Togaviridae
Čeleď Togaviridae zahrnuje jediný rod RNA virů – Alphavirus. Dříve byl k této čeledi řazen i rod Rubivirus, nyní řazený do čeledi Matonaviridae z řádu Hepelivirales a rod Arterivirus, nyní rozdělený do více rodů čeledi Arteviridae z řádu Nidovirales[1]; pouze viry rodu Alphavirus jsou přenášeny členovci. Gradient hustoty v sacharóze je 1,2 g/ml. Některé togaviry vykazují hemaglutinační aktivitu v závislosti na pH.
 | |
| Baltimorova klasifikace virů | |
| Skupina | IV (ssRNA viry s pozitivní polaritou) |
| Vědecká klasifikace | |
| Realm | Riboviria |
| Říše | Orthornavirae |
| Kmen | Kitrinoviricota |
| Třída | Alsuviricetes |
| Řád | Martellivirales |
| Čeleď | Togaviridae |
| Rod | |
| Některá data mohou pocházet z datové položky. | |
Taxonomie
Čeleď Togaviridae (podle římského oděvu, tógy), zahrnovala skupinu A, Alphaviry, a skupinu B, Flaviviry. Rody Rubivirus a Pestivirus byly poté přidány po prozkoumání umístění nestrukturních a strukturních genů na genomu (viz dále). I když jsou genomy a viriony obou rodů stejné, jejich replikační strategie jsou dostatečně odlišné, aby se dalo pochybovat o jejich společném původu.
Virologie
Morfologie
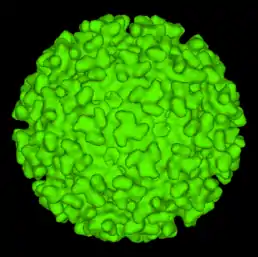
Virus se řadí mezi obalené viry (má membránovou schránku), přičemž se na membráně (o průměru 65-70 nm) vyskytuje 80 špic složených z trojic heterodimerů glykoproteinů E1 a E2, společně s menším proteinem 6K. Membrána je bohatá na cholesterol a sfingolipid, což jsou složky nezbytné pro vstup a výstup z buňky. Kapsid je sférický (ikosahedrický) o průměru 35-40 nm a skládá se z 240 podjednotek, (T=4). Molekulová váha je 52×10^6 d a hustota je 1,22 gramů na čtverečný centimetr.
Genom
Genom tvoří jedna molekula jednovláknové RNA s pozitivní polaritou o délce 9,7 (rubivirus) nebo 11,8 (alphavirus) kb a o mol.hm. 4 x 106. Nestrukturní proteiny (replikáza) jsou umístěné na 5' konci, zaujímající zhruba 2/3 délky genomu, zatímco strukturní (kapsidový protein a 3 obalové proteiny) jsou v poslední třetině, na 3' konci. Obě tyto skupiny jsou překládány jako polyprotein, který je následně rozštěpen. Genom je očepičkován a polyadenylován. nestrukturní proteiny jsou překládány rovnou z genomu, zatímco strukturní proteiny jsou překládány z subgenomické mRNA. Na začátku genomu je průměrně 59 nukleotidů dlouhá nepřekládaná oblast, zatímco na konci je dlouhá průměrně 322 nukleotidů.
Proteom
Proteom togavirů se seskládá z několika proteinů, které jsou translatovány v nestrukturním polyproteinu a v strukturním polyproteinu. Oba proteiny jsou schopny se samy rozštěpit a uvolnit tak funkční proteiny.
nsP1
Tento protein je součástí nestrukturního polyproteinu P123(4). Skládá se z 540 aminokyselinových zbytků a jako jediný je v asociaci s membránou, skrze palmytolilovaný cystein(420). Protein se však může vázat i hydrofobními zbytky aminokyselin 245-264. Má methyltransferázovou a guanylyltranferázovou aktivitu.
nsP2
Tento protein je součástí nestrukturního polyproteinu P123(4). Skládá se z 807 aminokyselinových zbytků. N koncová doména má helikázovou NTP a RNA-TP aktivitu, zatímco C koncová doména má papain-like serinovou proteázovou aktivitu a nefunkční methyltransferázovou aktivitu. Obsahuje také sekvenci, díky které je transportován do jádra, což vysvětluje fakt, že 50 % celkového množství tohoto proteinu je lokalizováno právě v jádře.
nsP3
Tento protein je součástí nestrukturního polyproteinu P123(4). Skládá se z 556 aminokyselinových zbytků. Funkce tohoto proteinu zůstávají sporné, avšak ukazuje se, že má roli v RNA syntéze. Zatímco N koncová doména je konzervativní napříč alphaviry, C koncová doména, obsahující asi 200 aminokyselinových zbytků, je bohatá na serinové a threoninové zbytky, které mohou být fosforylované, avšak význam této modifikace je stále nejasný. Protein má k membránám slabou afinitu, avšak asociuje s nimi v případě, že je exprimován v nepřítomnosti dalších nsP.
nsP4
Tento protein je součástí nestrukturního polyproteinu P123(4). Skládá se z 610 aminokyselinových zbytků. Jde o hlavní replikační protein. Jeho hladina je nižší, kvůli opal stop kodonu v polyproteinu P1234 mezi nsP3 a nsP4. Ukázalo se však, že odstranění tohoto stop kodonu spíše sníží replikaci virové RNA. I když je z větší části homologní s dalšími RdRP, N koncová doména obsahuje úsek, který nemá u jiných replikáz obdoby a slouží zřejmě jako vazebná doména pro ostatní nsP. N konec však také obsahuje tyrosinové zbytky, které zodpovídají za nestabilitu tohoto proteinu a za jeho degradaci. Protein se však stává relativně stabilním v komplexu s ostatními replikačními proteiny.
Capsid
Tento protein se skládá z 264 aminokyselinových zbytků a jeho úloha je obalení RNA a vytvoření nukleokapsidu. Prostupuje membránou a jeho 33 aminokyselin tvořících C koncovou doménu má strukturu dvou řeckých klíčů a aktivitu serionové proteázy. Jde o strukturní protein.
E3
Tento protein se skládá z 64 aminokyselinových zbytků a je součástí pE2 prekurzoru, jehož součástí je i E2. Jde o strukturní protein.
E2
Tento protein se skládá z 423 aminokyselinových zbytků a je součástí pE2 prekurzoru, jehož součástí je i E3. Je zodpovědný za navázání na receptor. Jeho doména tyr-ala-leu interaguje s hydrofobní kapsou nukleokapsidu. Jde o strukturní protein.
E1
Tento protein se skládá z 439 aminokyselinových zbytků a je zodpovědný za fúzi membrán. Membránou protein prostupuje, avšak dovnitř zasahují pouze 2 aminokyselinové zbytky. Obsahuje fúzní peptid, který je však v heterodimeru s E2 blokován. Tento heterodimer se však v nízkém pH rozpadne, čímž se peptid odkryje. Ten se pak zanoří do cílené membrány (obsahující cholesterol), načež dojde k trimerizaci E1 a masivní konformační změně tohoto glykoproteinu. Trimer má pak za následek deformaci membrány a její spojení s membránou endozomu, čímž vznikne pór, kterým nukleokapsid vnikne do buňky. E1 má také zjevně vlastnosti iontového kanálu, což je důležité pro replikaci. Jde o strukturní protein.
6K
Tento protein se skládá z 55 aminokyselinových zbytků. Zesiluje aktivitu jednotlivých částic. Jde o strukturní protein.
Životní cyklus
Vir do buňky vstupuje pomocí heparan sulfátu, avšak zdá se, že neslouží jako vstupní receptor, ale pouze jako „záchytný receptor“, díky kterému pak může dojít k interakci se vstupním receptorem. Některé výzkumy také naznačují, že se virulence zvýší, pokud je heparan sulfát z povrchu odstraněn, čímž se zabrání sekvestraci. Poté je indukována endocytóza, načež je translatována replikáza jako polyprotein, který je následně rozštěpen na nsP4 a P123, čímž se zajistí syntéza komplementárního (negativního) řetězce. Následně je z P123 rozštěpen nsP1, čímž se zajistí změna afinity k negativním řetězcům a syntéze pozitivního řetězce. Nakonec je zbytek polyproteinu znovu rozštěpen na nsP2 a nsP3, čímž se zaručí syntéza pozitivního řetězce a subgenomická mRNA. Následně pak dojde k translaci strukturních proteinů (kapsidový protein, E1-3). Virus poté následně vypučí z hostitelské buňky.
Epidemiologie
Alphaviry jsou rozšířené prakticky po celém světě. Většina z nich byla zavlečena člověkem, ovšem některé z nich jsou lidmi nepřenositelné. Příkladem jsou nemoci lososovitých. Bylo také zjištěno, že viry, které napadají mořské savce mohou být přenášené skrze vši. Tato schopnost infikovat velké množství organismů ukazuje na možnost identifikace ptačích a plazích alphavirů.
Odkazy
Literatura
- JURAJDA, Vladimír. Nemoci drůbeže a ptactva – virové infekce. 1. vyd. Brno: ES VFU Brno, 2002. 184 s. ISBN 80-7305-436-1. (anglicky)
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu Togaviridae na Wikimedia Commons
Obrázky, zvuky či videa k tématu Togaviridae na Wikimedia Commons