peptidy v extracelulárním prostředí je terčem debat. Alespoň v případě hsp70 se zdá, že vazba peptidu by měla být dostatečně stabilní, aby HSP mohl peptid přenést až k antigen prezentující buňce[4].
Působení extracelulárních HSP na imunitní systém je rozmanité. Mohou podporovat antigenní prezentaci a stimulovat řadu receptorů denritických buněk. Výsledný imunostimulační (Th1, Th17, Th2) či imunosupresivní (Treg) efekt je výrazně závislý na kontextu tkáně. HSP dovedou v principu indukovat obojí prostřednictvím tolerogeních či imunogeních dendritických buněk[3].
V důsledku klinické využití HSP zahrnuje jak protinádorovou terapii tak i terapii autoimunitních onemocnění, jako je revmatoidní artritida, či diabetes 1. typu[3][8].
Reference
- http://biomikro.vscht.cz/documents/biochemie_mikroorganismu/pdf/HSP.pdf%5B%5D
- Oxford dictionary of biochemistry and molecular biology; revised edition. Příprava vydání R. Cammack et al. New York: Oxford university press, 2006. ISBN 0-19-852917-1.
- BINDER, Robert Julian. Functions of Heat Shock Proteins in Pathways of the Innate and Adaptive Immune System. The Journal of Immunology. 2014-12-15, roč. 193, čís. 12, s. 5765–5771. PMID: 25480955. Dostupné online [cit. 2018-06-13]. ISSN 0022-1767. DOI 10.4049/jimmunol.1401417. PMID 25480955. (anglicky)
- MURSHID, Ayesha; GONG, Jianlin; CALDERWOOD, Stuart Keith. The Role of Heat Shock Proteins in Antigen Cross Presentation. Frontiers in Immunology. 2012, roč. 3. Dostupné online [cit. 2018-06-13]. ISSN 1664-3224. DOI 10.3389/fimmu.2012.00063. PMID 22566944. (English)
- TUKAJ, Stefan; WĘGRZYN, Grzegorz. Anti-Hsp90 therapy in autoimmune and inflammatory diseases: a review of preclinical studies. Cell Stress and Chaperones. 2016-01-20, roč. 21, čís. 2, s. 213–218. Dostupné online [cit. 2018-06-13]. ISSN 1355-8145. DOI 10.1007/s12192-016-0670-z. PMID 26786410. (anglicky)
- GRANER, Michael W. HSP90 and Immune Modulation in Cancer. [s.l.]: Elsevier Dostupné online. ISBN 9780128022900. DOI 10.1016/bs.acr.2015.10.001. S. 191–224.
- DEFFIT, Sarah N.; BLUM, Janice S. A central role for HSC70 in regulating antigen trafficking and MHC class II presentation. Molecular Immunology. 2015-12, roč. 68, čís. 2, s. 85–88. Dostupné online [cit. 2018-06-13]. ISSN 0161-5890. DOI 10.1016/j.molimm.2015.04.007. PMID 25953005.
- JANSEN, Manon A. A.; SPIERING, Rachel; BROERE, Femke. Targeting of tolerogenic dendritic cells towards heat-shock proteins: a novel therapeutic strategy for autoimmune diseases?. Immunology. 2017-09-18, roč. 153, čís. 1, s. 51–59. Dostupné online [cit. 2018-06-13]. ISSN 0019-2805. DOI 10.1111/imm.12811. PMID 28804903. (anglicky)
Externí odkazy
 Obrázky, zvuky či videa k tématu Protein teplotního šoku na Wikimedia Commons
Obrázky, zvuky či videa k tématu Protein teplotního šoku na Wikimedia Commons