T-buněčný receptor
T-buněčný receptor (TCR, z anglického T-cell receptor) je molekula nacházející se na povrchu T-lymfocytů. Ve většině případů TCR rozpoznává antigenní peptidy prezentované na molekulách hlavního histokompatibilního komplexu (MHC). Signalizace přes TCR a s ním asociované molekuly (CD3 komplex, koreceptory) vede k aktivaci T-buňky a odpovědi na příslušný ligand.
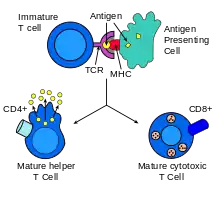
Struktura
T-buněčný receptor svou strukturou připomíná Fab fragmenty imunoglobulinů. TCR je transmembránový heterodimer složený z řetězců α a β popřípadě γ a δ. Na každém řetězci lze rozlišit domény extracelulární (ta je největší), transmembránovou a intracelulární. Extracelulární část se nadále dělí na variabilní oblast (V), konstantní oblast (C) a stonkový segment. Variabilní oblast všech řetězců obsahuje tři hypervariabilní smyčky zvané CDR (Complemetarity Determining Regions). CDR oblasti jsou hlavními prostředníky interakce TCR s komplexem peptid:MHC (viz dále). Řetězec β navíc obsahuje čtvrtou hypervariabilní smyčku HV4.
Konstantní domény obou řetězců spolu pravděpodobně interagují prostřednictvím uhlovodíkových zbytků z navázaných cukrů. Stonkový segment je charakteristický bohatým zastoupením cysteinových zbytků, díky nimž dochází ke vzniku disulfidických můstků mezi oběma řetězci heterodimeru.
Rozdíl ve struktuře mezi receptory αβ a γδ je v úhlu, který svírají podélné osy variabilní a konstantní oblasti. Zatímco u γδ TCR je tento úhel přibližně 111°, úhel u αβ TCR je větší: 147°. Je možné, že tento rozdíl se může podílet na rozdílném přenosu signálu do buňky a odlišné interakci s koreceptory.
Přestavba genů pro TCR
TCR v genomu
TCRα lokus leží na 14. chromozomu a obsahuje V (variable) a J (joining) genové segmenty (Vα, Jα). Lidské Vα obsahuje 70-80 segmentů, každému z nich předchází exon kódující vedoucí (Leader, L) sekvenci. L sekvence je po vstupu do endoplazmatického retikula (ER) odštípnuta.Za genovými segmenty kódující variabilní oblast leží shluk přibližně 60 segmentů kódujících Jα část. Konstantní část (Cα) je kódována jedním segmentem.
TCRβ lokus leží na 7. chromozomu (u myši na 6. chromosomu) a je jinak organizován. Kromě Vβ a Jβ genových segmentů jsou součástí genu i segmenty kódující Dβ segmenty (diversity). Na chromozomu jsou genové segmenty uspořádány následovně: přibližně 52 funkčních Vβ segmentů je lokalizováno daleko od dvou separátních shluků (klastrů), z nichž každý obsahuje 1 Dβ, 6-7 Jβ a 1 Cβ.
V případě řetězců α a β je CDR3 smyčka kódována segmenty D a J. Smyčky CDR1 a 2 segmentem V. CDR1 a CDR2 jsou méně variabilní, kontaktují méně variabilní oblast komplexu peptid:MHC, zatímco vysoce variabilní CDR3 oblast interaguje s unikátní peptidovou složkou TCR ligandu.
TCRδ lokus leží mezi genovými segmenty Vα a Jα. Toto umístění způsobuje, že jakákoliv přestavba v α lokusu vede ke ztrátě δ lokusu. TCRδ lokus obsahuje genové segmenty V, D i J.
TCRγ lokus leží na 7. chromozomu (u myši na 13. chromozomu). TCRγ lokus kóduje genové segmenty V a J. Počet jednotlivých typů genových segmentů v lokusech γ i δ se pohybuje v řádu jednotek (výjimkou je Vγ segment se 14 variantami). Odtud tedy pramení menší diverzita γδ TCR, která je však zvyšována adicí P- a N- nukleotidů.
Genová přestavba
Přestavba TCR se odehrává v thymu (viz dále) a je homologní s přestavbou BCR/Ig. Celý proces je označován jako somatická rekombinace. Každý z TCR genových segmentů je ohraničen tzv. Recombination signal sequences (RSSs). RSSs jsou tvořeny heptamerem (CACAGTG), spacerem (o délce 12 nebo 23 páru bází) a nonamerem (ACAAAAACC). RSSs jsou rozeznávány stejnými enzymy jako RSSs ohraničující genové segmenty genů pro BCR/Ig. Při genových přestavbách je ve většině případů dodržováno pravidlo připojování 12 bp RSS k 23 bp RSS.
DNA kroužky, které vzniknou při přestavbě TCR, jsou nazývány TCR excision circles (TRECs) a jsou využívány jako markery T-buněk, které čerstvě opustily thymus.
Při přestavbě/sestřihu genových segmentů dochází k adici tzv. P- (vytvářejí palindromatickou sekvenci) a N- (non-template encoded) nukleotidů mezi úseky V, D a J. Za adici N-nukleotidů je odpovědná Terminální deoxynukleotidyl transferáza (TdT), která může na každém spoji přidat až 6 nukleotidů. Je odhadováno, že adicí P- a N-nukleotidů vzniká řádově 1013 možných kombinací spojení.
Při přestavbě β řetězce se uplatňuje mechanismus zvaný alelická exkluze. Po úspěšné přestavbě jedné z alel pro β řetězec a vzniku funkčního řetězce dojde k zastavení přestavby druhé alely pro tento řetězec. U α řetězce není alelická exkluze striktně dodržována a nejsou tedy výjimkou T-buňky exprimující na svém povrchu dva rozdílné αβ TCR. Avšak pouze jeden z těchto dvou TCR je schopen vázat vlastní MHC a je tudíž funkční.
Exprese TCR komplexu
Aby byl TCR exprimován na povrchu buňky, musí dojít k jeho správnému sbalení v ER. K tomu je potřeba asociace s dalšími molekulami. Jedná se molekuly tvořící tzv. CD3 komplex (řetězce CD3ε, CD3γ, CD3δ) a řetězce ζ, popřípadě η. ζ řetězce tvoří homodimer (90 %; popřípadě heterodimer s η řetězcem) spojený disulfidickými můstky. Předpokládá se, že TCRα řetězec interaguje s dimerem CD3δ:CD3ε a jedním řetězcem ζ, zatímco TCRβ interaguje s dimerem CD3γ:CD3ε. Tyto interakce jsou zprostředkovány odlišným nábojem aminokyselinových zbytků v transmembránových řetězcích (TCR domény jsou tvořeny bazickými aminokyselinami – lysin, arginin; zatímco CD3 domény obsahují především kyselé aminokyseliny – kyselina asparagová, glutamová). Asociace CD3 molekul s TCR v ER stabilizuje dimer αβ a umožní tak celému komplexu transport na povrch buňky. Seskupení TCR a CD3 komplexu se označuje jako TCR komplex
CD3 komplex
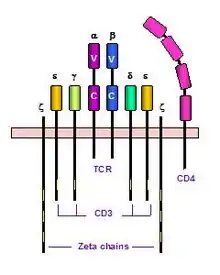
ζ a η řetězce jsou kódovány stejným genem, jedná se o dvě sestřihové varianty téže mRNA. ζ řetězec má krátkou extracelulární doménu, transmembránovou část a velkou cytoplazmatickou doménu. γ, δ a ε řetězec jsou členy imunoglobulinové super rodiny. Každý z těchto řetězců se skládá z Ig-like extrecelulární domény, transmembránové a cytoplazmatické domény. Transmembránové domény všech řetězců obsahují negativně nabité aminokyseliny (kyselina glutamová a asparagová).
Thymocytární vývoj a přestavba TCR
T-buňky z kostní dřeně přichází do thymu v tzv. double-negativním (DN) stádiu (T-buňky nemají na svém povrchu ani jeden z koreceptorů CD4/8). Tyto buňky ještě nezačaly s přestavbou genů pro TCR řetězce. Jsou popsány 4 stupně DN stádia, každé z nich je definováno ne/expresí určitých povrchových markerů. Přestavba genů pro TCR –γ, -δ a –β začíná prakticky ve stejném okamžiku, ve stádiu DN2. γδ TCR tedy vznikají mezi stádii DN2 a DN3 a stávají se téměř okamžitě zralými γδ T-buňkami (projdou ještě několika povrchovými změnami). Tento sled dějů vysvětluje, proč na většině γδ T-buněk není exprimován ani jeden z koreceptorů a proč je v těle více αβ TCR buněk – aby se buňka stala γδ pozitivní, musí úspěšně přestavět dva řetězce, zatímco k postupu směrem k pozitivitě na αβ TCR stačí v tomto stádiu přestavba pouze jednoho řetězce.
Ve stádiu DN3 buňky přestávají proliferovat. V cytoplazmě je možné nalézt proteinové produkty TCR β přestavby. Vzniklý β řetězec se pojí s 33kDa glykoproteinem zvaným pre-Tα. Tento dimer dále asociuje s proteiny CD3 komplexu a vzniká tak komplex zvaný pre-T-cell receptor (pre-TCR).
Imunoglobulinová doména na pre-Tα zajišťuje dva důležité kontakty – jedním z nich je interakce s Vβ řetězcem vlastního pre-TCR. Druhým je kontakt s Vβ doménou jiného pre-TCR. Exprese pre-TCR tedy vede k dimerizaci TCR nezávislé na ligandu, jejímž výsledkem je signalizace do buňky. Tento signál vede k proliferaci a zastavení další přestavby β řetězce. Buňka postupuje do stádia DN4, z něj pak do stádia double pozitivního (DP) – buňky exprimují oba dva typy koreceptorů – CD4 i CD8. V DP stádiu dochází k masivní proliferaci.
Přestavba α řetězce začíná v okamžiku, kdy DP thymocyty přestanou proliferovat a v buňce stoupne hladina RAG2 proteinu, který se účastní genové přestavby. Proliferační fáze před přestavbou α řetězce zvyšuje diverzitu T-buněk. DP thymocyt s kompletním TCR je oprávněn projít pozitivní a negativní selekcí.
- Výsledkem pozitivní selekce je tzv. MHC restrikce – TCR jsou selektovány na to, aby dokázaly vázat vlastní MHC molekuly. Přesto je v těle až 5 % alloreaktivních T-lymfocytů – tedy lymfocytů, které rozpoznají peptid na tělu cizím MHC glykoproteinu. Během pozitivní selekce jsou stále exprimovány RAG1/2 a TdT, buňky tedy stále mohou přestavovat α řetězec.
- Negativní selekcí neprojdou buňky, které na svém povrchu exprimují TCR s vysokou afinitou ke komplexu peptid:MHC.
Doposud není zcela jasné, jak je zajištěno, že signalizace přes TCR někdy vede k pozitivní, někdy k negativní selekci. Pravděpodobně hraje kritickou roli síla signálu.[1]
Vazba αβ TCR na antigen
TCR rozpoznává antigen v kontextu MHC molekul (Major histocompatibility complex). V závislosti na přítomném koreceptoru se jedná o MHC I. třídy (koreceptor CD8) či MHC II. třídy (koreceptor CD4). Navíc se pomocí těchto CD molekul rozlišují mezi T-buňkami odlišné populace. Molekula CD8 je exprimována na cytotoxických T-lymfocytech (CTL), zatímco molekula CD4 je spojována s populacemi pomocných T-lymfocytů (Th) a regulačních T-lymfocytů (Treg).
CD4
CD4 je jedno řetězcový transmembránový protein složený ze 4 Ig-like domén. Tento korececeptor se váže na MHC glykoproteiny II. třídy v místě styku α2 a β2 domény MHC. Tato oblast je vzdálena od místa vazby peptidu/TCR na MHC molekulu, tudíž se na jednu molekulu MHC mohou současně vázat TCR i CD4. Intracelulární část CD4 molekuly interaguje s proteinovou kinázu Lck. V případě kontaktu CD4 s komplexem TCR:peptid:MHC přivádí Lck blízko k intracelulárním signálním komponentám asociovaným s TCR. Vzhledem ke způsobu, jakým dochází ke generaci signálu do buňky po vazbě TCR na příslušný ligand, je takto zajištěno jeho zesílení. Pokud se na komplex peptid:MHC váže zároveň TCR i molekula CD4, stává se T-buňka přibližně 100× senzitivnější k antigenu než bez CD4 molekuly.
CD8
CD8 molekula je heterodimerní transmembránový protein složený ze dvou řetězců: α a β. Každý z řetězců obsahuje 1 Ig-like doménu, extracelulární části jsou silně glykosylovány a oba řetězce jsou spojeny disulfidickými můstky. Koreceptor CD8 se slabě váže k invariantní části α3 domény MHC molekuly I. třídy a k interakci dochází i mezi aminokyselinovými zbytky na bázi α2 domény. I v tomto případě je stejně jako u koreceptoru CD4 zajištěna vazba na jednu MHC molekulu jak TCR tak i CD8. Síla vazby CD8 na MHC I. třídy je ovlivněna mírou glykosylace CD8 molekuly – větší množství kyseliny sialové snižuje sílu interakce. Míra sialylace se v průběhu zrání T-buňky mění, mění se také při aktivaci T-buňky a není vyloučeno, že glykosylace hraje roli při modulaci rozpoznávání antigenu. Intracelulární část molekuly CD8 také asociuje s proteinovou kinázou Lck, uplatňujíc tak stejný mechanizmus jako CD4 molekuly.
Molekulární kontakt
TCR leží diagonálně přes peptid a vazebnou rýhu na MHC molekule. TCRα leží přes α2 doménu MHC I. molekuly (interakci zajišťuje především CDR2 smyčka) a N-konec peptidu (s peptidem interaguje smyčka CDR1). TCRβ leží přes α1 doménu MHC I. molekuly (CDR2) a C-konec peptidu (CDR1). CDR3 smyčky řetězců α i β jsou umístěny nad středem peptidu ležícího ve vazebné rýze MHC I molekuly. Kontakt s komplexy peptid:MHC II. třídy je zajištěn podobným způsobem – α řetězec interaguje s N-koncem peptidu, zatímco β řetězec s C-koncem peptidu. Podíl jednotlivých smyček na vazbě je však variabilní a není symetrický s ohledem na příslušné mateřské řetězce.
TCR interaguje rozdílně s MHC I. a II. třídy. S MHC II. třídy interaguje víc aminokyselinových reziduí. Tento poznatek je konsistentní s faktem, že TCR vykazuje vyšší afinitu interakce právě s MHC II. třídy.
Afinita TCR ke komplexu peptid:MHC je nízká až střední: Kd~ 10−4-10−7 M (ve srovnání s Ig: Kd~10−6-10−10 M se jedná o slabou interakci). Vazbu k buňce prezentující antigen (APC) zesilují adhezívní molekuly. Tím je prodloužena doba interakce mezi APC a T-buňkou a T-buňka získá dostatečný čas ke generaci signálu.
Buněčný kontakt
Zdá se, že na počátku kontaktu mezi příslušnými buňkami převládá interakce mezi TCR a MHC, T buněčný receptor je tak naváděn do správné polohy vůči peptidu navázanému na MHC. Poté dochází k sekundární, detailnější interakci s peptidem, stejně tak i s molekulou MHC. Je obtížné odhadnout, zda za celkovou vazebnou energií stojí především kontakt TCR s MHC či s peptidem. Jisté je, že pouze několik aminokyselin na rozhraní obou komplexů (peptid:MHC a TCR) uskutečňuje esenciální kontakt, který předurčuje specifitu a sílu vazby. Mutace jak v molekule MHC tak v peptidu mohou vést k odlišné imunitní odpovědi.
- Aby však došlo ke generaci signálu, je potřeba ještě dalších interakcí (B7:CD28). Díky těmto aditivním vazbám dochází ke stabilizaci celého komplexu (TCR/MHC) a k efektivní odpovědi T-buňky na antigen.
Signalizace přes TCR
Molekuly asociované s TCR obsahují v cytoplazmatické oblasti tzv. ITAMs (Immunoreceptor Tyrosin-based Activation Motifs). Po jednom motivu obsahují řetězce CD3γ, -δ a –ε. Řetězec ζ obsahuje 3 tyto motivy. Na jeden TCR komplex tedy připadne 10 motivů ITAM. V každém ITAM jsou dva tyrosinové zbytky, které mohou být fosforylovány. Základem signalizace přes T-buněčný receptor je fosforylace právě těchto tyrosinů, a to protein-tyrosinovými kinázami (PTK) z rodiny Src. Nejdůležitější je PTK Lck (p56Lck), dále pak PTK Fyn. Na fosforylované ζ řetězce se váže další PTK se dvěma SH2 doménami. Jedná se o ZAP-70. Tato PTK je po navázání na ζ řetězec také fosforylována Lck a tudíž aktivována. Aktivovaná ZAP-70 fosforyluje adaptorové proteiny (SLP-76, LAT), které umožňují následnou signalizaci několika směry:
- Dochází k aktivaci fosfolipázy Cγ ([PLCγ), která způsobí vznik diacylglycerolu (DAG) a inositol 1,4,5-trifosfátu (IP3) z fosfoinositol bisfosfátu(PIP2). DAG dále aktivuje signalizační dráhu protein kinázy C (PKC), signalizace přes IP3 vede k vylití Ca2+ z ER.
- Druhou signalizační větví je aktivace Ras. Tento malý G-protein dále spouští signalizační dráhu MAPK (mitogen-activated protein kinase).
V signalizaci hraje svou roli i membránově vázaná fosfatáza CD45, která má vliv na udržení aktivity PTK.
Aktivace T-buněk
T-buňky aktivované interakcí TCR komplexu s komplexem peptid:MHC vstupují do buněčného cyklu, začínají proliferovat a diferencovat se v efektorové či paměťové buňky. Signalizace přes TCR komplex vede k aktivaci tzv. immediate genů (exprese začíná do půl hodiny po aktivaci, jedná se např. o geny c-Fos, c-Myc, c-Jun, NFAT, NF-κB), časných genů (exprese začíná během 1 -2 hodin, mezi tyto geny patří např. IL-2, IL-2R, IL-3, IL-6, IFNγ) a pozdních genů (jedná se o geny kódující především adhezívní molekuly).
Imunologická synapse
Jedním z rysů iniciační fáze TCR signalizace je vznik supramolekulární struktury zvané imunologická synapse (někdy je též nazývána bull’s-eye-like, doughnut-like ring, SMAC – supramolecular activation complex). Jedná se o dynamickou, vysoce organizovanou strukturu, která vzniká v místě styku T-buňky a APC. Centrální oblast této struktury je bohatá na TCR/CD3 komplexy, zatímco vnější část je tvořena převážně adhezívními molekulami (např. LFA1). Vznik synapse není nutný pro iniciaci TCR-signalizace, ale T-buňky se tímto způsobem efektivněji aktivují.
Naivní T-buňky však ke své aktivaci potřebují ještě další, nespecifické kostimulační signály. Bez těchto signálů buňky přecházejí do stavu anergie.
Superantigeny
Superantigeny jsou rozpoznávány T-buňkami, aniž by tyto byly rozštěpeny na peptidy a navázány na MHC molekuly. Superantigen se váže na vnější povrch MHC II. třídy, které již váže nějaký peptid, a na Vβ oblast mnoha T-buněčných receptorů. Bakteriální superantigeny váží především CDR2 smyčku, méně často pak smyčku CDR1. Hlavní vazebné místo pro virové superantigeny je tzv. hypervariabilní smyčka 4 (HV4 loop) na TCRβ.
Každý superantigen je specifický pro jeden či několik různých Vβ genových produktů. Z tohoto důvodu může dojít k aktivaci 2–20 % všech T-buněk. Takto aktivované CD4+ T-buňky následně sekretují cytokiny. Vzhledem k obrovské produkci těchto látek dochází u postiženého jedince k systémové toxicitě a potlačení adaptivní odpovědi na patogen. Mezi bakteriální superantigeny se řadí Staphylococcální enterotoxiny (SES), které způsobují otravu jídlem, či toxin způsobující syndrom toxického šoku.
Superantigeny mohou být i endogenního původu, jedná se o membránové proteiny, které jsou kódovány určitými viry. Příkladem je myší savčí nádorový virus (MTV, Mammary tumor virus), který dává vzniknou proteinu MLS (Minor lymphocyte stimulating) determinants.
Mitogeny
Buňky mohou být nespecificky aktivovány k proliferaci látkami zvanými mitogeny. Mezi T-buněčné mitogeny patří Phytohemaglutinin (PHA) a Concanavalin A(ConA). Tyto molekuly se váží na povrchové molekuly (jako např. na TCR komplex), shlukují je na povrchu T-buněk a mimikují tak shlukování způsobené prezentací antigenu na APC. Výsledek takovéto stimulace však není odpovídající odpovědi buňky na specifický ligand (rozdíl je v produkci cytokinů apod.).
γδ TCR
Populace T-buněk v lidské krvi je z 90–95 % tvořena T-lymfocyty nesoucími αβ TCR. Jen minoritní populace nese TCR tvořené heterodimerem řetězců γδ. V epitelech (kůže, střevo, apod.) mohou však γδ T-buňky dominovat. γδ TCR jsou pravděpodobně specializovány k vazbě určitých typů ligandů (jedná se např. o určité HSP, tetanus toxoid, ligandy nepeptidové povahy, fosforylované ligandy či mykobakteriální lipidové antigeny). Zvláštností tohoto typu buněk jest schopnost vázat volné antigeny, tedy antigeny nenavázané na MHC molekulách. γδ TCR však mohou vázat antigeny i na MHC-like molekulách (jedná se o molekuly připomínající MHC I. třídy). γδ TCR jsou relativně nepolymorfní.[2]
O funkci γδ T-buněk toho není příliš známo; otázka jejich úlohy při imunitních odpovědích doposud nebyla uspokojivě zodpovězena, díky svému charakteru však bývají γδ T-buňky spojovány spíše s vrozenou než s adaptivní imunitou (mimo jiné pro jejich rychlou schopnost reakce na antigen). Většina γδ T-buněk neexprimuje ani CD4 ani CD8 koreceptor, navíc většina γδ T-buněk exprimuje stejný γδ TCR. U lidí má tato majoritní populace γδ T-buněk na svém povrchu TCR (Vγ9/Vδ2)[3] rozpoznávající mikrobiální fosfolipid 3-formyl-1-butyl pyrofosfát (Mycobacterium tuberculosis a další bakterie a parazité). U myši i jiných druhů je majoritní populace γδ T-buněk namířena proti jiným, druhově specifickým patogenům. γδ T-buňky působí obdobným mechanismem jako CTL (využívají granulysin, perforin apod.). Během embryonálního vývoje dochází nejdříve k vývoji γδ T-buněk.
B-buněčný receptor/Imunoglobuliny a TCR.
Mezi oběma typy imunoreceptorů lze najít několik homologií – na úrovni strukturální (V a C oblasti na TCR odpovídají V a C doménám na imunoglobulinech), na úrovni signalizační (oba dva typy receptorů jsou asociovány s proteinovými kinázami a nejsou kinázami sami o sobě), či na úrovni genomové (uspořádání genových segmentů pro α a β řetězce je homologní s uspořádáním genových segmentů pro lehký a těžký řetězec BCR/Ig). Navíc přestavba genů pro oba typy imunoreceptorů využívá stejných enzymů (např. RAG1/2, TdT).
Imunoglobuliny se však na rozdíl od TCR vyskytují i v sekretované (solubilní) podobě, zatímco TCR jsou pouze membránově vázané. B-buňky, jejich imunoreceptory, procházejí somatickými mutacemi, afinitním zráním v průběhu imunitní reakce, avšak T-buňky po opuštění thymu již své imunoreceptory nepřestavují. Dochází pouze ke klonální expanzi specifických T-buněk. Další rozdíl tkví v počtu vazebných míst pro antigen – TCR má pouze jedno vazebné místo, zatímco BCR/Ig mají dvě.
NKT buňky
Dalším typem buněk, které nesou TCR jsou NKT buňky. Tyto buňky mají charakter CTL i NK buněk a bývají řazeny do vrozené imunity. TCR na lidských NKT buňkách je invariantní, s α i β řetězci, které jsou kódovány specifickými genovými segmenty (Vα24-Jα18; Vβ11). TCR na NKT buňkách nerozeznávají komplexy peptid:MHC ale glykolipidy prezentované na nepolymorfních CD1d molekulách.
Současný výzkum na poli TCR
V současné době se výzkum zaměřuje na několik odvětví. Jedním z nich je výzkum γδ T-buněk, jejich TCR a úlohy v imunitním systému. Další velkou skupinu tvoří výzkum zaměřený na proces spouštění TCR signalizace. Neposledním polem zájmu je příprava transgenních TCR[4] a jejich využití v klinické praxi[5] – zejména při léčbě leukémií a jiných neoplázií.
Reference
- Moran AE, Hogquist KA. T-cell receptor affinity in thymic development. Immunology. 2011, s. 261-267 PMID:22182461. (anglicky)
- Born WK, Zhang L, Nakayama M, Jin N, Chain JL, Huang Y, Aydintug MK, O'brien RL. Peptide antigens for gamma/delta T cells. Cellular and Molecular Life Sciences. 2011, s. 2335-2343 PMID:21553233. (anglicky)
- Riganti Ch, Massaia M, Davey MS, Eberl M. Human γδ T-cell responses in infection and immunotherapy: Common mechanisms, common mediators?. European Journal of Immunology. 2012, s. 1668-1676 PMID:22806069. (anglicky)
- Bettini ML, Bettini M, Vignali DAA. T-cell receptor retrogenic mice: a rapid, flexible alternative to T-cell receptor transgenic mice. Immunology. 2012, s. 265-272 PMID:22348644. (anglicky)
- Uttenthal BJ, Chua I, Morris EC, Stauss HJ. Challenges in T cell receptor gene therapy. The Journal of Gene Medicine. 2012, s. 386-399 PMID:22610778. (anglicky)
Literatura
- Murphy, Travers, Walport, Janeway (2011) – Janeway‘s Imunnobiology, 8th edition, Garland Science
- Alberts, Johnson, Lewis, Raff, Roberts, Walter (2008): Molecular Biology of the Cell, 5th edition, Garland Science
- Alberts, Johnson, Lewis Raff, Roberts, Walter (2002): Molecular Biology of the Cell, 4th edition, Garland Science, ISBN 0-8153-4072-9
- Male, Brostoff, Roth, Roitt (2006) – Immunology, 7th edition, Elsevier Health Science,
- Kindt, Goldsby, Osborne (2007) – Kuby Immunology, 6th edition, W. H. Freeman and Company
Externí odkazy
 Obrázky, zvuky či videa k tématu T-buněčný receptor na Wikimedia Commons
Obrázky, zvuky či videa k tématu T-buněčný receptor na Wikimedia Commons