Mystacina novozélandská
Mystacina novozélandská, též zemolez novozélandský či netopýr novozélandský[3] (Mystacina tuberculata, maorsky pekapeka-tou-poto či jen pekapeka), je netopýr z rodu mystacina, který je endemitem Nového Zélandu. Spolu s mystacinou větší, která je však patrně vyhynulá, se jedná se o jediné zástupce hrabavých netopýrů a čeledi mystacinovití. Biotop mystaciny představují husté novozélandské pralesy od oceánského pobřeží až po hranici lesa do nadmořských výšek okolo 1000 m.
 Ilustrace mystaciny novozélandské | |
| Stupeň ohrožení podle IUCN | |
 zranitelný[1] | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Podkmen | obratlovci (Vertebrata) |
| Třída | savci (Mammalia) |
| Řád | letouni (Chiroptera) |
| Podřád | netopýři (Microchiroptera) |
| Čeleď | mystacinovití (Mystacinidae) |
| Rod | mystacina (Mystacina) |
| Binomické jméno | |
| Mystacina tuberculata Gray, 1843 | |
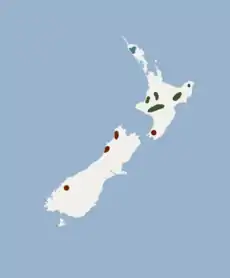 Areál rozšíření:
M. t. aupourica
M. t. rhyacobia
M. t. tuberculata Areál rozšíření:
M. t. aupourica
M. t. rhyacobia
M. t. tuberculata | |
| Synonyma | |
Mystacina velutina Hutton, 1871 | |
| Některá data mohou pocházet z datové položky. | |
Potravu si shání v letu nebo na zemi, kde se pohybují po čtyřech končetinách, což je v rámci netopýrů výjimečné (jediným dalším známým netopýrem, který využívá k pohybu po zemi všechny čtyři končetiny, je upír obecný). Při pohybu po zemi jsou mystaciny velmi čilé a dokáží vyvinout rychlost téměř 1 m/s. Druh má nejširší záběr jídelníčku ze všech netopýrů – pojídá jakékoliv dostupné členovce, dále ovoce, semínka a pyl. Mystaciny žijí v koloniích na stanovištích v dutinách prastarých pabuků. Kolonie může mít stovky až tisíce jedinců a svá dutinová stanoviště pravidelně střídá, v létě častěji než v zimě. Mystaciny se rozmnožují v létě, i když období námluv začíná již počátkem jara. Samice rodí jen jedno mládě, které nejdříve saje mateřské mléko, avšak po 6 týdnech se osamostatňuje a opouští mateřské stanoviště.
Před lidským osídlením Nového Zélandu ve 13. století byly mystaciny široce rozšířené po celém území novozélandské pevniny i přilehlých ostrovech. Odlesňování Maory a později evropskými kolonisty však mělo na jejich populaci drastický vliv a z desítek milionů jedinců klesla populace mystacin na cca 30–50 tisíc netopýrů. Vedle přirozených predátorů, jako je ostříž novozélandský a sovka bubuk, ohrožuje mystacinu především úbytek a degradace původních lesů a částečně i zavlečení predátoři (především krysy). S mystacinami žije v symbióze unikátní muška Mystacinobia zelandica.
Taxonomie
První exemplář mystaciny novozélandské pro vědecké účely byl polapen během druhé plavby Jamese Cooka (1772–1774). Netopýra odchytil Cookův přírodovědec Johann Reinhold Forster ve fjordu Queen Charlotte Sound / Tōtaranui. Formálně byl druh popsán až v roce 1843 britským zoologem Johnem Edwardem Grayem. Gray nově popsaný druh pojmenoval jako Vespertilio tuberculatus a zařadil jej do čeledi Vespertilionidae. Teprve později byl druh vyzdvihnut do samostatné čeledi mystacinovití (Mystacinidae) a rodu mystacina (Mystacina).[4]
Rod mystacina zahrnuje dva druhy, a to mystacinu novozélandskou a mystacinu větší, která je s největší pravděpodobností vyhynulá. Jelikož rod mystacina je jediný rod čeledi mystacinovití, představuje mystacina novozélandská jediného žijícího zástupce celé čeledi.[5] Mystacinovití zahrnují i několik fosilních druhů.[6]
Mystacinovití náleží do nadčeledi Noctilionoidea, prastaré skupiny netopýrů z období Gondwany, ze které se mystacinovití vydělili někdy před 51–41 miliony let.[7] Lze se však setkat s udávaným rozpětím vydělení od 35 až do 68 milionů let. Každopádně to je až poté, co se novozélandský kontinent vydělil z Gondwany (circa před 80 miliony lety).[8] Nejbližší příbuzné mystacinovitých patrně představuje široce rozšířená čeleď listonosovitých.[9]
Česky se Mystacina tuberculata označuje jako mystacina novozélandská, zemolez novozélandský či netopýr novozélandský. Petr Benda nicméně navrhl, aby se jako netopýr novozélandský označoval spíše Chalinolobus tuberculatus (New Zealand long-tailed bat), též známý jako netopýr hrbolatý.[3]
Poddruhy
.jpg.webp)
Rozlišují se tři poddruhy:[10][11]
- M. t. tuberculata – vyskytuje se na ostrově Whenua Hou (západně od Stewartova ostrova), severozápadně od Nelsonu a ve Fiordlandu;
- M. t. auporica – Northland a ostrov Hauturu;
- M. t. rhyacobia – Northland, střední část Severního ostrova a Taranaki.
Toto rozdělení druhu na tři poddruhy je však problematické.[8] Studie DNA od Briana D. Lloyda z Ministerstva památkové péče z roku 2003 totiž upozorňuje na to, že rozdělení na tři poddruhy bylo založeno na geografickém rozšíření a jemných morfologických odlišnostech, zejména délce předloktí. Ta se však u jednotlivých poddruhů značně překrývá a analýza mitochondriální DNA identifikovala v rámci druhu hned šest linií. Lloyd proto navrhl taxon rozdělit buďto na šest poddruhů nebo alespoň na šest evolučně významných jednotek.[10]
Popis
Jedná se o malý až středně velký druh netopýra s podsaditým tělem. Hedvábná srst je velmi hustá, krátké chlupy měří kolem 7 mm. Hřbetní srst je šedavá až tmavě hnědá, zatímco srst na břiše je světlejší. Neosrstěné oblasti jako jsou uši, křídla, nos, nohy a ocas odhalují šedo-hnědou kůži.[2]
Tělo dosahuje délky 60–70 mm, s roztaženou blánou mezi zadními končetinami (uropatagium) je ještě o 20 mm delší. Kondylobazální délka lebky (vzdálenost od mezičelisti po týlní výběžky) je kolem 17–19 mm. Protáhlá lebka je zhruba dvakrát delší než širší. Špičaté uši bývají 17,5–19,1 mm dlouhé a ve spodní část jsou široké 9 mm. Tragus (část vnějšího ucha) je zašpičatělý a s délkou 10 mm poměrně dlouhý. Čelo je strmé. Na čenichu kuželovitého tvaru jsou nápadné nosní dírky oválného tvaru, které směřují dopředu. Kolem čenichu a úst vyrůstají nápadná hmatová pera (vibrisy). Robustní řezáky jsou výrazné a špičaté. Zubní vzorec je I 1/1, C 1/1, Pm 2/2, M 3/3, tedy 28 zubů.[2][5] Pohlavní dimorfismus je nevýrazný; samice bývají v průměru o cca 4,3 % těžší než samci a také mívají v průměru o zhruba 0,9 % delší předloktí než samci.[8]
Ocas je dlouhý až 12 mm a vyčuhuje kolem 6–8 mm za uropatagium. Rozpětí křídel je 280–300 mm. Délka předloktí je 37–46,9 mm, holenní kost měří 14,5–17 mm. Statné nohy dosahují délky až 6 mm, kůže na chodidlech je volná a vrásčitá. Dospělí jedinci váží 10–22 g, avšak po lovu může jejich váha narůst o 20–30 %. Na podzim před hibernací narůstá výchozí váha (tzn. váha před lovem) rovněž o 20–30 %. Březí samice mohou před porodem vážit až o 35 % více než obyčejně.[5]
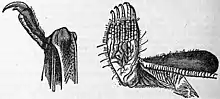
Jazyk je dlouhý kolem 12 mm,[2] přičemž mystaciny ho dokáží vypláznout až 5 mm za špičku čenichu.[12] Na konci jazyka se nachází malé výběžky připomínající ochlupení, které netopýrovi pomáhají v pojídání nektaru. Mezi jedinečné vnější znaky patří dobře definovaná, tlustá, vroubkovaná proximální (bližší k tělu) oblast křídelních membrán a malé bazální spáry (typ prstového zakončení běžný u dravců), které se nachází u kořene spodní strany zaoblených drápů na předních i zadních končetinách. Tyto malé spáry jsou patrně uzpůsobeny pro lezení a plazení. V rámci netopýrů jsou přítomné pouze u rodu mystacina.[2]
.jpg.webp) Vycpaný exemplář, na kterém je patrná délka uropatagia (blána mezi zadními končetinami)
Vycpaný exemplář, na kterém je patrná délka uropatagia (blána mezi zadními končetinami) Portrét – nápadné jsou velké ušní boltce a nosní dírky směřující dopředu
Portrét – nápadné jsou velké ušní boltce a nosní dírky směřující dopředu Mystacina za letu s patrným vroubkováním na membráně, které je pro rod typické
Mystacina za letu s patrným vroubkováním na membráně, které je pro rod typické
Chování
Život v koloniích

Mystaciny žijí samotářsky i v koloniích. Kolonie si nachází svá stanoviště v dutinách stromů a zahrnují desítky až tisíce jedinců (největší zaznamenaná kolonie mystacin měla přes 6 tisíc netopýrů[13]). Kolonie je takřka nemožné najít ve volné přírodě bez potřebné radiové techniky, protože na 100 km² těžko přístupného pralesa připadají jen dvě, maximálně tři kolonie. Strom, v jehož dutině se nachází netopýří kolonie, navíc často postrádá vnější znaky jejich přítomnosti netopýrů. V některých případech lze kolem kmene stromu nalézt pižmově zapáchající netopýří guáno, které může dosahovat výše přes 1 m. Často se však guano nachází přímo v dutině stromu a není možné ho zpozorovat. Občas může přítomnost kolonie prozradit nápadné cvrlikání, které netopýři používají pro sociální komunikaci. Ani to však není pravidlem, protože netopýří kolonie může zrovna hřadovat (nocovat / odpočívat) nebo může vchod do kolonie, odkud se cvrlikání šíří, být vysoko nad zemí.[14]
Kolonie bývají takřka výhradně v dutinách živých stromů, a pouze pokud strom uhyne, kolonie občas nadále obývá i mrtvý strom. V jeskyních nebyly mystaciny zaznamenány, i když fosilní nálezy jejich koster potvrzují, že mystaciny při nejmenším v minulosti využívaly jeskyně za svá stanoviště.[15] Kolonie mystacin má na svém území velké množství dutinových stanovišť, které často střídá, avšak dříve či později se vrátí na své starší stanoviště. Kolonie užívá stejnou dutinu desítky a potenciálně i stovky let. V létě dochází ke střídání stanovišť každých několik dnů až týdnů, v zimě se jedná spíše o týdny až měsíce. Zatímco počátkem léta populace mystacin v určité lokalitě obývá pouze dvě či tři stanoviště najednou, jindy jsou jejich kolonie mnohem menší (tzn. populace je rozdělena do menších kolonií obývajících více stanovišť) nebo mystaciny hřadují samostatně (především v zimě). K přestěhování kolonie dochází velmi rychle a vždy se přesune drtivá většina jedinců. Není úplně jasné, z jakého podnětu k přesunu dochází, avšak vypadá to, že na stěhování netopýrů má vliv roční období. Pokud dojde k vyrušení kolonie predátory nebo člověkem, může se stát, že mystaciny stromové stanoviště nadobro opustí.[16]

Mystaciny zůstávají přes den v koloniích, kde většinu dne spí. Vylétávají za potravou až po setmění a navrací se před rozbřeskem.[14] Vchod do kolonie tvoří díra v průměru nejméně 15 mm (minimální šíře na to, aby jí mystaciny prolezly[17]), avšak může se jednat i o obrovské otvory dlouhé 3 m a široké až kolem 1 metru. Dutinu uvnitř stromu si někdy upravují do požadovaného tvaru s pomocí ostrých zubů (byly pozorovány vykousané chodby až do délky 1 m[18]). V případě samostatného hřadování mystaciny obývají nejen dutiny stromů, ale i malé prostory pod kůrou stromů a stromových kapradin, otvory v zemi nebo si své stanoviště najdou pod kořeny stromů. Klíčovým důvodem, proč se mystacina rozhodne hřadovat samostatně nebo v kolonii, je patrně otázka energetické efektivity. V zimě, kdy mystaciny často hibernují, je běžné samostatné hřadování z důvodu možného narušení hibernace ostatními jedinci v kolonii, čímž dochází ke zvýšenému energetickému výdeji. Naproti tomu v létě se netopýři spíše shlukují do kolonií, kde jsou jedinci natěsnáni blízko vedle sebe, čímž narůstá teplota kolonie a netopýři nemusí vydat tolik energie na regulaci své teploty.[16]
Rozmnožování
Od září do prosince probíhají u mystacin námluvy. Samci vábí samice svým vysoce položeným zpěvem doplněným o ultrazvukovou hladinu. V tomto období si samci vybírají k hřadování menší pukliny v kmenech menších stromů, jejichž vchod potřou hnědým pižmově zapáchajícím sekretem z krčních žláz, což pomáhá k nalákání samic. Samice jsou schopny uletět až 10 km, aby navštívily samce v jeho pářicím stanovišti. Samci často používají stejná stanoviště každý rok a občas je musí bránit proti ostatním dorážejícím samcům. K páření dochází od ledna do května. Po kopulaci se samicím vytvoří vaginální zátka (sphragis). K zabřeznutí však dochází až o několik měsíců později blíže neurčeným typem odložené nidace. Samice rodí pouze jedno mládě, a to od poloviny března do poloviny ledna, nicméně v rámci jedné kolonie se většina mláďat narodí během jednoho týdne.[19][20]
Mláďata se rodí v malých dutinách stromů buď přímo v mateřské kolonii, nebo mimo ni. Pokud se mládě narodí mimo mateřskou kolonii, samice ho tam brzy po narození přenese. První cca 2 týdny samice zůstávají s mláďaty během dne a v noci se vydávají lovit, avšak několikrát za noc se vrátí do kolonie, aby své mládě nakojily mateřským mlékem. Po zhruba dvou týdnech začnou samice přes den hřadovat mimo kolonii mláďat a vrací se krmit své potomky v noci v méně častých intervalech. Někdy s kolonií mláďat hřadují i samci či nepářící se samice.[21][22]
Po narození mláďata váží kolem 5 g, délka předloktí dosahuje kolem 19 mm. Jejich oči jsou zavřené na další dva týdny. Mláďata nemají srst a jejich tělo pokrývá zvrásněná volná kůže, která je na zádech šedočerná a na přední části těla světle šedá až bělavá.[22] Rodí se s mléčnými zuby, které jsou kolem 3. týdne nahrazeny stálým chrupem. Ve 3. týdnu se mláďata začnou učit létat a o týden později již začínají létat kolem mateřského stanoviště. Někdy v této době se plně osrstí. V 6 týdnech juvenilní jedinci opouští mateřské stanoviště a v 8. týdnu je jejich kostra plně vyvinutá. Váhy dospělce dosahují po několika měsících.[21]
Každoročně se páří kolem 80 % pohlavně dospělých samic.[22]
Regulace tělesné teploty
Mystaciny jsou heterotermní, čili regulují svou tělesnou teplotu buďto samy, nebo s pomocí teploty okolního prostředí. Torpor (krátkodobý stav útlumu životních funkcí) využívají na denní bázi během samostatného hřadování; při pobytu v koloniích k torporu nedochází.[23] Během torporu tělesná teplota netopýrů klesne takřka na teplotu jejich okolního prostředí. Probuzení z torporu trvá 10–20 minut.[24]
.jpg.webp)
Do hibernace (dlouhodobý stav útlumu životních funkcí) mystaciny upadají v zimním období, případně v době nedostatku potravy. Období hibernace závisí na lokaci, např. v lese Rangataua při okraji národního parku Tongariro období hibernace nastává od konce dubna do poloviny října. Nicméně v době dostatku potravy se počátek období hibernace může oddálit. Během hibernačního období netopýři tlumí své životní funkce na 2–10 dní. Poté se na několik hodin až nocí probudí, aby si nalovili potravu. Nejčastěji k probuzení dochází během vlhkých období, kdy bezobratlí živočichové vylézají na povrch a pro mystaciny je snadné je polapit.[25]
Potrava
Mystaciny jsou převážně hmyzožravé, hojně však pojídají i nektar, pyl a ovoce. Mystacina novozélandská je jediný známý zástupce podřádu netopýrů v mírném podnebném pásu, který pojídá a opyluje rostliny. Výjimečná je i v rámci letounů pro svou velkou rozmanitost jídelníčku. V rámci netopýrů má vůbec nejpestřejší jídelníček.[26][27]
Klíčovou složku potravy tvoří létaví i nelétaví členovci, jako jsou pavouci, roztoči, sekáči, stonožkovci, švábi, polokřídlí, třásnokřídlí, síťokřídlí, brouci, dvoukřídlí, motýli, blanokřídlí a další. Mystaciny chované v zajetí sežerou v podstatě jakékoliv členovce, kterými jsou krmeny. Preferují členovce s měkkým tělem, které pořádají typicky tak, že nejdříve jim zuby ucvaknou končetiny a křídla a poté zkonzumují šťavnatá těla.[28] K nalezení hmyzu používají čich a pasivní naslouchání (při lovu na zemi) a echolokaci (během lovu za letu). Pokud mystacina lokalizuje kořist za letu, dosedne těsně vedle ní a teprve poté se na ni vrhne. Při hledání potravy v lesním podrostu se mystaciny často celé ponoří do nánosu listí a pouze občasně se na chvíli „vynoří“ zpět na povrch. Při lovu létajícího hmyzu dokáží mystaciny pozměnit echolokační frekvenci takovým způsobem, aby dostávaly co nejvíce informací o pohybující se kořisti. Zvuky, které přitom vydávají, jsou slyšitelné lidskému uchu a připomínají vysoko položené bzučení. Jelikož toto bzučení je u netopýrů chovaných v zajetí i v blízkosti kolonií ve volné přírodě poměrně vzácné; předpokládá se, že mystaciny hmyz za letu příliš neloví a specializují se na hmyz na zemi.[12] Během lovu stráví okolo 30 % času sběrem hmyzu na zemi.[29]

Mystaciny takřka nikdy neopouští les, avšak během shánění potravy jsou schopné přeletět otevřené paseky či louky až do vzdálenosti 2 km a na ostrovech Whenua Hou a Hauturu byly pozorovány případy sběru potravy mimo les na loukách a skalních útesech.[22] Rostliny, na kterých hodují, jsou typické třemi charakteristikami, a sice 1) zřetelně viditelným pylem; 2) hojností květů v plném květenství; 3) v případě přítomnosti nektaru je tato cukernatá šťáva snadno dostupná jazykem. Hojně konzumován je pyl místních rostlin Knightia excelsa, Collospermum hastatum a Collospermum microspermum.[12] V zajetí netopýři sežerou velké množství druhů pylu nejen z místních, ale i z exotických rostlin.[28]
Netopýři se patrně živí i některými druhy hub a výtrusy.[29] V zajetí pravidelně konzumují během jediné noci 5–7 g hmyzu (tzn. 36–50 % tělesné váhy před krmením), ve volné přírodě se toto množství odhaduje na 5,6 g.[30]
Pohyb
Mystacinám k pohybu po zemi dobře slouží jejich silné zadní končetiny. Membrána mezi prsty a u ocasu se dokáže srolovat, takže jim nepřekáží v pohybu. Vedle silných noh jsou pro pohyb po zemi adaptovány i přední končetiny, jejichž zápěstí mystaciny používají místo předních noh.[29] Při pohybu po zemi jsou mystaciny velmi rychlé a na rozdíl od lovu ve vzduchu nepoužívají k hledání hmyzu na zemi echolokaci, nicméně využívají ji k orientaci v prostoru.[31] Mystacina novozélandská spolu s upírem obecným jsou jedinými známými netopýry, kteří při pohybu po zemi využívají principy chůze, resp. pohybu po čtyřech končetinách (kvadrupedie).[32] Maximální zaznamenaná rychlost pohybu mystacin po zemi byla 0,95 m/s.[22]
Při letu se mystaciny pohybují typicky pod korunami vzrostlých stromů, ale nad podrosty spodního patra. Dokáží vyvinou rychlost až 60 km/h, avšak v hustě zarostlých původních pralesích se většinou pohybují pomaleji. Dokáží se obratně vyhýbat překážkám a jsou schopny vzlétnout takřka vertikálně bez potřeby horizontálního pohybu, což přijde vhod v pohybu v hustém houští.[29]
Echolokace
Echolokace mystacin je krátká, frekvenčně modulační, více harmonická a s nízkou amplitudou.[31] Zahrnuje tři a někdy i čtyři harmonické řady. První harmonie má frekvenci c. 20–37 kHz, druhá 40–60 kHz, třetí 66–90 kHz a čtvrtá harmonie, pokud je přítomná, se pohybuje ve frekvencích 85–105 kHz. Jednotlivá volání trvají pouze 1–3,9 ms.[17] Echolokace mystacin je podobná echolokaci netopýrů z čeledi listonosovití, což může odrážet blízkou fylogenetickou příbuznost mezi čeledi mystacinovitých a listonosovitích.[31]
Výskyt
Historické rozšíření
Mystaciny se původně vyskytovaly na většině území Nového Zélandu. Vedle historických zmínek o tom svědčí hlavně fosilní nálezy z jeskyň, závrtů a skalních převisů z oblastí Severního (Waikato, Hawke's Bay, Wairarapa) i Jižního ostrova (Canterbury, severozápadní Nelson, národní park Fiordland).[33] Na základě genetických dat se odhaduje, že jen ve střední části Severního ostrova bylo před lidským osídlením Nového Zélandu přítomno nejméně 12,5 milionů mystacin.[21]
Současné rozšíření

Současná populace je značně fragmentovaná[34] a většina mystacin se vyskytuje na Severním ostrově. Tam je netopýr rozšířen zejména v lesích Omahuta a Pukeki (Northland) a v páse původních lesů od Taranaki směrem na východ po East Cape a dále na jih po jižní oblasti pohoří Tararua. Konkrétně se jedná např. o lesy a parky Waitaanga, Pureora, Waitotara, Rangataua, Kaimanawa, Whirinaki a Urewera. Na Jižním ostrově se mystaciny nachází pouze na několika známých lokacích, a to v nížině Oparara (severozápadní Nelson) a v národním parku Fiordland.[35] Druh je přítomen i na menších novozélandských pobřežních ostrovech jako je Whenua Hou a Hauturu[36][13] a Ministerstvo památkové péče v roce 2005 přemístilo 20 netopýrů na ostrov Kapiti, kde se pokouší o založení nové populace.[33]
Celková populace mystacin se odhaduje na 30[13]–50[37] tisíc. Jen několik kolonií přitom přesahuje jeden tisíc jedinců.[13]
Biotop
Mystaciny jsou přítomny v různorodých biotopech od pobřežních oblastí (Northland, ostrovy Whenua Hou a Hauturu) až po vyšší nadmořské výšky po hranici lesa (kolem 1000 m na Severním ostrově a cca 550 m na Jižním ostrově). Pro kolonie je klíčová přítomnost původních pralesů se starými vzrostlými stromy (obvod alespoň 1 m a výška alespoň 25 m) s dostatečně velkými dutinami pro přítomnost kolonie.[27] Takové lesy se vyznačují dominancí pabuků, damaroňů, nohoplodů a stromů rodu Dacrydium.[32] Ke konkrétním druhům stromů, v jejichž dutinách byly nalezeny stanoviště mystacin, patří damaroň jižní, Vitex lucens, Prumnopitys taxifolia, Nothofagus fuscus, Nothofagus truncata, Nothofagus menziessi, Podocarpus hallii, Metrosideros umbellata, Weinmannia racemosa a Dacrydium cupressinum.[38]
Typické kolonie bývají na závětrném, teplém a úrodném stanovišti, dostatečně chráněném proti dešti. Netopýří kolonie se nevyskytují v oblastech s jasnou přítomností člověka (např. v blízkosti oblastí vypalování strání, těžby dřeva či vysokého počtu introdukovaných savců). Výjimku tvoří pouze jedinci, kteří při hledání potravy občas zavítají do druhotných lesů, borovicových plantáží a do oblasti farem.[27]
Ohrožení

Největší hrozbu pro populace mystacin představuje ztráta původních pralesů. S jejich vypalováním započali nejdříve Maorové (zejména na Jižní ostrově) a poté s ním pokračovali evropští kolonisté. Dnes již k vypalování lesů nedochází v takové míře jako v minulosti, nicméně původní lesy stále ubývají, ať už přímou aktivitou člověka (zejména získávání orné půdy) nebo následkem šíření zavlečených organismů (např. fytoftorové onemocnění způsobené patogenem Phytophthora agathidicida, který napadá prastaré damaroně jižní).[39]
Další klíčovou hrozbou mystacin jsou introdukovaní savčí predátoři, a to především krysy, hranostaji a zdivočelé kočky domácí. Mystaciny jsou zvláště náchylné k predaci v době sbírání potravy na zemi a během torporu, kdy jim trvá dlouhou dobu se probrat a v případě ohrožení i uletět. Mystaciny větší vymizely z novozélandské pevniny následkem invaze krys ostrovních a z posledních potvrzených míst výskytu, malých pobřežních ostrůvků Taukihepa a Rerewhakaupoko, vymizely po introdukci krys obecných.[39][40] Ohrožení představují i kusu liščí – byl zaznamenán případ, kdy se kusu snažil dostat k mláďatům v kolonii mystacin.[41]
Vliv velkých introdukovaných savců (dobytka, ovcí, koz, prasat, jelenů) na populaci mystacin není zcela zřejmý. Nicméně tito zavlečení savci mají negativní vliv na novozélandské pralesy, a tím patrně mohli způsobit snížení populace v některých částech Nového Zélandu, nezdá se však pravděpodobné, že by mohli způsobit vymření celé populace.[42]
Přirozené predátory netopýrů představují ostříž novozélandský a sovka bubuk. V minulosti mystaciny lovila i dnes vyhynulá sovka bělolící.[43]
Parazité a symbiont

Na rozdíl od řady jiných netopýrů mystaciny nehostí žádné parazitické blechy. Parazituje na nich nepopsané klíště z rodu klíšťák (Argas). Vedle toho mystaciny hostí i několik druhů nepopsaných roztočů.[44]
Mystaciny žijí v symbiotickém vztahu s muškou Mystacinobia zelandica, která představuje jediného zástupce čeledi Mystacinobiidae. Tato muška měří 4–9 mm. Na konci jejích pavoučích noh se nachází drápky, které pomáhají muškám při pohybu po netopýří srsti. Na rozdíl od ostatních mušek spojovaných s netopýry (parazitické mušky z čeledi muchulovití), se Mystacinobia zelandica neživí netopýří krví, ale guánem, čímž urychluje jeho rozklad. Mušky se navíc starají mystacinám o jejich srst, ze které vybírají zbytky jídla včetně pylu a mateřského mléka. Většinou jedna muška připadá na jednoho netopýra, avšak byl zaznamenán případ 60 mušek na jediné mystacině.[44][45][46]
Ochrana
Vedle snahy ochranářů o zpomalení úbytku původních pralesů patří k hlavním způsobům ochrany výzkum a regulace nepůvodních predátorů, zprostředkovaná především pomocí pesticidu 1080. Vzhledem k obrovské ploše území, na kterém je třeba predátory regulovat, není reálné klást a denně kontrolovat dostatečné množství fyzických pastí přímo v terénu,[41] a tak se predátoři hubí hlavně pesticidem 1080 shazovaným z vrtulníků v podobě granulí. Pesticid nicméně představuje nebezpečí i pro samotné mystaciny, které má v první řadě chránit. Byly zaznamenány případy otravy mystacin pesticidem 1080 – a to buď po přímém požití jedu nebo na následky sekundární otravy (tzn. požití něčeho otráveného, patrně členovců).[39][47]
Mezinárodní svaz ochrany přírody hodnotí mystacinu novozélandskou jako zranitelný taxon s tím, že populace mystacin je klesající.[13] Ministerstvo památkové péče vyhodnocuje každý ze tří poddruhů zvlášť a jejich status je následující:[48]
- M. t. tuberculata – zotavuje se (Recovering)
- M. t. auporica – národně zranitelný (Nationally Vulnerable)
- M. t. rhyacobia – na ústupu (Declining)
Vztah k lidem
Maorové (první obyvatelé Nového Zélandu) nazývali všechny novozélandské netopýry „peka peka“. Považovali je za ptáky noci, jejichž úkolem bylo provázet duše zemřelých do podsvětí (Rarohenga). Maorové netopýry s oblibou jedli. Vykuřovali je z jeskyní a kmenů stromů, aby je pak po tisících chytali a hodovali na nich. První netopýr, který byl chycen během loveckých výprav, byl rituálně usmrcen a nabídnut jako oběť bohovi lesa a ptáků Tānemu.[49]
Odkazy
Reference
- Červený seznam IUCN ohrožených druhů 2021.3. 9. prosince 2021. Dostupné online. [cit. 2021-12-22]
- Lloyd 2005, s. 110.
- BENDA, Petr. České vernakulární jmenosloví netopýrů. I. Návrh úplného jmenosloví. Vespertilio. 2010, čís. 13–14, s. 263–308. Dostupné online [cit. 2021-04-06]. ISSN 1213-6123.
- Winnington 2000, s. 2.
- CARTER, Gerald G.; RISKIN, Daniel K. Mystacina tuberculata. S. 1. Mammalian Species [online]. 2006-05 [cit. 2020-11-11]. Roč. 790, s. 1. DOI 10.1644/790.1. (anglicky)
- HAND, Suzanne J.; WORTHY, Trevor H.; ARCHER, Michael. Miocene mystacinids (Chiroptera, Noctilionoidea) indicate a long history for endemic bats in New Zealand. Journal of Vertebrate Paleontology. 2013-11, roč. 33, čís. 6, s. 1442–1448. Dostupné online [cit. 2020-11-21]. ISSN 0272-4634. DOI 10.1080/02724634.2013.775950. (anglicky)
- MILLER-BUTTERWORTH, Cassandra M.; MURPHY, William J.; O'BRIEN, Stephen J.; JACOBS, David S.; SPRINGER, Mark S.; TEELING, Emma C. A Family Matter: Conclusive Resolution of the Taxonomic Position of the Long-Fingered Bats, Miniopterus. S. 1558. Molecular Biology and Evolution [online]. 2007-07. Roč. 24, čís. 7, s. 1558. DOI 10.1093/molbev/msm076. (anglicky)
- Lloyd 2005, s. 113.
- KENNEDY, Martyn; PATERSON, Adrian M.; MORALES, Juan Carlos. The Long and Short of It: Branch Lengths and the Problem of Placing the New Zealand Short-Tailed Bat, Mystacina. Molecular Phylogenetics and Evolution. 1999-11, roč. 13, čís. 2, s. 405–416. Dostupné online [cit. 2020-12-08]. DOI 10.1006/mpev.1999.0660. (anglicky)
- LLOYD, Brian D. Intraspecific Phylogeny of the New Zealand Short-Tailed Bat Mystacina tuberculata Inferred from Multiple Mitochondrial Gene Sequences. S. 460–476. Systematic Biology [online]. 2003-08-01. Roč. 52, čís. 4, s. 460–476. DOI 10.1080/10635150390218187. (anglicky)
- Short-tailed bat [online]. Department of Conservation [cit. 2020-11-14]. Dostupné online. (anglicky)
- Lloyd 2001, s. 67.
- O'DONNELL, C. Mystacina tuberculata: e.T14261A4427784 [online]. The IUCN Red List of Threatened Species 2008, 2008-06-30 [cit. 2020-11-14]. DOI 10.2305/IUCN.UK.2008.RLTS.T14261A4427784.en. (anglicky)
- Lloyd 2001, s. 62.
- WORTHY, T. H.; HOLDAWAY, R.N. Quaternary fossil faunas from caves in Takaka Valley and on Takaka Hill, northwest Nelson, South Island, New Zealand. S. 297–391. Journal of the Royal Society of New Zealand [online]. 1994-09 [cit. 2020-11-17]. Roč. 24, čís. 3, s. 297–391. DOI 10.1080/03014223.1994.9517474. (anglicky)
- Lloyd 2001, s. 69–70.
- Lloyd 2005, s. 119.
- Carter & Riskin 2006, s. 5.
- Lloyd 2005, s. 121–122.
- Carter & Riskin 2006, s. 3–4.
- Lloyd 2005, s. 122.
- Carter & Riskin 2006, s. 3.
- SEDGELEY, Jane A. Roost site selection and roosting behaviour in lesser short‐tailed bats (Mystacina tuberculata) in comparison with long‐tailed bats (Chalinolobus tuberculatus) in Nothofagus forest, Fiordland. S. 227–241. New Zealand Journal of Zoology [online]. 2003-01 [cit. 2020-11-15]. Roč. 30, čís. 3, s. 227–241. DOI 10.1080/03014223.2003.9518341. (anglicky)
- Lloyd 2005, s. 120–121.
- Lloyd 2005, s. 121.
- HAND, Suzanne J.; LEE, Daphne E.; WORTHY, Trevor H.; ARCHER, Michael; WORTHY, Jennifer P.; TENNYSON, Alan J. D.; SALISBURY, Steven W. Miocene Fossils Reveal Ancient Roots for New Zealand’s Endemic Mystacina (Chiroptera) and Its Rainforest Habitat. S. e0128871. PLOS ONE [online]. 2015-06-17 [cit. 2020-11-15]. Roč. 10, čís. 6, s. e0128871. DOI 10.1371/journal.pone.0128871. (anglicky)
- Lloyd 2005, s. 116.
- Lloyd 2005, s. 116–117.
- Lloyd 2005, s. 118.
- LLOYD, B. D.; MCQUEEN, S. M. An assessment of the probability of secondary poisoning of forest insectivores following an aerial 1080 possum control operation. S. 53. New Zealand Journal of Ecology [online]. 2000 [cit. 2020-11-15]. Roč. 24, čís. 1, s. 53. Dostupné online. ISSN 0110-6465. (anglicky)
- JONES, Gareth; WEBB, Peter I.; SEDGELEY, Jane A.; O’DONNELL, Colin F. J. Mysterious Mystacina: how the New Zealand short-tailed bat (Mystacina tuberculata) locates insect prey. S. 4209–4216. Journal of Experimental Biology [online]. 2003-12-01 [cit. 2020-11-21]. Roč. 206, čís. 23, s. 4209–4216. DOI 10.1242/jeb.00678. (anglicky)
- HAND, Suzanne J; WEISBECKER, Vera; BECK, Robin MD; ARCHER, Michael; GODTHELP, Henk; TENNYSON, Alan JD; WORTHY, Trevor H. Bats that walk: a new evolutionary hypothesis for the terrestrial behaviour of New Zealand's endemic mystacinids. S. 2. BMC Evolutionary Biology [online]. 2009. Roč. 9, čís. 1, s. 2. DOI 10.1186/1471-2148-9-169. (anglicky)
- Carter & Riskin 2006, s. 2.
- O'DONNELL, C. Mystacina tuberculata: e.T14261A4427784 [online]. The IUCN Red List of Threatened Species 2008, 2008-06-30 [cit. 2020-11-14]. DOI 10.2305/IUCN.UK.2008.RLTS.T14261A4427784.en. (anglicky)
- JACKSON, Bex. Murchison Mountains Lesser Short Tailed Bat Monitoring [online]. Department of Conservation, 2019 [cit. 2020-11-14]. Dostupné online. (anglicky)
- Lloyd 2005, s. 114.
- Lloyd 2005, s. 123.
- Lloyd 2005, s. 120.
- MOLLOY, Janice. Bat (Peka peka) recovery plan (Mystacina, Chalinolobus) [online]. Wellington, N.Z.: Threatened Species Unit, Dept. of Conservation [cit. 2020-11-17]. S. 7. Dostupné online. ISBN 0-478-01570-4. (anglicky)
- Lloyd 2005, s. 129.
- Helping our native New Zealand bats [online]. Department of Conservation, 2015 [cit. 2020-11-17]. Dostupné online. (anglicky)
- LLOYD, B. D. Advances in New Zealand mammalogy 1990–2000: Short‐tailed bats. S. 64. Journal of the Royal Society of New Zealand [online]. 2001-03 [cit. 2020-11-17]. Roč. 31, čís. 1, s. 64. DOI 10.1080/03014223.2001.9517639. (anglicky)
- Lloyd 2005, s. 124.
- Lloyd 2005, s. 125.
- HOLLOWAY, Beverley A. A new bat‐fly family from New Zealand (Diptera: Mystacinobiidae). S. 279–301. New Zealand Journal of Zoology [online]. 1976-12 [cit. 2020-11-18]. Roč. 3, čís. 4, s. 279–301. DOI 10.1080/03014223.1976.9517919. (anglicky)
- HUNT, Roy. Batfly [online]. New Zealand Geographic [cit. 2020-11-23]. Dostupné online. (anglicky)
- DENNIS, Gillian C.; GARTRELL, Brett D. Nontarget mortality of New Zealand lesser short-tailed bats (Mystacina tuberculata) caused by diphacinone. S. 177–186. Journal of Wildlife Diseases [online]. 2015-01 [cit. 2020-11-22]. Roč. 51, čís. 1, s. 177–186. DOI 10.7589/2013-07-160. (anglicky)
- O'DONNELL, Colin F. J.; BORKIN, K.M.; CHRISTIE, J.E.; LLOYD, B.; PARSONS, S.; HITCHMOUGH, R.A. Conservation status of New Zealand bats, 2017 [online]. Wellington, New Zealand: 2017 [cit. 2020-11-17]. Dostupné online. ISBN 9781988514529. (anglicky)
- WINNINGTON, Andrew Paul. Ecology, genetics and taxonomy of Peka peka (Chiroptera: Mystacina tuberculata and Chalinolobus tuberculatus). , 2000. Doctor of Philosophy thesis. University of Otago. Vedoucí práce Hamish Spencer & David Lambert. s. 1. Dostupné online.
Literatura
- LLOYD, B., 2005. The handbook of New Zealand mammals. Příprava vydání Carolyn M. King. South Melbourne, Victoria, Australia: Oxford University Press. ISBN 0195584775. Kapitola New Zealand lesser short-tailed bat, s. 110–126. (anglicky)
Externí odkazy
 Obrázky, zvuky či videa k tématu mystacina novozélandská na Wikimedia Commons
Obrázky, zvuky či videa k tématu mystacina novozélandská na Wikimedia Commons  Taxon Mystacina tuberculata ve Wikidruzích
Taxon Mystacina tuberculata ve Wikidruzích- Mystacina novozélandská na webu Ministerstva památkové péče
- Video zachycující pohyb mystacin po zemi