Ardipithecus ramidus
Ardipithecus ramidus je druh vyhynulého hominida, žijící ve starším pliocénu (před 4,4 miliony let) na území dnešní Etiopie ve východní Africe. Je jedním z prvních zástupců lidské vývojové linie po oddělení od předků šimpanzů. Starší jsou pouze rody Sahelanthropus a Orrorin a druh Ardipithecus kadabba, jejichž postavení ale zatím není zcela jisté. Předpokládá se, že Ardipithecus ramidus mohl být předchůdcem mladšího rodu Australopithecus.
Stratigrafický výskyt: starší pliocén | |
|---|---|
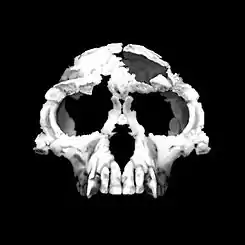 Rekonstrukce lebky druhu Ardipithecus ramidus' | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Třída | savci (Mammalia) |
| Řád | primáti (Primates) |
| Čeleď | Hominidé (Hominidae) |
| Podčeleď | Homininae |
| Tribus | Hominini |
| Rod | Ardipithecus White et al., 1995 |
| Binomické jméno | |
| Ardipithecus ramidus White et al., 1995 | |
| Některá data mohou pocházet z datové položky. | |
Rodové jméno Ardipithecus je složeninou afarského slova Ardi („zem“, „podlaha“) a řeckého πίθηκος - pithekos („opice“). Druhový přívlastek ramidus znamená v lokálním jazyce etnika Afar „kořen“. Společně lze tedy označení druhu Ardipithecus ramidus volně přeložit jako „kořenová zemní opice“, což odkazuje k jeho postavení v počátcích lidského rodu.[1]
Objevy
První ostatky druhu Ardipithecus ramidus zachytil japonský paleoantropolog Gen Suwa 17. prosince 1992 na lokalitě Aramis na středním toku řeky Awaš v Afarské pánvi (severovýchodní Etiopie).[2]

Americko-etiopský výzkumný tým, který vedl paleoantropolog Tim D. White, následně v letech 1992–1993 objevil na téže lokalitě dalších 16 zlomků kostí. Nálezy, zahrnující především jednotlivé zuby, ale i zlomky lebky a končetin, byly popsány v roce 1994 jako druh Australopithecus ramidus.[3] Již v roce 1995 však bylo vydáno doplnění původní publikace, ve kterém byl navržen zcela nový rod Ardipithecus s druhem Ardipithecus ramidus.[4] Zároveň byl také oznámen nález unikátně dochované kostry, na jejíž publikaci se poté čekalo až do roku 2009.[5] Výzkumy v Aramis a na přilehlých lokalitách Kuseralee Dora a Sagantole ale pokračovaly i nadále, takže v roce 2009 bylo známo celkem 115 nálezů, představujících ostatky nejméně 36 jedinců, včetně zmíněné částečně kompletní kostry.[6]
V letech 1997–2001 byly na pěti nových lokalitách v oblasti středního toku řeky Awaš (též Middle Awash) zachyceny kosterní ostatky, určené jako poddruh Ardipihecus ramidus kadabba.[7] Ty ale byly v roce 2004 přeřazeny do samostatného druhu Ardipithecus kadabba.[8]
V letech 1999–2003 bylo nalezeno celkem 11 zlomků kostí také na lokalitě As Duma v oblasti Gona, severně od Middle Awash. Jsou značně nekompletní a jejich příslušnost k druhu Ardipithecus ramidus se předpokládá zejména na základě podobného stáří.[5] Ostatky, představované především jednotlivými zuby a čelistmi, ale také články prstů, patřily nejméně devíti jedincům.[9]
Ardi
V roce 2009 byl po 15 letech od objevu publikován dlouho očekávaný popis unikátně dochované kostry druhu Ardipithecus ramidus.[10] Zachyceno bylo 45% všech kostí, takže je možné učinit si poměrně přesnou představu o velikosti, vzhledu i způsobu pohybu tohoto jedince. Jednalo se o ostatky samice, která byla podle rodového názvu pojmenována Ardi.
Nálezu bylo v prvním říjnovém čísle prestižního časopisu Science věnováno celkem 11 článků.[11] Publikace způsobila světovou senzaci, kterou zmínilo velké množství médií. Nález byl přitom označován jako „objev století“. Ardi se tak stala slavnější než kostra Lucy (Australopithecus afarensis), kterou vědci našli v roce 1974 v Hadaru, pouhých 75 km severně od Aramis.[12]
Ardi (katalogové číslo ARA-VP-6/500) nalezl etiopský paleoantropolog Yohannes Hailes-Selassie, který 5. listopadu 1994 zachytil nejprve dva zlomky záprstních kostí.[13] Následoval odkryv pánve, horních a dolních končetin, lebky i dolní čelisti se zuby - celkem asi 125 zlomků. Nedochovala se pouze páteř, obratle a pažní kosti. Kostra Ardi však nebyla odhalena v původní poloze. Jednotlivé části byly díky erozi roztroušeny na větší ploše v rozsahu zhruba 4 m².[6] Všechny byly navíc ve velmi špatném stavu a takřka před očima se rozpadaly. Větší kosti i lebka byly rozdrcené na drobné zlomky. Proto byly při výzkumu vyzvedávány s celými bloky okolní zeminy a pečlivě baleny, aby mohly být vyjímány a preparovány až v přijatelnějších podmínkách laboratoří Národního muzea Etiopie v Addis Abeba.[13]
Kostra byla na lokalitě Aramis odkrývána celé tři sezóny, než bylo detailně propátráno celé okolí nálezu. Veškerá zemina byla prosívána na jemných sítech, aby byly odhaleny i nejmenší kousky kostí. Kromě samotných ostatků Ardi tak bylo zachyceno na 150 000 vzorků fosilních rostlin a živočichů, které byly využity k rekonstrukci životního prostředí na počátku pliocénu.[13]
Po dokončení prací přímo na lokalitě následovala zdlouhavá preparace kostí, vytváření pracovních odlitků a jejich sestavování v laboratořích. Zároveň probíhaly virtuální rekonstrukce rozdrcené pánve a lebky. Tyto části kostry byly nasnímány a následně sestaveny do původní podoby pomocí počítačové tomografie. Jednotlivé zlomky byly virtuálně zbaveny zbytků sedimentů a postupně podle průběhu prasklin usazovány na původní místo.[13]
Místo nálezu je v současnosti vyznačeno hromadou kamení, navršenou v roce 1997 podle místního afarského zvyku.[6]
Datování
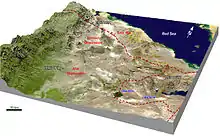
Ostatky druhu Ardipithecus ramidus z oblasti Middle Awash byly nalezeny mezi dvěma vrstvami sopečného tufu. Obě tyto vrstvy mají podle radiometrického datování téměř shodné stáří (4,419 a 4,416 milionů let). To podle zákonů stratigrafie znamená, že také kosti ardipitéka jsou staré 4,4 miliony let.[14] Sedimenty, ze kterých nálezy pocházejí, byly uloženy během pouhých 100 - 10 000 let, pokud se vezme v úvahu možná odchylka výsledků měření. To znamená, že také údaje o životním prostředí, získané analýzou fosilních zbytků rostlin i živočichů jsou velmi přesné. Zahrnují totiž pouze organismy, žijící na místě během relativně krátkého časového úseku, kdy se přírodní podmínky mohly změnit jen minimálně.[14]
Nálezy z lokality As Duma jsou zřejmě o něco starší, asi z doby před 4,51 - 4,32 miliony let. Datování ostatků z této lokality je ovšem problematické.[9]
Popis
Ardipithecus ramidus je nejlépe doloženým a poznaným zástupcem nejstarších homininů. Oproti ostatním druhům (Sahelanthropus tchadensis, Orrorin tugenensis, Ardipithecus kadabba) nebyly odhaleny pouze drobné zlomky kostí mnoha jedinců z různých lokalit a časových období, ale také poměrně kompletní kostra jediného tvora. Většina poznatků o vzhledu a způsobu života druhu Ardipithecus ramidus tak vychází právě z unikátního nálezu samice Ardi.
Ardi dosáhla výšky asi 120 cm a hmotnosti kolem 50 kg.[15] Byla tak zhruba stejně velká jako samice moderního šimpanze a měla i obdobně velkou hlavu a mozek.[13] Na základě nálezů mnoha dalších jedinců podobné velikosti se nepředpokládá výrazný pohlavní dimorfismus, takže samci mohli být zhruba stejně velcí.[6]

- malý objem - lebeční kapacita dosahovala u samic jen asi 300 - 350 cm³, což přibližně odpovídá velikosti mozku u samic šimpanzů
- nápadně pneumatizovaná šupina spánkové kosti (squama occipitalis) podobně jako u šimpanzů
- velmi plochý čelistní kloub, tvarově odpovídající šimpanzům
- chybějící žlábek za nadočnicovým valem, který je typický pro gorily a šimpanze
- málo vystupující čelisti v porovnání s lidoopy
- málo vystupující lícní kosti v porovnání s australopitéky
- poměrně velké očnice
- malé hrbolky pro skloubení s krční páteří (condylus occipitalis)
- krátká spodina lebeční v porovnání s lidoopy
- vpřed posunutý velký týlní otvor (foramen magnum) v porovnání s lidoopy
- tupý bradavkový výběžek (processus mastoideus) v porovnání s výrazným pyramidálním výčnělkem šimpanzů
- výrazně hlubší žlábek incisura mastoidea pro úpon dvojbříškového svalu v porovnání se šimpanzi
- poměr velikosti předních a zadních zubů na pomezí mezi australopitéky a dnešními lidoopy - stoličky jsou větší než u současných šimpanzů, ale menší než u rodu Australopithecus; naopak řezáky jsou oproti šimpanzům výrazně menší a užší, ale v porovnání s australopitéky a lidmi ještě velké
- poměrně tenká zubní sklovina - mírně silnější než u moderních šimpanzů, ale výrazně slabší než u rodu Australopithecus
- špičáky diamantového průřezu, které jen minimálně vystupují z řady zubů, na rozdíl od výrazných špičáků moderních i vyhynulých lidoopů - není velký rozdíl ve velikosti špičáků samců a samic
- nápadně primitivní třenové zuby (premoláry), tvarově bližší spíše lidoopům než homininům - korunka má zřetelně asymetrický tvar, ale některými detaily se již blíží třenovým zubům australopitéků
- stoličky s nízkými zaoblenými hrbolky, tvarově podobné mladším homininům - jsou již zvětšené, ale nedosahují ještě extrémních rozměrů jako u rodu Australopithecus; postrádají specializace moderních lidoopů (nápadné zvrásnění povrchu šimpanzů či výrazné hrbolky goril)
- mléčná první stolička m1 morfologicky nerozeznatelná od šimpanzů - je malá, úzká a postrádá odvozené znaky homininů
- velmi malá diastema mezi řezáky a špičáky horní čelisti, v porovnání s lidoopy
- chybějící ostřící komplex[17] mezi špičáky a třenovými zuby, který se vyskytuje u lidoopů, ale vymizel u lidí a australopitéků - špičáky i třenové zuby mají ještě poměrně primitivní tvar, ale stopy vzájemného ostření již postrádají
- stopy opotřebení podobné mladším homininům, odlišné od lidoopů
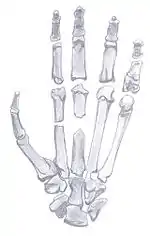
- značná délka v poměru k dolním končetinám v porovnání s člověkem
- celkově primitivní stavba bez patrných adaptací k brachiaci nebo kotníkochodectví (chůzi s oporou o kotníky prstů předních končetin, která je charakteristická pro gorily a šimpanze)
- konzervativní, nespecializovaná morfologie pažní i loketní kosti a ruky, odpovídající mladším homininům
- plně natažitelný loketní kloub, který však postrádá jakékoliv znaky běžně užívaného pohybu v závěsu na pažích pod větvemi
- značně pohyblivé zápěstí i záprstí oproti tuhým kloubům se silnými vazy u šimpanzů a goril, jimž přední končetina slouží jako významná opora při chůzi
- poměrně krátké záprstní kosti, nepřizpůsobené kotníkochodectví
- dlouhé a zakřivené články prstů, schopné pevného úchopu, podobně jako prsty australopitéků
- poměrně dlouhý, robustní a dobře osvalený palec - delší než u lidoopů, ale ještě krátký proti lidem
- dobře patrný úpon plně funkčního dlouhého ohýbače palce (musculus flexor pollicis longus) - u afrických lidoopů tento sval zcela chybí nebo je redukovaný
Znaky pánve:[6]
- sedací kost tvarem podobná gorilám a šimpanzům, ale již s patrným sedacím zářezem (incisura ischiadica) homininů
- nízké a široké lopaty kyčelní kosti, stavbou podobné australopitékům a lidem
- dobře patrný přední dolní trn kyčelní (spina iliaca anterior inferior), který u lidoopů chybí
- relativně krátká plocha pro sponu stydkou (facies symphysialis)
Znaky chodidla:[6]
- poměrně primitivní stavba, bez adaptací moderních lidoopů
- protistojný palec, na rozdíl od ostatních homininů, kteří již mají palec na noze přitažený k ostatním prstům
- přítomnost drobné kůstky os peroneum ve šlaše dlouhého svalu lýtkového, která je součástí hlezna lidí i opic Starého světa, ale vymizela u šimpanzů a goril
- dlouhé zánártí v porovnání s africkými lidoopy
- nártní kosti s rozměrnými diafýzami
- dlouhé, prohnuté a poměrně robustní články prstů v porovnání s lidmi
- méně pohyblivý kotník v porovnání se šimpanzi
Způsob života
.JPG.webp)

Podle obrovského množství vzorků dalších zvířat i rostlin, zachycených při výzkumu lokality Aramis žil Ardipithecus ramidus v lužním lese, tvořeném břestovcem, fíkovníky, voskovníky a palmami.[19] Nejednalo se o hustý tropický deštný les, ale spíše o otevřenější prosvětlený porost, kde sluneční svit mohl pronikat korunami stromů, takže zemi pokrývaly traviny či nízké keře.[14] Mezi nepočetnější obyvatele patřily úzkonosé opice a antilopy rodu Tragelaphus, zatímco ardipiték samotný se vyskytoval poměrně vzácně.[19]
Stromy se však ve větším množství zřejmě nacházely pouze v úzkém pásu podél řeky nebo jezerního břehu, kde byl dostatek vláhy pro hustší vegetaci s bohatou faunou, zatímco dál od vody se rozkládala sušší, otevřenější krajina savany, jen s občasným porostem.[20] Té se ovšem ardipiték spíše vyhýbal, protože v korunách stromů hledal potravu i úkryt před dravci.[19]
Celková stavba chrupu bez nápadných specializací ukazuje, že byl nevyhraněným všežravcem, který mohl převažující rostlinnou stravu doplňovat hmyzem, vejci i drobnými savci.[13] Na nižší podíl ovoce ve stravě ve srovnání s šimpanzi či orangutany ukazují menší řezáky, kterými dnešní lidoopi odstraňují vnější obaly plodů. Malé stoličky bez výrazně silné skloviny stejně jako mikroskopické stopy opotřebení na dochovaných zubech zase vylučují převažující konzumaci výrazně abrasivní, vláknité a tuhé stravy (např. ořechů či hlíz), kterou mohli alespoň částečně konzumovat australopitékové.[16]
Nápadná redukce špičáků může svědčit o menší míře soupeření mezi samci, zmírnění bojů o samice či teritorium a možná i o větší spolupráci samců a samic v péči o potomstvo.[16] To je odvozováno z faktu, že samci dnešních lidoopů používají výrazné špičáky zejména k vyjádření agrese a dominance v bojích mezi jedinci i skupinami. Stejně tak se ale špičáky mohly zmenšovat v důsledku změny stravování, zejména se zvyšující se potřebou konzumace tužší potravy. Při žvýkání a drcení tužších rostlinných částí v dobách nedostatku měkkých plodů mohly dlouhé vyčnívající zuby překážet.[21] Přesný důvod redukce špičáků u časných homininů však zatím není známý a mohlo se jednat i o kombinaci více faktorů.

Bipedie
Většina odborníků v současnosti nepochybuje, že jedinci druhu Ardipithecus ramidus používali určitou formu bipedie (vzpřímené chůze po dvou končetinách), protože pro tuto schopnost svědčí hned několik znaků.
Poměrně vpředu umístěný velký týlní otvor spolu se zkrácenou spodinou lebeční naznačují, že lebka seděla nahoře na páteři jako u lidí, nikoliv před ní jako u kvadrupedních lidoopů.[2] Bipedie jedinců z oblasti Gona je odvozována z článku prstu levé nohy, na němž je kloubní ploška pro spojení s nártem orientovaná stejně jako u australopitéků. Shodně je utvářen také článek prstu druhu Ardipithecus kadabba. Podle tohoto znaku se předpokládá, že Ardipithecus se při chůzi odrážel od špičky nohy, na rozdíl od šimpanzů, kteří se odrážejí z vnější hrany chodidla.[1][22]
Hlavní oporou pro předpokládanou schopnost vzpřímené chůze je však pánev. Po náročné rekonstrukci je patrné, že se její tvar značně liší od pánve šimpanzů, ačkoliv do pánve vzpřímeně chodících australopitéků nebo lidí má ještě daleko.[23] Morfologie svalových úponů a zejména dobře patrný přední dolní trn kyčelní (spina iliaca anterior inferior) ovšem ukazují, že hýžďové svalstvo již bylo přestavěno tak, aby ardipiték mohl chodit po dvou a nekýval se přitom ze strany na stranu jako moderní lidoopi.[23] Stehenní kost a pánev byly spojeny pevnými vazy, které zajišťovaly dostatečnou stabilitu při chůzi a přímý sval stehenní (rectus femoris musculus) fungoval stejně jako u lidí, takže ardpiték mohl plně natáhnout koleno.[24] Malá výška pánve, které bylo docíleno díky nízkým a širokým lopatám kyčelní kosti, snížila celkové těžiště těla, a tak byl tento tvor schopen během chůze udržet rovnováhu na jediné noze. Utváření horní části pánve také naznačuje, že páteř již mohla být v bedrech prohnutá na způsob lidské lordózy, protože široce rozložené lopaty kyčelní kosti netísnily bederní obratle jako u dnešních šimpanzů.[23]
Významná je také přítomnost drobné kůstky os peroneum, která ukazuje, že chodidlo ardipitéka bylo oproti šimpanzům a gorilám poměrně tuhé. Poskytovalo tak dostatečně silný odraz od země při každém kroku.[25] Tato kůstka byla navíc u ardipitéka poměrně masivní a velká, takže dlouhý sval lýtkový mohl zároveň dobře zabezpečit přitažení palce i protažení nohy do špičky.[26][23]
Přestože na zemi se ardipiték mohl pohybovat pomocí dvojnohé chůze, patrně trávil ještě velkou část života v korunách stromů. Zde ovšem nevyužíval brachiaci ani vertikální šplhání jako gorily či šimpanzi. Spíše se po větvích pohyboval s využitím všech čtyř končetin a vzhledem ke značné velikosti těla musel být tento pohyb pomalejší a méně obratný než u výrazně lehčích opic. K tomu se ardipiték na předních končetinách neopíral o kotníky prstů jako moderní afričtí lidoopi, ale o plosky dlaní stejně jako opice i mnozí vyhynulí hominidé.[13] Kvadrupednímu ploskochodectví nasvědčuje množství znaků - poměr délky předních a zadních končetin, konzervativní, nespecializovaná stavba paže a ruky, pohyblivé zápěstí i záprstí.[18] Právě pohyblivost umožnila ardipitékovi dobře roznášet váhu těla na plosky dlaně a udržovat dobrý kontakt s povrchem, po kterém chodil. Naproti tomu ruka lidoopů musela být vyztužena silnými vazy, aby odolala značné váze těla při kotníkochodectví i při pohybu v závěsu na pažích.[18]
Častý pobyt ardipitéka v korunách stromů naznačuje i pánev. Její horní část byla přizpůsobená dvojnohé chůzi, ale spodní partie se zcela podobaly dnešním lidoopům, protože musely poskytnout potřebnou oporu pro silně osvalená stehna a kyčle, potřebné k aktivnímu lezení po větvích. Bipedie rodu Ardipithecus tudíž zjevně nebyla plně vyvinutá.[13] Na nízkou efektivitu chůze ukazuje také chodidlo s palcem v opozici. Ten sice spolu s dlouhými zakřivenými prsty pomáhal bezpečnému pohybu v korunách stromů, ale zároveň se kvůli němu nemohla vytvořit typicky lidská klenba nártu, tlumící nárazy při vzpřímené chůzi. Ani kolena nesměřovala k sobě tolik jako u australopitéků a lidí. Ardipiték tak nemohl chodit dobře, rychle ani daleko.[26]
Kultura
Nepředpokládá se, že by Ardipithecus ramidus vytvářel nějakou specifickou kulturu a vyráběl nástroje jako mladší homininé. Nelze však vyloučit, že využíval neupravené valouny nebo větve a klacíky podobně jako dnešní gorily, orangutani a šimpanzi.[27]
Fylogeneze

Většina odborníků v současnosti souhlasí s tím, že Ardpithecus ramidus patří do taxonu Hominina, i když stál ještě velmi blízko poslednímu společnému předku lidí a šimpanzů.[28] Zásadním argumentem pro toto zařazení je schopnost vzpřímené dvojnohé chůze, důležitými znaky jsou ale i vpřed posunutý velký týlní otvor a nevýrazné špičáky.[29] Všechny tyto detaily také posilují domněnky, že i starší rody Sahelanthropus, Orrorin a druh Ardipithecus kadabba patří mezi zástupce lidské vývojové větve. Byl dokonce vysloven názor, že rody Sahelanthropus, Orrorin a Ardipithecus mohly patřit jedinému tvoru, který se průběžně vyvíjel. K tomu jsou ovšem mnozí badatelé spíše skeptičtí.[13]
Problematická je u ardipitéka pouze slabá zubní sklovina, protože všechny mladší druhy homininů včetně člověka, ale také mnozí miocénní hominidé mají sklovinu relativně silnou.[30] Je však možné, že síla skloviny se relativně rychle měnila v důsledku potravní adaptace a nemusí být tudíž příliš spolehlivým znakem.[31]
Někteří badatelé také pochybují o způsobu, efektivitě a četnosti využití bipedie. Nedůvěra či rezervovanost ohledně vzpřímené chůze ardipitéka částečně vychází z velmi špatného stavu nalezených ostatků. Podle odpůrců mohlo během virtuální rekonstrukce původního tvaru pánve dojít k mnoha chybám, takže některé důležité úhly a vzdálenosti nemusí odpovídat realitě.[13][32]
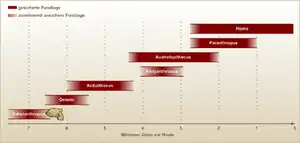
Objevitelský tým předpokládá, že Ardipithecus ramidus byl přímým nástupcem druhu Ardipithecus kadabba a předkem australopitéků.[13] Je možné, že jeho potomkem byl druh Australopithecus anamensis, který obýval týž prostor středního toku řeky Awaš jen o 200 tisíc let později. To by ovšem znamenalo, že k velkým změnám v anatomii muselo dojít během velmi krátkého časového úseku. Ardipiték ale mohl být i slepou vývojovou větví. V každém případě velmi dobře ukazuje, jak takový předek australopitéků mohl vypadat.[24]
Nález Ardi také výrazně změnil představy o podobě posledního společného předka šimpanzů a lidí. V současnosti tak převládá názor, že šimpanzi se vyvíjeli stejně intenzivně jako lidé a od společného předka se liší zhruba stejnou měrou. Tento předek se vůbec nepodobal dnešním lidoopům, nepoužíval ve velké míře brachiaci, vertikální šplhání ani kotníkochodectví a blížil se tak mnoha starším druhům miocénních hominidů.[33] Lidé přitom ve své kostře uchovali v mnoha ohledech spíše konzervativní znaky úzkonosých opic, zatímco šimpanzi, ale i gorily se nezávisle na sobě specializovali.[18]
Zajímavý je také fakt, že ardipiték byl patrně schopný chodit po dvou, přitom ale dával přednost zalesněným prostorám před otevřenou krajinou. To znamená, že bipedie má svůj původ v lesích, což přímo odporuje starším teoriím o vzniku bipedie v otevřené savaně v důsledku ústupu lesních porostů.[10]
Reference
- GIBBONS, A. První lidé: závody v hledání nejstaršího předka. Praha: Academia, 2011. 307 s.
- CARTMILL, M.; SMITH, F. H. The Human Lineage. Oxford: Wiley-Blackwell, 2011.
- WHITE, T. D.; SUWA, G.; ASFAW, B. Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia. Nature. 1994, roč. 371, s. 306–312.
- WHITE, T. D.; SUWA, G.; ASFAW, B. Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia. Nature. 1995, roč. 375, s. 88.
- SAWYER, G. J.; DEAK, V. The Last Human: A Guide to Twenty-two Species of Extinct Humans. [s.l.]: Yale University Press, 2007. 256 s.
- WHITE, T. D., ASFAW, B., BEYENE, Y., HAILE-SELASSIE, Y., LOVEJOY, C. O., SUWA, G., WOLDEGABRIEL, G. Ardipithecus ramidus and the Paleobiology of Early Hominids. Science. 2009, roč. 326, s. 64, 75–86.
- STRINGER, C.; ANDREWS, P. The Complete World of Human Evolution. Londýn: Thames and Hudson Ltd, 2005. 240 s.
- HAILE-SELASSIE, Y.; SUWA, G.; WHITE, T. D. Late Miocene Teeth from Middle Awash, Ethiopia, and Early Hominid Dental Evolution. Science. 2004, roč. 303, s. 1503–1505.
- SEMAW, S, SIMPSON, S. W., QUADE, J., RENNE, P. R., BUTLER, R. F., MCINTOSH, W. C., LEVIN, N., DOMINGUEZ-RODRIGO, M., ROGERS, M. J. Early Pliocene hominids from Gona, Ethiopia. Nature. 2005, roč. 433, s. 301–305.
- GIBBONS, A. Breakthrough of the year: Ardipithecus ramidus. Science. 2009, roč. 326, s. 1598–1599.
- Číslo časopisu Science věnované objevu Ardi
- EDGAR, B. Digging up the family bones. BioScience. 1995, roč. 45, s. 659–662. Dostupné online.
- GIBBONS, A. A New Kind of Ancestor: Ardipithecus Unveiled. Science. 2009, roč. 326, s. 36–40.
- WOLDEGABRIEL, G., AMBROSE, S. H., BARBONI, D., BONNEFILLE, R., BREMOND, L., CURRIE, B. et al. The geological, isotopic, botanical, invertebrate and lower vertebrate surroundings of Ardipithecus ramidus. Science. 2009, roč. 326, s. 65e1–65e5.
- Podobné váhové a výškové odhady byly učiněny i v případě rodů Orrorin a Sahelanthropus.
- SUWA, G., KONO, R. T., SIMPSON, S. W., ASFAW, B., LOVEJOY, C. O., WHITE, T. D. Paleobiological implications of the Ardipithecus ramidus dentition. Science. 2009, roč. 326, s. 69, 94–99.
- Špičáky jsou ostřeny o třenové zuby ve spodní čelisti.
- LOVEJOY, C. O., SIMPSON, S. W., WHITE, T. D., ASFAW, B., SUWA, G. Careful climbing in the Miocene: The forelimbs of Ardipithecus ramidus and humans are primitive. Science. 2009, roč. 326, s. 70, 70e1–70e8.
- WHITE, T. D., AMBROSE, S. H., SUWA, G., SU, D. F., DEGUSTA, D., BERNOR, R. L. et al. Macrovertebrate paleontology and the Pliocene habitat of Ardipithecus ramidus. Science. 2009, roč. 326, s. 67, 87–93.
- GANI, M. R.; GANI, N. D. River-margin habitat of Ardipithecus ramidus at Aramis, Ethiopia 4.4 million years ago. Nature communications. 2011, roč. 2, s. 602.
- HYLANDER, W. L. Functional links between canine height and jaw gape in catarrhines with special reference to early hominins. American journal of physical anthropology. 2013, roč. 150, s. 247–259.
- HARCOURT-SMITH, W. E. The Origins of Bipedal Locomotion. In: HENKE, W. C.; TATTERSALL, I. Handbook of palaeoanthropology 3. Berlín: Springer, 2007. S. 1483–1518.
- LOVEJOY, C. O., SUWA, G., SPURLOCK, L., ASFAW, B., WHITE, T. D. The pelvis and femur of Ardipithecus ramidus: the emergence of upright walking. Science. 2009, roč. 326, s. 71, 71e1–71e6.
- SIMPSON, S. W. Before Australopithecus: The Earliest hominins. In: BEGUN, D. R. A Companion to Paleoantropology. [s.l.]: Wiley-Blackwell, 2012. S. 417–433.
- Lidoopi, u nichž tato kůstka vymizela, se díky pružnějšímu chodidlu obratně pohybují ve větvích i při zachování poměrně značné tělesné váhy.
- LOVEJOY, C. O., LATIMER, B., SUWA, G., ASFAW, B., WHITE, T. D. Combining prehension and propulsion: the foot of Ardipithecus ramidus. Science. 2009, roč. 326, s. 72, 72e1–72e8.
- Ardipithecus ramidus na webu Australian Museum
- BEGUN, D. R. The earliest hominins - is less more?. Science. 2004, roč. 303, s. 1478–1480.
- BEGUN, D. R. Miocene hominids and the origins of the African apes and humans. Annual Review of Anthropology. 2010, roč. 39, s. 67–84.
- FISCHMAN, J. Putting our oldest ancestors in their proper place. Science. 1994, roč. 265, s. 2011–2012.
- STRINGER, C.; ANDREWS, P. The Complete World of Human Evolution. Londýn: Thames and Hudson, 2005. 240 s.
- Diskuse o pánvi rodu Ardipithecus (John Hawks weblog)
- LOVEJOY, C. O.; MCCOLLUM, M. A. Spinopelvic pathways to bipedality: Why no hominids ever relied on a bent-hip-bent-knee gait. Philosophical transactions of the Royal Society. 2010, roč. 365, s. 3289–3299.
Literatura
- GIBBONSOVÁ, A. První lidé: závody v hledání nejstaršího předka. Praha: Academia, 2011. 307 s.
- ROBERTSOVÁ, A. Evoluce: Příběh člověka. Praha: Knižní klub, 2012. 256 s.
- SVOBODA, J. Čas lovců: Aktualizované dějiny paleolitu. Brno: Akademické nakladatelství CERM, 2009. 298 s.