Försterův rezonanční přenos energie
Försterův rezonanční přenos energie (FRET) či nesprávně Fluorescenční rezonanční přenos energie je mechanismus nezářivého přenosu energie mezi dvěma molekulami prostřednictvím dipól-dipólové interakce. Molekula, z níž se energie přenáší, se nazývá donor (dárce), kdežto molekula, která energii přijímá, se nazývá akceptor (příjemce).
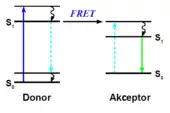
Účinnost tohoto mechanismu je nepřímo úměrná šesté mocnině vzdálenosti mezi donorem a akceptorem, k jevu tedy prakticky dochází jen tehdy, jsou-li obě molekuly v těsné blízkosti. Z intenzity FRET (kterou lze určit různými metodami analýzy fluorescence vzorku) lze proto získat informaci o míře interakce mezi donorem a akceptorem. To je velmi cenné především při studiu živých buněk.[1]
Historie
Jedny z prvních experimentálních důkazu existence FRET získali roku 1922 G. Cairo a J. Franck. Pozorovali, že ozařují-li páry rtuti a thallia světlem o vlnové délce spadající do oblasti absorpce rtuti, dochází částečně ke fluorescenci thallia. Tento jev nazvali „senzibilizovaná fluorescence“.[2] Na objasňování principu tohoto jevu se podíleli např. Jean-Baptiste Perrin,[3] Francis Perrin,[4] Hartmut Kallman či Fritz London.[5] Se správným modelem však přišel až v roce 1948 Theodor Förster, po němž je dnes jev pojmenován.[6]
Fyzikální popis
Odvození
FRET je čistě elektromagnetickým jevem spočívajícím v nezářivém přenosu energie mezi dvěma dipóly (donorem a akceptorem) na vzdálenost podstatně menší, než je vlnová délka záření emitovaného kmitajícím donorem. K odvození správného vztahu pro FRET lze použít klasický popis.[7] Energie Coulombické interakce mezi nabitými tělesy a s nábojovou hustotou a , vzdálenými od sebe , je





|
(1) |

Provedeme-li multipólový rozvoj uvedeného potenciálu, dostaneme
|
(2) |

kde .

Pro nenabité částice je nenulový pouze poslední ze členů v závorce, který odpovídá dipól-dipólové interakci. Právě tento člen je zodpovědný za existenci FRET, ale také např. za van der Waalsovské interakce. Ostatní členy pro jednoduchost zanedbáme, budeme tedy uvažovat, že akceptor je dipól indukovaný polem dipólu donoru.
Intenzitu FRET lze vyjádřit jako
|
(3) |

kde poměr rychlostní konstanty přenosu energie z donoru na akceptor a rychlostní konstanty deexcitace donoru prostřednictvím jiných mechanismů (obě uvedené rychlostní konstanty mají hodnotu převrácených hodnoty příslušných dob života) je vyjádřen jako podíl energie absorbované akceptorem za jednotku času a energie vyzářené za jednotku času donorem v nepřítomnosti akceptoru . Hodnotu lze určit pomocí vztahu pro energii vyzařovanou dipólem kmitajícím s frekvencí :





|
(4) |

Veličinu na základě Poyntingoy věty vyjádříme jako
|
(5) |

kde je elektrické pole generované donorem a je proudová hustota asociovaná s přítomností akceptorového dipólu. Tu lze vyjádřit jako


|
(6) |

čímž se (5) zjednoduší na
|
(7) |

Jelikož je dipól indukovaný polem donoru, můžeme ho vyjádřit prostřednictvím tenzoru polarizovatelnosti akceptoru :


|
(8) |

Za předpokladu, že akceptor lze polarizovat pouze ve směru vektoru jeho dipólového momentu (tedy že lze psát ), rovnice (7) přejde na


|
(9) |

Po relativně složitém odvození (zahrnujícím vyjádření pomocí Greenovy funkce, středování vzájemné orientace donoru a akceptoru a další kroky) získáme důležitý vztah pro závislost FRET na vzdálenosti obou zúčastněných dipólů:

|
(10) |

přičemž je tzv. Försterova vzdálenost. Ze vztahu vyplývá, že intenzita FRET se vzdáleností silně klesá, přičemž intenzitě ostatních deexcitačních procesů se vyrovná, je-li vzdálenost donoru a akceptoru právě . Typické hodnoty jsou 1-10 nm.

Försterovu vzdálenost lze vyjádřit jako
|
(11) |

Zde je emisní spektrum donoru, je index lomu prostředí. Veličina představuje účinný průřez akceptoru pro danou frekvenci , který vyjadřuje schopnost akceptoru na dané frekvenci absorbovat:




|
(12) |

FRET tedy závisí na překryvu emisního spektra donoru a absorpčního spektra akceptoru.
Faktor , který v (11) rovněž figuruje, vyjadřuje závislost FRET na vzájemné orientaci donoru a akceptoru:

|
(13) |

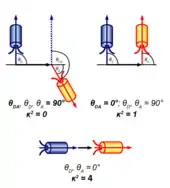
Vektor , resp. je jednotkový vektor ve směru dipólového momentu akceptoru, resp. donoru a je jednotkový vektor ve směru spojnice donoru a akceptoru.



Výraz může nabývat hodnot mezi 0 a 4. Minimální hodnotu 0 má v případě, že oba dipólové momenty jsou na sebe kolmé. Hodnotu 1 má v případě, že jsou rovnoběžné a zároveň kolmé na . Maximální hodnotu 4 má v případě, že oba dipóly leží v jedné přímce.
Stojí za povšimnutí, že (a tedy i intenzita FRET) má nejvyšší hodnotu, leží-li akceptor ve směru osy dipólu donoru. Hodnota je zde nenulová, na základě výrazu (5) tedy k přenosu energie může dojít, a to přesto, že v aproximaci dalekého pole se tímto směrem nešíří žádné elektromagnetické vlny. Proto se jedná o „nezářivý přenos“.

V případě, kdy se donor i akceptor v roztoku průběžně náhodně reorientují, vystředuje se na hodnotu . Jsou-li naopak donor a akceptor částečně či zcela fixovány (na stejné makromolekule či v membráně), může se od hodnoty odchylovat. FRET pak může poskytnout informaci o vzájemné orientaci donoru a akceptoru.

Účinnost FRET

FRET lze alternativně popsat prostřednictvím účinnosti, která se spočítá jako kvantový výtěžek příslušného přechodu, tedy podíl uskutečněných přenosů vzhledem k počtu excitací. Lze ji též vyjádřit pomocí rychlostních konstanty FRET a rychlostní konstanty reprezentující ostatní deexcitační jevy:
|
(14) |

To lze po dosazení z (10) upravit na
|
(15) |

Pomocí dob života lze účinnost FRET vyjádřit jako
|
(16) |

kde a jsou doby života excitovaných stavů donoru v přítomnosti, resp. absenci akceptoru.


Aby byla účinnost FRET nenulová, musí být splněny 3 základní podmínky:
- Překryv spekter: donor musí fluoreskovat a jeho emisní spektrum se musí alespoň částečně překrývat s absorpčním spektrem akceptoru. Akceptor naopak nemusí být fluoroforem.
- Vzdálenost: donor a akceptor od sebe nesmí být dál než několik nanometrů.
- Orientace: donor a akceptor vůči sobě nesmí být natočené tak, aby orientační faktor byl nulový
Kvantový přístup
V rámci kvantové teorie pole lze interakce mezi částicemi obecně popsat s pomocí Feynmanových diagramů jako výměnu tzv. virtuálních částic. FRET lze proto popsat jako výměnu virtuálních fotonů mezi donorem a akceptorem.
Aplikace
V biofyzice a biochemii FRET představuje užitečný nástroj umožňující in vivo kvantifikaci buněčných dějů jako jsou interakce molekul či jejich konformační změny (je ovšem třeba mít k dispozici metodu, kterou lze na tyto molekuly připevnit vhodný donor, resp. akceptor). Lze tak však získat pouze informaci o průměrných vzdálenostech donoru a akceptoru (s výjimkou metody smFRET).
Konkrétně se FRET uplatňuje např. v následujících oblastech:
- studium struktury a konformace proteinů[8]
- metoda imunoassay[9]
- studium struktury a konformace nukleových kyselin[10]
- real-time PCR[11]
- studium distribuce a transportu lipidů[12]
- studium fúze membrán[13]
- v přírodě se FRET uplatňuje při svádění světelné energie ze světlosběrné antény do reakčního centra fotosystému[14][15]
Metody měření FRET
Senzitizovaná emise
Jelikož FRET vede ke snížení intenzity fluorescence donoru, lze jej z měření tohoto poklesu teoreticky určit. Je-li , resp. intenzita fluorescence donoru v nepřítomnosti, resp. přítomnosti akceptoru, pak účinnost FRET lze vyjádřit jako


|
(17) |

V praxi však vždy představuje komplikaci tzv. crosstalk,[16] neboli skutečnost, že excitační, resp. emisní spektra donoru a akceptoru se alespoň částečně překrývají. V případě překryvu excitačních spekter pak dochází k nežádoucí přímé excitaci akceptoru, který následně může fluoreskovat i v případě, kdy k FRET nedochází.
V případě překryvu emisních spekter fluorescence donoru a akceptoru částečně splývají. To do měření rovněž vnáší nepřesnost, neboť pokles fluorescence donoru v důsledku FRET se pak jeví menší než ve skutečnosti je.
Proto je nutné zároveň se vzorkem obsahujícím donor i akceptor měřit i kontroly obsahující pouze donor, resp. pouze akceptor, a tyto kontroly od samotného vzorku odečíst. To však ve výsledném obraze zvyšuje úroveň šumu. Pro měření malých signálů proto metoda senzitizované emise není příliš vhodná.
Vybělování akceptoru
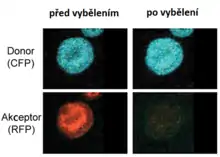
Fluoreskuje-li kromě donoru též akceptor, lze k určení intenzity FRET též využít jeho vybělování. Měření sestává z následujících kroků:
- Měření fluorescence donoru ve vzorku s donorem i akceptorem
- Vybělení akceptoru
- Měření fluorescence donoru ve vzorku s donorem a vyběleným akceptorem
V prvním kroku je měřená fluorescence donoru umenšovaná přenosem energie na akceptor prostřednictvím FRET. Po vybělení akceptoru je FRET znemožněn, z rozdílu signálu lze tedy velikost FRET určit.
Výhodou této metody je oproti senzitizované emisi to, že stačí měřit jediný vzorek. Nevýhodou je, že vzorek lze změřit pouze jednou, neboť vybělení akceptoru je nevratné. Ve vzorku tedy nelze sledovat dynamiku procesů[16].
Měření dob života fluorescence
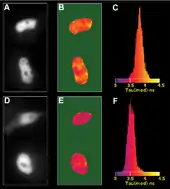
Všechny fluorofory vykazují (multi)exponenciální dohasínání, typicky na nanosekundových škálách. Rychlost dohasínání přitom závisí na řadě faktorů a jevů (včetně FRET), které fluorescenci zháší. Právě toho metoda měření dob života využívá. Postup je podobný jako u metody vybělování akceptoru: napřed se provede měření doby života fluorescence donoru na vzorku s donorem i akceptorem, posléze je akceptor vybělen a vzorek je změřen znovu. Intenzita FRET je potom stanovena z rozdílu v obou vzorcích.
Měření dob života představuje poměrně precizní metodu určování FRET. Vzhledem k tomu, že je sledována pouze fluorescence donoru, může jako akceptor sloužit i nefluorescenční molekula, díky čemuž nedochází k přeslechům v emisních spektrech.
Mezi nevýhody patří, že dobu života fluorescence donoru ovlivňují kromě přítomnosti akceptoru i další faktory prostředí, jako jsou např. změny pH. Aparatury schopné měření časově rozlišené fluorescence na nanosekundových škálách se též vyznačují vyšší cenou.
Měření dob života fluorescence se využívá při technice Fluorescence Lifetime Imaging Microscopy (FLIM). Obrazy vzorku pořízené touto metodou odráží rozdíly v dobách života fluorescence v jeho různých místech.
Měření polarizace (Homo-FRET)
Zkratka “FRET” obvykle označuje techniky, při nichž se používá akceptor jiného typu (chemického složení), než je donor. V mnoha situacích je však zapotřebí studovat interakce mezi proteiny stejného typu, nebo dokonce interakci různých částí stejné proteinové molekuly (skládání neboli folding), které jsou značeny stejným fluoroforem (odtud Homo-FRET).
V takovémto případě, kdy donor zároveň plní roli akceptoru (to je možné za předpokladu, že se jeho excitační a emisní spektrum částečně překrývají), nelze FRET měřit sledováním spektrálních změn, neboť emisní spektra obou zúčastněných molekul jsou shodná. Situace, kdy dochází, resp. nedochází k FRET však lze přesto rozlišit, a to měřením rozdílu v polarizaci dopadajícího a emitovaného světla. Vzorek excitujeme lineárně polarizovaným světlem. V případě, že k FRET nedochází, je depolarizace nastalé fluorescence způsobena pouze rotací vzorku. Pokud však k FRET dochází, má navíc část emitovaných fotonů polarizaci odlišnou od polarizace dopadajícího světla. To je způsobeno tím, že byly vyzářeny akceptorem, který může mít oproti donoru jinou prostorovou orientaci. Intenzitu FRET pak lze určit z hodnoty veličiny jménem anizotropie.[17]
Bioluminiscenční resonanční přenos energie (BRET)
Nevýhodou FRET je nutnost vzorek osvětlovat externím zdrojem, což může vést ke vzniku šumu způsobeného přímou excitací akceptoru nebo k vybělování. Tomuto problému se lze vyhnout s pomocí techniky zvané Bioluminiscenční Resonanční Přenos Energie neboli BRET. Tato technika místo CFP využívá bioluminiscenčních látek označovaných jako luciferiny. Tyto látky jsou s pomocí katalytického enzymu luciferázy schopny podstoupit oxidaci, jejímž produktem je molekula v excitovaném stavu. Ta se následně deexcituje emisí fotonu - nebo právě prostřednictvím FRET.
Single molecule FRET (smFRET)
Při běžném měření FRET je získáván souhrnný signál souboru donor-akceptorových párů ve vzorku, získaná hodnota FRET je tedy hodnotou průměrnou. Při smFRET jsou naopak donor-akceptorové páry sledovány jednotlivě. Tímto způsobem je možné monitorovat i drobné změny vzdálenosti a orientace jednotlivých molekul, které by jinými variantami FRET měření kvůli středování nebyly detekovatelné. Sledování jednotlivých donor-akceptorových párů je dosaženo buďto s pomocí konfokálního mikroskopu, který sbírá světlo pouze z určitého optického řezu vzorkem, nebo pomocí tzv. TIRFM mikroskopu využívajícího totální odraz. V takovém případě jsou excitovány jen donory nacházející se blízko rozhraní vzorku s podložkou.[18][19]
Molekuly využívané při měření FRET
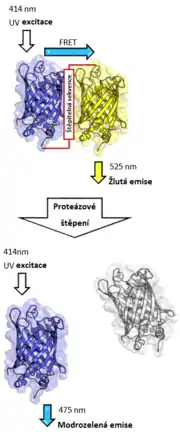
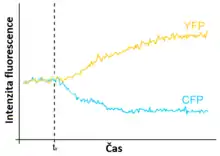
Z neproteinových látek se často používá např. pár fluorescein (donor) - rhodamin (akceptor).
Mezi proteiny je častou volbou dvojice modrozelený fluorescenční protein (CFP) - žlutý fluorescenční protein (YFP). Oba proteiny jsou odvozeny ze zeleného fluorescenčního proteinu (GFP). Použití variant GFP ke značení proteinů má oproti použití jiných molekul tu výhodu, že je lze připevnit pomocí genetického inženýrství (modifikací genu značeného proteinu), což může být jednodušší než metody používané ke značení neproteinovými molekulami.
Dalším důležitým proteinem je tzv. Cameleon vytvořený spojením proteinu BFP, kalmodulinu, EGFP a peptidu M13.[20]
Tento protein lze použít jako senzor přítomnosti vápenatých iontů. Každá jeho molekula má donorovou i akceptorovou část a v přítomnosti Ca2+ změní konformaci tak, že dochází k FRET.
Ve variantě FRET zvané BRET jsou jako donory používány látky zvané luciferiny, jejichž fluorescence je spouštěna chemickou reakcí katalyzovanou enzymem luciferázou.
Reference
- Archivovaná kopie [online]. [cit. 2014-05-27]. Dostupné v archivu pořízeném dne 2011-07-16. (Anglicky)
- CAIRO G, FRANCK J. Über Zerlegung von Wasserstoffatomen durch angeregte Quecksilberatome. Z. Physik.. Roč. 1922, čís. 11, s. 161–166.
- PERRIN J. Fluorescence et induction moleculaire par resonance. C. R. Hebd. Seances Acad. Sci.. Roč. 1927, čís. 184, s. 1097–1100.
- PERRIN F. Théorie des transferts d'activation entre molécules de méme espèce. Ann. Chim. Phys. (Paris).. Roč. 1932, čís. 17, s. 283–314.
- KALLMANN H, LONDON F. Über quantenmechanische Energieübertrangungen zwischen atomaren Systemen. Z. Physik. Chem. B. 2. Roč. 1928, s. 207–243.
- FÖRSTER T. Zwischenmolekulare Energiewanderung und Fluoreszenz. Ann. Physik.. Roč. 1948, čís. 437, s. 55. DOI 10.1002/andp.19484370105.
- NOVOTNY, L., HECHT, B. Principles of Nano-optics. [s.l.]: Cambridge University Press, 2006. [www.cambridge.org/9780521832243 Dostupné online]. ISBN 978-0-521-53988-3. Kapitola 8.6, s. 35–56. (anglicky)
- http://www.lifetechnologies.com/search/support/supportSearchAction.action?query=PN68049%20PN67514%20PN66631%20PN58377%20PN58125&supportSearchArea=Citations+%26+References&initialSearch=true&refineSearch=true&mode=or
- http://www.lifetechnologies.com/search/support/supportSearchAction.action?query=PN68132%20PN67190%20PN35384&supportSearchArea=Citations+%26+References&initialSearch=true&refineSearch=true&mode=or
- http://www.lifetechnologies.com/search/support/supportSearchAction.action?query=PN68706%20PN58125&supportSearchArea=Citations+%26+References&initialSearch=true&refineSearch=true&mode=or
- Didenko VV: DNA probes using fluorescence resonance energy transfer (FRET): designs and applications. In: BioTechniques. 31, Nr. 5, November 2001, S. 1106–16, 1118, 1120–1. PMID 11730017. PMC: 1941713 (freier Volltext)
- http://www.lifetechnologies.com/search/support/supportSearchAction.action?query=PN68376%20PN67217%20PN57759&supportSearchArea=Citations+%26+References&initialSearch=true&refineSearch=true&mode=or
- http://www.lifetechnologies.com/search/support/supportSearchAction.action?query=PN67099%20PN58732%20PN44200&supportSearchArea=Citations+%26+References&initialSearch=true&refineSearch=true&mode=or
- Laible PD et al.: Detailed Balance in Förster–Dexter Excitation Transfer and Its Application to Photosynthesis. In: J. Phys. Chem. B. 102, Nr. 9, 1998, S. 1641–1648.doi:10.1021/jp9730104
- Hochspringen↑ Kê, Bacon: Photosystem II. In: Photosynthesis: photobiochemistry and photobiophysics. Kluwer Academic, 2001, ISBN 0-7923-6791-X, S. 199-322
- Basics of FRET Microscopy: Fundamental Principles of Förster Resonance Energy Transfer (FRET) Microscopy with Fluorescent Proteins [online]. Nikon Instruments Inc., 2021 [cit. 2021-03-15]. Dostupné online. (anglicky)
- Fluorescence Anisotropy Studies [online]. HORIBA Jobin Yvon, Inc. [cit. 2021-03-15]. Dostupné online. (anglicky)
- HA T, ENDERLE T, OGLETREE DF, CHEMLA DS, SELVIN PR, WEISS S. Probing the interaction between two single molecules: fluorescence resonance energy transfer between a single donor and a single acceptor. Proc. Natl. Acad. Sci. U.S.A.. Roč. June 1996, čís. 93 (13), s. 6264–8. DOI 10.1073/pnas.93.13.6264. PMID 8692803.
- KASTANTIN, M.; SCHWARTZ, D. K. Connecting Rare DNA Conformations and Surface Dynamics Using Single-Molecule Resonance Energy Transfer. Acs Nano 2011. Čís. 5 (12), s. 9861–9869.
- MIYAWAKI A, GRIESBECK O, HEIM R, TSIEN RY. Dynamic and quantitative Ca2+ measurements using improved Cameleons. Proc Natl Acad Sci USA. Roč. 1999, čís. 96 (5), s. 2135–40. DOI 10.1073/pnas.96.5.2135. PMID 10051607.