Organela
Organely jsou drobné mikroskopické útvary uvnitř buněk, se specifickou funkcí, které jsou funkční obdobou orgánů u živočichů. Nejde však o skutečné orgány, ty jsou totiž tvořeny tkáněmi (u živočichů) nebo pletivy (u rostlin), složenými z jednotlivých buněk. Buňka je rozčleněna (kompartmentována) na mnoho takových organel, které mají vždy svou specifickou úlohu. Organely jsou dle převažující definice takové buněčné struktury, které jsou obalené vlastní membránou.

Organely jsou dle takové definice převážně výsadou tzv. eukaryotických buněk, což jsou buňky tvořící tělo živočichů, rostlin, hub a prvoků. Naopak bakterie a archebakterie obvykle membránové útvary uvnitř svých buněk nemají (tzv. mesozomy jsou zřejmě artefakty vzniklé při přípravě buněk na elektronovou mikroskopii,[1] určitými výjimkami mohou být neobvyklé karboxyzomy,[2] magnetozomy[3] a jádra připomínající útvar planktomycet[4]).
Definice
Prvním vědcem, který použil slovo „organela“ (zdrobnělina ze slova orgán, jakoby „malý orgán“) pro označení buněčných struktur, byl pravděpodobně německý zoolog Karl August Möbius[5][6][7] (používá výraz „organula“ jako množné číslo od lat. „organulum“).[8] Z kontextu je zřejmé, že tento výraz používá pro rozmnožovací útvary v buňkách jednobuněčných prvoků (aby tak zdůraznil rozdíl mezi orgány mnohobuněčných a organelami jednobuněčných). Trvalo několik let, než se termín organulum uchytil pro subcelulární (podbuněčné, menší než buňka) struktury i v buňkách mnohobuněčných organismů, například v lidských buňkách. Knihy napsané kolem roku 1900 stále používají označení „buněčné orgány“.[9][10][11] V následujících letech se postupně organela stávala známějším a známějším označením[12] a kolem roku 1920 už takto byly označovány struktury umožňující pohyb (např. bičík) a další struktury na buňkách např. nálevníků.[13][14] V této době použil Alfred Kühn označení „organela“ také pro centriolu.[15]
Současná definice organely vykrystalizovala až poměrně pozdě,[16][17][18][19] načež byly za organely považovány ty buněčné struktury, které jsou obklopeny membránou. Některé práce však stále používají starší definici, podle níž je organela jakákoliv funkční jednotka uvnitř buněk.[20][21]
Organely s DNA
Organely s DNA neboli semiautonomní organely v širším slova smyslu jsou obklopeny dvěma nebo více membránami. V současné době se obecně uznává jejich vznik procesem endosymbiózy a existují pozorování nově vznikajících semiautonomních organel, například nový vznik chloroplastů u prvoka Paulinella chromatophora.[22]
Jádro
Jádro (karyon, nucleus) je řídící organela. Je kryto dvouvrstvou jadernou membránou, s póry tvořenými speciálními bílkovinami. Mají za úkol usnadnit transport makromolekul (zejména RNA). V jádře je uložen chromatin (DNA) – v době dělení buňky se organizuje do formy chromozomů, dále jadérko, ribozomy a karyolymfa. Jeho funkcí je tedy uchování genetické informace a na jejím základě řízení funkcí buňky. Jádro lze najít pouze u eukaryot. Lze v něm najít jadérko, jehož funkcí je tvorba rRNA (ribozomální RNA) a účast na regulaci buněčného dělení.
Semiautonomní organely

2 – externí membrána,
3 – mezimembránový prostor,
4 – matrix
Semiautonomní organely v užším smyslu jsou buněčné kompartmenty s vlastní genetickou informací, a to ve formě DNA prokaryotního uspořádání (DNA v kruhovém uspořádání a v plazmidech). Na základě endosymbiotické teorie se v současnosti má za to, že tyto organely jsou přímými potomky bakterií (mitochondrie) a sinic (chloroplasty a ostatní plastidy), které prošly intenzivní symbiózou s hostitelskou buňkou (pravděpodobně archea) a staly se na buňce plně závislou strukturou.
Semiautonomními jsou nazývány, protože jejich existence již není možná mimo prostředí buňky, s jejímiž signálními drahami interagují a jimiž jsou usměrňovány, ale zachovaly si ještě velkou část původní genetické informace, jejíž realizace je pro existenci a funkci této organely stále nezbytná. Semiautonomní organely nalezneme pouze u eukaryot a jejich získání představuje základní krok v evoluci od prokaryot k eukaryotám.
Pro každou semiautonomní organelu je také typické, že je obklopena dvojitou membránou. Každá membrána je tvořena tzv lipidovou dvouvrstvou, jde tedy o dvojici takovýchto již „dvojitých“ membrán (předpokládá se, že vnitřní membrána z dvojice představuje původní cytoplazmatickou membránu pohlcené bakterie a vnější membrána představuje původní cytoplazmatickou membránu hostitelské buňky, která obklopila pronikající bakterii při endocytóze – průniku bakterie dovnitř host. buňky). Obě membrány mají částečně odlišné vlastnosti, ať v již zastoupení membránových proteinů, zastoupení odlišných lipidů a často také odlišnou hodnotou gradientu redoxního potenciálu.
- Mitochondrie je zdrojem velké většiny buněčného ATP a představuje tak základní zdroj energie eukaryotické buňky. V mitochondriích probíhá oxidativní fosforylace – velmi efektivní způsob získávání energie štěpením cukrů až na oxid uhličitý a vodu. Vnitřní membrána je místem vlastní oxidace a je všelijak zprohýbaná v kristy a tvoří tak oddělené prostory (kompartmenty) pro různé chemické reakce a zároveň tím zvětšuje svou reakční plochu. Vnitřní prostor se nazývá matrix.
Mitochondrie jsou přítomné v buňkách většiny eukaryotních organismů.
Stejnou funkci jako mitochondrie plní u některých eukaryot hydrogenozomy, mající i obdobné strukturní součásti. Předpokládá se, že se vyvinuly z mitochondrií.
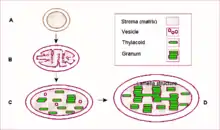
A–B – proplastid
C–D – chloroplast
- Plastidy, stejně jako mitochondrie mají dvojvrstvou membránu a vlastní DNA. Předpokládá se, že pravděpodobně vznikly z endosymbiotických sinic.[23] Sinice jsou eubakterie schopné využít energie světla v procesu fotosyntézy, symbiózou a pozdějším přisvojením si sinic tak eukaryotní buňky mohly získat schopnost vlastní fotosyntézy a mohly se orientovat na zásadně odlišnou strategii získávání energie a následně i odlišnou životní strategii.
Plastidy se vyskytují u organismů náležejících do skupiny Archaeplastida (tedy rostliny v širším slova smyslu), která obsahuje tři linie fotosyntetizujících eukaryot: glaukofyty, červené řasy, zelené řasy a ze zelených řas vyvinuvší se rostliny (Plantae sensu stricto). Na základě bioinformatického rozboru plastidové DNA se usuzuje, že plastidy obsažené v jakémkoliv z eukaryotních organismů (mimo rostlin jsou plastidy přítomné také v buňkách všech typů řas) jsou jednoho původu.[23] Mimo výše zmíněných linií však existují ještě některé řasy, jejichž plastidy vznikly sekundární endosymbiózou (Rhizaria, Excavata). Hostitelská buňka v tomto případě pohltila jednobuněčnou řasu již i s plastidem,[24] tyto plastidy pak mohou mít tři i čtyři membrány.
U zelených řas se v buňce vyskytuje jen jediný plastid, u vyšších rostlin počet plastidů na buňku není vázaný, dělí se nezávisle na buněčném dělení a bývá jich v jediné buňce mnoho. Plastidy prodělávají po rozdělení různý vývoj a můžeme je proto dále dělit na:- Chloroplasty – Jsou plastidy s plně vyvinutým fotosyntetickým aparátem, představují jedinou strukturu eukaryotní buňky, v níž může fotosyntéza probíhat, jsou zelené, dávají barvu celé rostlině.
- Proplastidy – dosud nedozrálé plastidy
- Chromoplasty – Obsahují nechlorofylová barviva, mají různé barvy, obvykle ochrannou funkci a také zabarvují rostlinu (např. podzimní listí).
- Leukoplasty – Mají zásobní funkci, obsahují především škrob.
Další membránové útvary
Jako membránové útvary se označují další buněčné kompartmenty, které jsou ohraničené a od svého okolí oddělené buněčnou membránou, a sama vnější cytoplazmatická membrána buňky. Na rozdíl od semiautonomních organel, které jsou vždy obklopeny dvojicí membrán, tyto kompartmenty mají vždy jen jednu membránovou vrstvu. Komplikovanější vnitřní membránové struktury jsou charakteristické pro evolučně rozvinutější eukaryotické buňky, prokaryota má svou vnitřní strukturu prostou, membránami dále již nečleněnou.
- Cytoplazmatická membrána (též plazmalema nebo plasmalema; plazmatická membrána, zkr. PM) je semipermeabilní (polopropustná) membrána uzavírající obsah buňky. Zprostředkovává kontakt buňky s okolím (ať už aktivním transportem, či osmózou) a zároveň ji před okolím chrání. Je tvořena lipidovou dvouvrstvou, do které jsou zabudovány různé bílkoviny. Je nezbytnou součástí buněk všech typů.
- Endoplazmatické retikulum (ER) je složitá membránová struktura obvykle v blízkosti jádra. Lze rozlišit dva typy – hladké endoplazmatické retikulum (bez přisedlých ribozómů), jehož funkcí je syntéza tuků a glykogenu , a hrubé endoplazmatické retikulum (s přisedlými ribozómy), jehož funkcí je syntéza bílkovin. Nachází se pouze u eukaryot.
- Golgiho aparát (GA) je složitá membránová struktura, jejíž funkcí je shromažďovat a dále zpracovávat produkty endoplazmatického retikula, se kterým sousedí a probíhá mezi nimi váčkový – vezikulární transport. Nachází se pouze u eukaryot.
- Lysozomy jsou typické pro živočišné buňky, obsahují proteolytické a hydrolytické enzymy, slouží k degradaci látek buňkou fagocytovaných nebo do buňky proniklých pinocytózou.
- Vakuolu nalezneme především u rostlin, plní zde funkci zásobního orgánu, obvykle u dospělých buněk vyplňuje velkou většinu buněčného obsahu. Rovněž některá protista mají vyvinutou vakuolu, která může plnit mnoho funkcí (potravní, pulsující…).
- Thylakoidy jsou jednoduché membránové váčky v prokaryotických buňkách a semiautonomních organelách.
- Mikrotělíska (peroxizómy a glyoxizómy) mají na starosti přeměnu aminokyselin a tuků na cukry.
Odkazy
Reference
V tomto článku byl použit překlad textu z článku Organelle na anglické Wikipedii.
- Ryter A. Contribution of new cryomethods to a better knowledge of bacterial anatomy. Ann. Inst. Pasteur Microbiol.. 1988, s. 33–44. DOI 10.1016/0769-2609(88)90095-6. PMID 3289587. (anglicky)
- Kerfeld CA, Sawaya MR, Tanaka S, Nguyen CV, Phillips M, Beeby M, Yeates TO. Protein structures forming the shell of primitive bacterial organelles. Science. 2005, s. 936–8. DOI 10.1126/science.1113397. PMID 16081736. Bibcode 2005Sci...309..936K. (anglicky)
- Komeili A, Li Z, Newman DK, Jensen GJ. Magnetosomes are cell membrane invaginations organized by the actin-like protein MamK. Science. 2006, s. 242–5. DOI 10.1126/science.1123231. PMID 16373532. Bibcode 2006Sci...311..242K. (anglicky)
- Fuerst JA. Intracellular compartmentation in planctomycetes. Annu. Rev. Microbiol.. 2005, s. 299–328. DOI 10.1146/annurev.micro.59.030804.121258. PMID 15910279. (anglicky)
- BÜTSCHLI, O. Dr. H. G. Bronn's Klassen u. Ordnungen des Thier-Reichs wissenschaftlich dargestellt in Wort und Bild. Erster Band. Protozoa. Dritte Abtheilung: Infusoria und System der Radiolaria.. [s.l.]: [s.n.], 1888. S. 1412. (anglicky)
- Amer. Naturalist. 23, 1889, S. 183: „It may possibly be of advantage to use the word organula here instead of organ, following a suggestion by Möbius. Functionally differentiated multicellular aggregates in multicellular forms or metazoa are in this sense organs, while, for functionally differentiated portions of unicellular organisms or for such differentiated portions of the unicellular germ-elements of metazoa, the diminutive organula is appropriate.“ Cited after : Oxford English Dictionary online, entry for „organelle“.
- 'Journal de l'anatomie et de la physiologie normales et pathologiques de l'homme et des animaux' at Google Books
- MÖBIUS, K. Das Sterben der einzelligen und der vielzelligen Tiere. Vergleichend betrachtet. Biologisches Centralblatt. 1884, s. 389–392, 448. Dostupné v archivu pořízeném dne 2011-07-18. (anglicky)
- HÄCKER, Valentin. Zellen- und Befruchtungslehre. Jena: Verlag von Gustav Fisher, 1899. (anglicky)
- WILSON, Edmund B. The cell in Development and Inheritance. second. vyd. New York: The Macmillan Company, 1900. (anglicky)
- HERTWIG, Oscar. Allgemeine Biologie. Zweite Auflage des Lehrbuchs "Die Zelle und die Gewebe". Jena: Verlag von Gustav Fischer, 1906. (anglicky)
- LIDFORSS, B. Allgemeine Biologie. Redakce Paul Hinneberg. Leipzig, Berlin: Verlag von B.G.Teubner, 1915. Kapitola Protoplasma, s. 227 (218–264). (anglicky)
- Kofoid CA, Swezy O. Flagellate Affinities of Trichonympha. Proc. Natl. Acad. Sci. U.S.A.. 1919, s. 9–16. DOI 10.1073/pnas.5.1.9. PMID 16576345. Bibcode 1919PNAS....5....9K. (anglicky)
- Cl. Hamburger, Handwörterbuch der Naturw. Bd. V, .S. 435. Infusorien. cited after PETERSEN, Hans. Über den Begriff des Lebens und die Stufen der biologischen Begriffsbildung. Archiv für Entwicklungsmechanik der Organismen (now: Development Genes and Evolution). 1919, s. 423–442. ISSN 1432-041X. DOI 10.1007/BF02554406. (anglicky)
- KÜHN, Alfred. Untersuchungen zur kausalen Analyse der Zellteilung. I. Teil: Zur Morphologie und Physiologie der Kernteilung von Vahlkampfia bistadialis. Archiv für Entwicklungsmechanik der Organismen (now: Development Genes and Evolution). 1920, s. 259–327. DOI 10.1007/BF02554424. (anglicky)
- Nultsch, Allgemeine Botanik, 11. Aufl. 2001, Thieme Verlag
- Wehner/Gehring, Zoologies, 23. Aufl. 1995, Thieme Verlag
- Alberts, Bruce et al. (2002). The Molecular Biology of the Cell, 4th ed., Garland Science, 2002, ISBN 0-8153-3218-1. online via "NCBI-Bookshelf"
- Brock, Mikrobiologie, 2. korrigierter Nachdruck (2003), der 1. Aufl. von 2001
- Strasburgers Lehrbuch der Botanik für Hochschulen, 35. Aufl. (2002), S. 42
- Alliegro MC, Alliegro MA, Palazzo RE. Centrosome-associated RNA in surf clam oocytes. Proc. Nat. Acad. Sci. USA. 2006, s. 9037–9038. DOI 10.1073/pnas.0602859103. PMID 16754862. Bibcode 2006PNAS..103.9034A. (anglicky)
- KEELING, PJ.; ARCHIBALD, JM. Organelle evolution: what's in a name?. Curr Biol. Apr 2008, roč. 18, čís. 8, s. R345-7. DOI 10.1016/j.cub.2008.02.065. PMID 18430636.
- Isao Inouye and Noriko Okamoto: “Changing concepts of a plant: current knowledge on plant diversity and evolution”. Plant Biotechnology, Vol. 22, pp. 505–514 (2005)
- S. E. Douglas, Curr. Opin. Genet. Dev. 8, 655 (1998).
Literatura
Externí odkazy
 Slovníkové heslo organela ve Wikislovníku
Slovníkové heslo organela ve Wikislovníku Obrázky, zvuky či videa k tématu organela na Wikimedia Commons
Obrázky, zvuky či videa k tématu organela na Wikimedia Commons