Citrátový cyklus
Citrátový cyklus (cyklus kyseliny citrónové, cyklus trikarboxylových kyselin, Krebsův cyklus) je řada reakcí, které tvoří společnou metabolickou dráhu při aerobní oxidaci sacharidů, lipidů a proteinů.
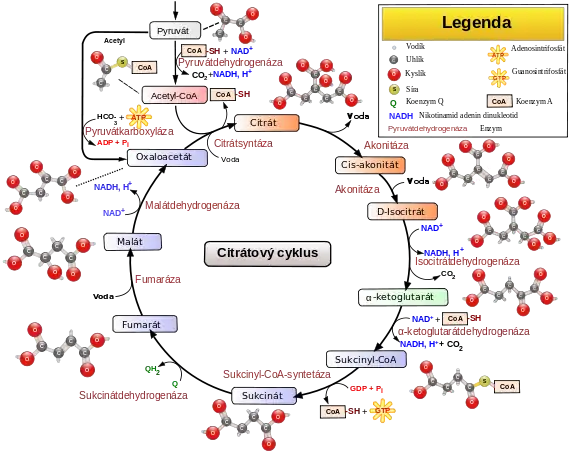
Postupnou dekarboxylací a oxidací šestiuhlíkaté kyseliny citrónové uvolňuje redukční ekvivalenty, které jsou použity při oxidativní fosforylaci k syntéze ATP, hlavního energetického zdroje buňky. Citrátový cyklus hraje klíčovou roli i v dalších metabolických dějích, jako je glukoneogeneze, transaminace, deaminace nebo lipogeneze. Některé reakce cyklu proto probíhají i v buňkách, které nemají aerobní metabolismus. V prokaryotických buňkách probíhá v cytosolu, u eukaryot se enzymy citrátového cyklu vyskytují v mitochondriích a to buď volně v mitochondriálním matrixu, nebo zakotvené k vnitřnímu povrchu vnitřní mitochondriální membrány.
Historie výzkumu
Citrátový cyklus se také nazývá cyklus Krebsův, na počest sira Hanse Adolfa Krebse (1900–1981). Ten již v roce 1932 objevil močovinový cyklus a dobře si tedy uvědomoval, že biochemické děje v těle často pracují v cyklech. V Krebsově době bylo známo, že se glukóza před svým rozkladem na oxid uhličitý mění na pyruvát, ale další sled reakcí byl neznámý. Měl k dispozici čerstvou holubičí svalovou tkáň, do které aplikoval různé organické látky a sledoval, zda se spalují na CO2. Ve třicátých letech na tomto tématu pracovali další vědci a objasňovali dokonce některé dílčí metabolické kroky. Až Krebse však napadlo, že by se mohl oxalacetát regenerovat vazbou na nějaký produkt rozkladu potravy a kondenzovat s ním na citrát, čímž by se uzavřel cyklus. Jeho práci však odmítl renomovaný časopis Nature, nicméně nakonec svou revoluční teorii v roce 1937 publikoval. Jeho žáci v několika dalších letech Krebsovu teorii prokázali pomocí radioaktivně značeného uhlíku. V padesátých letech byl cyklus upřesněn o acetylkoenzym A a v roce 1953 byla Krebsovi za objev citrátového cyklu udělena Nobelova cena za fyziologii a medicínu.[1]
Průběh reakce
Přehled
Produktem katabolismu živin je acetyl-CoA, acetylový (dvouuhlíkatý) zbytek navázaný na koenzym A. K syntéze koenzymu A je potřeba vitamín B5, kyselina pantothenová.
- Acetyl-CoA vstupuje do cyklu a kondenzuje s oxalacetátem (4 uhlíky) na citrát - kyselinu citrónovou (6 uhlíků).
- Postupné dekarboxylace a oxidace uvolňují oxid uhličitý jako odpadní produkt, hlavně ale dochází k redukci koenzymů. Konečným produktem je opět oxalacetát.
- Oxalacetát kondenzuje s dalším acetyl-CoA a cyklus se opakuje
Protony nesené redukovanými koenzymy jsou použity v dýchacím řetězci, kdy oxidativní fosforylací dochází k syntéze ATP, molekuly, která je hlavním zdrojem energie pro veškeré endotermní reakce probíhající v buňce.
Podrobnější pohled
Citrátový cyklus se dá rozdělit do několika fází, například takto:
1. acetyl-CoA + oxalacetát + H2O → citrát + CoA-SH
Jedná se o kondenzační reakci, kterou katalyzuje enzym citrátsyntáza. Acetylový zbytek navázaný na CoA a čtyřuhlíkatý oxalacetát dávají vzniknout šestiuhlíkatému citroyl-CoA. Hydrolýza vazby mezi koenzymem A a citrátem uvolní značné množství energie jako teplo.
2. citrát ↔ cis-akonitát ↔ isocitrát
Citrát je přeměňován na isocitrát, reakce je katalyzována enzymem akonitázou. Akonitáza obsahuje nehemové železo ve stavu Fe2+ a síru.[2] Citrát je nejprve dehydratován na cis-akonitát a pak rehydratován na isocitrát. Reakce je inhibována fluoracetátem, který kondenzuje jako fluoracetyl-CoA s oxalacetátem za vzniku fluorcitrátu, který působí na akonitázu inhibičně.
3. isocitrát + NAD+ ↔ oxalsukcinát ↔ α-ketoglutarát + CO2 + NADH+H+
Dehydrogenace isocitrátu za vzniku oxalsukcinátu je katalyzována isocitrátdehydrogenázou. Jsou známy tři různé isocitrátdehydrogenázy. NAD+ specifická dehydrogenasa je přítomna v mitochodriích, ostatní dvě, které jsou NADP+ specifické, se vyskytují v mitochondriích i v cytosolu. Oxidace isocitrátu pro dýchací řetězec je katalyzována výhradně NAD+ specifickou dehydrogenázou.[3] Oxalsukcinát je následně dekarboxylován na α-ketoglutarát. Tuto reakci také katalyzuje isocitrátdehydrogenáza. Důležitými složkami dekarboxylační reakce je Mn2+ nebo Mg2+.[3] Koenzym NAD+ se při reakci redukuje na NADH+H+. Protony nesené koenzymem jsou použity v dýchacím řetězci k syntéze 3 ATP.
4. α-ketoglutarát + NAD+ + CoA-SH → sukcinyl-CoA + CO2 + NADH+H+
V dalším kroku dochází k dekarboxylaci α-ketoglutarátu. Enzymy katalyzující reakci se sdružují do α-ketoglutarátdehydrogenázového komplexu. K reakci jsou potřebné kofaktory thiamindifosfát, kyselina lipoová, NAD+, FAD a koenzym A. Reakce je inhibována arsenitanem.
5. sukcinyl-CoA + Pi + GDP ↔ sukcinát + GTP + CoA-SH
Enzym sukcinátthiokinasa (sukcinyl-CoA-synthetasa) katalyzuje přeměnu sukcinyl-CoA na sukcinát. Uvolněná energie je použita k vytvoření makroergní vazby mezi fosfátem a GDP za vzniku jedné molekuly GTP. Díky GTP nakonec vzniká ATP. Reakci, při které vznikne ATP mimo dýchací řetězec, se říká substrátová fosforylace.
6. sukcinát + FAD ↔ fumarát + FADH2
Sukcinát je dehydrogenován na fumarát. Enzym sukcinátdehydrogenáza, který katalyzuje tuto reakci, je vázán na vnitřní povrch vnitřní membrány mitochondrie. Je to jediný enzym citrátového cyklu, který přenáší protony na flavoprotein, bez účasti NAD+. Malonát a oxalacetát kompetitivně inhibují sukcinádehydrogenázu. Redukovaný koenzym FADH2 přenáší redukční ekvivalenty do dýchacího řetězce. Na rozdíl od NADH ale vytvoří jen 2 molekuly ATP.
7. fumarát + H2O ↔ malát
Adicí vody na fumarát vznikne malát. Reakce je katalyzována enzymem fumarázou (fumaráthydratasa). Je specifická pro isomer malátu.
8. malát + NAD+ ↔ oxalacetát + NADH+H+
V poslední reakci je malát dehydrogenován enzymem malátdehydrogenázou zpět na oxalacetát.
Výtěžek
Během jedné otáčky vznikne při substrátové fosforylaci 1 molekula ATP. Při oxidaci acetyl-CoA se při každé otáčce cyklu vytvoří 3 molekuly NADH+H+ a jedna molekula FADH2. Při oxidativní fosforylaci v dýchacím řetězci se při oxidaci NADH+H+ na NAD+ vytváří 3 molekuly ATP, při oxidaci FADH2 se tvoří 2 molekuly ATP. Oxidací jedné molekuly acetyl-CoA by tedy mělo vzniknout 12 molekul ATP. Ve skutečnosti se ale vytvoří asi 10 ATP.[4][a]
Regulace
Citrátsynthasa, isocitrátdehydrogenasa a α-ketoglutarátdehydrogenásový komplex jsou enzymy, které jsou alostericky inhibované vysokou koncentrací ATP. To zabraňuje nadměrné spotřebě acetyl-CoA.
Některé enzymy jsou inhibovány, pokud je v mitochondrii velké množství redukovaných koenzymů. Rychlost zpětné oxidace koenzymů je závislá na dýchacím řetězci, tedy i na dostupnosti kyslíku. Nepřítomnost nebo částečný nedostatek O2 způsobí úplnou nebo částečnou inhibici cyklu. Naopak zvýšená rychlost respirace, ke které dochází při zvýšené potřebě ATP, zvyšuje i aktivitu citrátového cyklu.
Úloha v metabolismu
Citrátový cyklus nemusí proběhnout celý, některé jeho meziprodukty mohou být substrátem pro jiné metabolické dráhy, naopak jiné dráhy končí v některé součásti cyklu. Citrátový cyklus plní funkci v oxidativních i syntetických pochodech, je tzv. amfibolický. Oxalacetát může být přeměněn na pyruvát a použit ke glukoneogenezi, tzn. pro syntézu sacharidů. Cyklus může sloužit také jako zdroj uhlíkových koster k syntéze postradatelných aminokyselin. Naopak po transaminaci a deaminaci mohou aminokyseliny do cyklu vstupovat:
Glycin, alanin, cystein, hydroxyprolin, serin, threonin a tryptofan tvoří pyruvát, ze kterého je syntetizován acetyl-CoA; arginin, histidin, glutamin a prolin jsou substrátem pro tvorbu α-ketoglutarátu, isoleucin, methionin a valin tvoří sukcinyl-CoA, tyrosin a fenylalanin tvoří fumarát.
Acetyl-CoA je hlavním substrátem pro syntézu mastných kyselin.
Význam vitamínů
Čtyři vitamíny, patřící do B-komplexu, mají nezastupitelnou úlohu v citrátovém cyklu.
- Thiamin (B1) je ve své aktivní formě, thiamindifosfátu, koenzymem α-ketoglutarátdehydrogenázového komplexu.
- Riboflavin (B2) je ve formě FAD (nadenindinukleotidu) kofaktor α-ketoglutarátdehydrogenázového komplexu a sukcinátdehydrogenasy.
- Niacin (B3) je ve své aktivní formě (NAD) koenzymem dehydrogenáz cyklu
- Kyselina pantothenová (B5) je součást koenzymu A (CoA)
Odkazy
Poznámky
1 Důvodem by mohlo být to, že na jednu molekulu ATP musí protonový gradient poskytnout celkem čtyři protony (3 protony na syntézu ATP a 1 proton při symportu Pi− a H+).
Reference
- Richard L. Veech. Tricarboxylic Acid Cycle. In: Lennarz,W.J., Lane, M.D. Encyclopedia of Biological Chemistry , Four-Volume Set, 1-4. [s.l.]: [s.n.]
- metallo.scripps.edu [online]. [cit. 21-06-2010]. Dostupné v archivu pořízeném dne 08-06-2011.
- Robert K. Murray; Daryl K. Granner; Joe C. Davis; Peter A. Mayes; Victor W. Rodwell. Harper’s Illustrated Biochemistry; twenty-sixth edition. [s.l.]: [s.n.], 2003. ISBN 0-07-138901-6.
- MURRAY, Robert K., et al. Harperova biochemie. Z angl. 23. vyd. přel. Lenka Fialová et. al. 4. vyd. v ČR. Praha: H & H, 2002. ix, 872 s. ISBN 80-7319-013-3.
Literatura
- MURRAY, Robert K., et al. Harperova biochemie. Z angl. 23. vyd. přel. Lenka Fialová et. al. 4. vyd. v ČR. Praha: H & H, 2002. ix, 872 s. ISBN 80-7319-013-3.
Související články
- Beta-oxidace – přeměňuje mastné kyseliny z lipidů na acetyl-CoA
- Glykolýza – štěpí sacharidy na pyruvát, který se dekarboxylací a přidáním HS-CoA změní na acetyl-CoA
- Dýchací řetězec
Externí odkazy
 Obrázky, zvuky či videa k tématu citrátový cyklus na Wikimedia Commons
Obrázky, zvuky či videa k tématu citrátový cyklus na Wikimedia Commons