Alphavirus
Alphavirus je rod virů, který náleží k čeledi Togaviridae. Jde o viry lidí, savců, ptáků, moskytů a vačnatců, které jsou přenášeny hmyzem. Způsobují encefalitidu, horečky a bolesti. jsou celosvětově rozšířené, avšak antivirotikum neexistuje.
 Alphavirus | |
| Baltimorova klasifikace virů | |
| Skupina | IV (ssRNA viry s pozitivní polaritou) |
| Vědecká klasifikace | |
| Realm | Riboviria |
| Říše | Orthornavirae |
| Kmen | Kitrinoviricota |
| Třída | Alsuviricetes |
| Řád | Martellivirales |
| Čeleď | Togaviridae |
| Druh | Alphavirus |
| Druh | |
| |
| Některá data mohou pocházet z datové položky. | |
Virologie
Morfologie
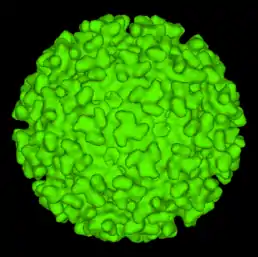
Virus se řadí mezi obalené viry (má membránovou schránku), přičemž se na membráně (o průměru 65-70 nm) vyskytuje 80 špic složených z trojic heterodimerů glykoproteinů E1 a E2, společně s menším proteinem 6K. Membrána je bohatá na cholesterol a sfingolipid, což jsou složky nezbytné pro vstup a výstup z buňky. Kapsid je sférický (ikosahedrický) o průměru 35-40 nm a skládá se z 240 podjednotek, (T=4).
Genom
Genom tvoří jedna molekula jednovláknové RNA s pozitivní polaritou o délce 11,8 (alphavirus) kb a o mol. hm. 4 x 106. Nestrukturní proteiny (replikáza) jsou umístěné na 5' konci, zaujímající zhruba 2/3 délky genomu, zatímco strukturní (kapsidový protein a 3 obalové proteiny) jsou v poslední třetině, na 3' konci. Obě tyto skupiny jsou překládány jako polyprotein, který je následně rozštěpen. Genom je očepičkován a polyadenylován. nestrukturní proteiny jsou překládány rovnou z genomu, zatímco strukturní proteiny jsou překládány z subgenomické mRNA. Na začátku genomu je průměrně 59 nukleotidů dlouhá nepřekládaná oblast, zatímco na konci je dlouhá průměrně 322 nukleotidů.
Proteom
Proteom alphavirů se seskládá z několika proteinů, které jsou translatovány v nestrukturním polyproteinu a v strukturním polyproteinu. Oba proteiny jsou schopny se samy rozštěpit a uvolnit tak funkční proteiny.
nsP1
Tento protein je součástí nestrukturního polyproteinu P123(4). Skládá se z 540 aminokyselinových zbytků a jako jediný je v asociaci s membránou, skrze palmytolilovaný cystein (420). Protein se však může vázat i hydrofobními zbytky aminokyselin 245-264. Má methyltransferázovou a guanylyltranferázovou aktivitu.
nsP2
Tento protein je součástí nestrukturního polyproteinu P123(4). Skládá se z 807 aminokyselinových zbytků. N koncová doména má helikázovou, NTP a RNA-TP aktivitu, zatímco C koncová doména má papain-like serinovou proteázovou aktivitu a nefunkční methyltransferázovou aktivitu. Obsahuje také sekvenci, díky které je transportován do jádra, což vysvětluje fakt, že 50 % celkového množství tohoto proteinu je lokalizováno právě v jádře.
nsP3
Tento protein je součástí nestrukturního polyproteinu P123(4). Skládá se z 556 aminokyselinových zbytků. Funkce tohoto proteinu zůstávají sporné, avšak ukazuje se, že má roli v RNA syntéze. Zatímco N koncová doména je konzervativní napříč alphaviry, C koncová doména, obsahující asi 200 aminokyselinových zbytků, je bohatá na serinové a threoninové zbytky, které mohou být fosforylované, avšak význam této modifikace je stále nejasný. Protein má k membránám slabou afinitu, avšak asociuje s nimi v případě, že je exprimován v nepřítomnosti dalších nsP.
nsP4
Tento protein je součástí nestrukturního polyproteinu P123(4). Skládá se z 610 aminokyselinových zbytků. Jde o hlavní replikační protein. Jeho hladina je nižší, kvůli opal stop kodonu v polyproteinu P1234 mezi nsP3 a nsP4. Ukázalo se však, že odstranění tohoto stop kodonu spíše sníží replikaci virové RNA. I když je z větší části homologní s dalšími RdRP, N koncová doména obsahuje úsek, který nemá u jiných replikáz obdoby a slouží zřejmě jako vazebná doména pro ostatní nsP. N konec však také obsahuje tyrosinové zbytky, které zodpovídají za nestabilitu tohoto proteinu a za jeho degradaci. Protein se však stává relativně stabilním v komplexu s ostatními replikačními proteiny.
Capsid
Tento protein se skládá z 264 aminokyselinových zbytků a jeho úloha je obalení RNA a vytvoření nukleokapsidu. Prostupuje membránou a jeho 33 aminokyselin tvořících C koncovou doménu má strukturu dvou řeckých klíčů a aktivitu serionové proteázy. Jde o strukturní protein.
E3
Tento protein se skládá z 64 aminokyselinových zbytků a je součástí pE2 prekurzoru, jehož součástí je i E2. Jde o strukturní protein.
E2
Tento protein se skládá z 423 aminokyselinových zbytků a je součástí pE2 prekurzoru, jehož součástí je i E3. Je zodpovědný za navázání na receptor. Jeho doména tyr-ala-leu interaguje s hydrofobní kapsou nukleokapsidu. Jde o strukturní protein.
E1
Tento protein se skládá z 439 aminokyselinových zbytků a je zodpovědný za fúzi membrán. Membránou protein prostupuje, avšak dovnitř zasahují pouze 2 aminokyselinové zbytky. Obsahuje fúzní peptid, který je však v heterodimeru s E2 blokován. Tento heterodimer se však v nízkém pH rozpadne, čímž se peptid odkryje. Ten se pak zanoří do cílené membrány (obsahující cholesterol), načež dojde k trimerizaci E1 a masivní konformační změně tohoto glykoproteinu. Trimer má pak za následek deformaci membrány a její spojení s membránou endozomu, čímž vznikne pór, kterým nukleokapsid vnikne do buňky. E1 má také zjevně vlastnosti iontového kanálu, což je důležité pro replikaci. Jde o strukturní protein.
6K
Tento protein se skládá z 55 aminokyselinových zbytků. zesiluje aktivitu jednotlivých částic. Jde o strukturní protein.
Receptory
Alphaviry vykazují neobyčejnou hostitelskou rozmanitost. Na základě pozorování byly vytvořeny 2 hypotézy, které se však nevylučují a nejspíše je pravda tvořena kombinací obou. První tvrdí, že alphaviry má E2 mnoho vazebných míst, což mu dovoluje navázat se na i vzdálenější buněčné receptory. Druhá hovoří o možnosti ubiquitinového receptoru, který je vysoce konzervovaný pro mnoho druhů.
| Název buňky | Tkáňový původ buňky | Virus | Gen a/nebo velikost | Reference |
|---|---|---|---|---|
| JY | Lidský B lymphoblastoid | SFV | HLA-A, HLA-B | Helenius et al., 1978(95) |
| CEF | Fibroblast kuřecího embrya | SINV | ?63-kDa | Ubol & Griffin, 1991 (261) |
| N18 | Myší neuroblastom | SINV | ?110 & 74-kDa | Wang et al., 1991 (277) |
| BHK-21 | Ledviny křeččího mláděte | SINV | 67-kDa high affinity | Wang et al., 1992 (278) |
| C6/36 | Moskytí larva | VEEV | Lamininový receptor | Ludwig et al., 1996 (152) |
| BHK-21 | Ledviny křeččího mláděte | SINV | 32-kDa lamininový receptor | Byrnes & Griffin, 1998 (21) |
| CHO | Vaječníky čínského křečka | SINV | Heparan sulfát | Jan et al., 1999 (103) |
| THP-1 | Lidská monocytická leukemie | SINV | heparan sulfát | Klimstra et al., 2003 (111) |
| HeLa | Lidskéý epiteliální nádor | RRV | DC-SIGN/L-SIGN Integrin ?1/ß1 | La Linn et al., 2005 (119) |
Replikace
Během životního cyklu se v buňce objevují 3 typy virové RNA, genomová +RNA, komplementární -RNA a subgenomická mRNA. Replikace je iniciována na cytosolické straně membrán endozomů a lysosomů. Nejprve je syntetizována -RNA pomocí P123 nebo P23 a nsP4, zatímco nerozštěpený P1234 je nefunkční stejně jako jednotlivé nsP. Jak probíhá syntéza -RNA, pokračuje translace nsP a tím se zvyšuje koncentrace proteázových prekurzorů. Vazby nsP1-nsP2 a nsP2-nsP3 jsou rozštěpeny, čímž se změní konformace replikázy a dojde k aktivaci syntézy +RNA. Jako u většiny +RNA virů, je poměr +RNA a −RNA syntézy asi 50-20:1. Subgenomická mRNA se syntetizuje z minimálního promotorového elementu umístěného −19 až +5 nukleotidů vůči pozici startu. Větší fragment umístěný -98 až +14 nukleotidů vůči pozici startu však vykazuje 3–6krát větší aktivitu a nahrazuje plně aktivní promotor. buňka jí obsahuje obvykle až 3× více než +RNA.
Syntéza strukturních proteinů
Pořadí strukturních proteinů na polyproteinu je kapsid-PE2(E3+E2)-6K-E1. Kapsid je odštěpen ihned po syntéze smyčky mezi ním a PE2 díky jeho C koncové doméně, která má chymotrypsin-like sbalení a má tak serin-like proteázovou aktivitu. Nový N konec polyproteinu obsahuje sekvenci, která ho navádí do endoplasmatického retikula. Další takové sekvence jsou na C koncích E2 a 6K, aby se tak do endoplasmatického retikula nasměroval i 6K a E1. E1, E2 a 6K jsou transmembránové proteiny přičemž E3 je z PE2 vystřižen. Genomová RNA obsahuje specifickou balicí sekvenci mezi nukleotidy 945 a 1076, která je rozpoznávána aminokyselinovými zbytky 81 až 113 kapsidu. Aura virus navíc do svého kapsidu balí i subgenomickou mRNA. Pro sbalení je potřeba exprimovat nsP123, není však jasné proč. Mezitím se na membránové proteiny navážou dlouhé mannosové řetězce a dále upravené podle dostupnosti upravovaného místa. Na několika místech E2 a 6K dochází k palmytaci. E1 a E2 navíc vytváří mezistavy ve svém sbalování, které vyžadují chaperony.
Vypučení
K vypučení je potřeba interakce kapsidových proteinů a obalových proteinů. Ukazuje se také, že je vyžadována i přítomnost cholesterolu. Kapsidový protein obsahuje hydrofobní kapsu, se kterou interaguje E2 skrze tyr-ala-leu motiv. Přesná funkce 6K proteinu není známa, protože jeho absence neovlivní pučení. Existují však domněnky, že tento jev se dá vysvětlit mutací v E2.
Efekt na hostitele
Zhruba 3 hodiny po infekci je kompletně zastavena buněčná proteosyntéza, zatímco virová stále pokračuje. Existují 4 teorie, které tento jev vysvětlují:
- rozdílné prostředí, jako například koncentrace draslíkových iontů, které upřednostní virovou translaci
- přímá soutěž o translační mašinérii
- inhibice buněčné translace kapsidovým proteinem
- inhibice buněčné translace nestrukturním proteinem
Výzkumy ukázaly, že strukturní proteiny nemají vliv na cytopatický efekt, ale mutace v nsP2, konkrétně Pro726 v serin, redukuje tento efekt. Také se ukázalo, že virová infekce podporuje dsRNA aktivovanou protein kinasu (PKR)-závislou a PKR-nezávislou cestu, čímž dochází k redukci buněčné translace. Mutace v nsp2 ukázaly, že zastavení buněčné transkripce a translace jsou vzdálené jevy a mají silný vliv na zmenšení produkce alfa a beta interferonů. Většina infekcí buněk obratlovců v kulturách končí apoptózou, což koreluje s neurovirulencí po smrti neuronů. Naproti tomu mozkytí buňky mohou projít různými příznaky, přes perzistenci až po nekrózu. Bylo popsáno množství proteinů, které se účastní antivirové obrany. Mozkytí buňky produkují malý hydrofobní peptid o 3,2 kD, který indukuje syntézu 55 kDa proteinu a inhibuje replikaci RNA viru. Krysí protein zinkového prstu (ZAP) byl původně označen jako inhibitor replikace retrovirů, avšak ukazuje se, že inhibuje celou řadu alphavirů navázáním na virovou mRNA. Přesný mechanismus není znám, předpokládá se však účast mnoha proteinů.
Epidemiologie
Alphaviry jsou rozšířené prakticky po celém světě. Většina z nich byla zavlečena člověkem, ovšem některé z nich jsou lidmi nepřenositelné. Příkladem jsou nemoci lososovitých. Bylo také zjištěno, že viry, které napadají mořské savce mohou být přenášené skrze vši. Tato schopnost infikovat velké množství organismů ukazuje na možnost identifikace ptačích a plazích alphavirů.
Odkazy
Literatura
- Fields Virology, 5. vydání
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu Alphavirus na Wikimedia Commons
Obrázky, zvuky či videa k tématu Alphavirus na Wikimedia Commons