Chameleónovité
Chameleónovité (lat. Chamaeleonidae) je čeľaď infraradu leguány.
| chameleónovité | |||
 chameleón leopardí (Furcifer pardalis). | |||
| Vedecká klasifikácia | |||
|---|---|---|---|
| Vedecký názov | |||
| Chamaeleonidae Werner, 1902 | |||
| Vedecká klasifikácia prevažne podľa tohto článku | |||
Výskyt
Táto čeľaď sa pravdepodobne vyvinula vo Východnej Afrike, odkiaľ sa rozšírila okrem územia celej Afriky aj na Madagaskar, Kanárske ostrovy, na ostrov Maurícius, Zanzibar, Arabský polostrov, do Ázie a na juh Európy (hlavne Cyprus, Kréta, Malta, Sicília, Španielsko, Portugalsko), kde sa vyskytujú druhy Chamaeleo chamaeleon a Chamaeleo africanus.
Chameleóny obývajú všetky druhy tropických a horských dažďových pralesov, savany, vyskytujú sa aj na okrajoch polopúští a stepí. Väčšina druhov žije na stromoch a kroch, niektoré malé druhy prežívajú na zemi pod napadaným lístím.
Druhy
V čeľadi Chamaeleontidae je v súčasnosti asi 150 druhov chameleónov. Absolútne presný počet sa nedá určiť, nakoľko sú zverejňované nové revízie systematického členenia ich druhov a poddruhov. Jednotlivé druhy chameleónov môžu dosahovať rôznu veľkosť. Najmenším druhom je Brookesia micra, ktorý dosahuje dĺžku 30 mm[1], jedným z najväčších je Chameleón obrovský (Furcifer oustaleti) so svojou dĺžkou až 79 cm.[2] Priemerná dĺžka väčšiny druhov chameleónov je medzi 15 až 30 cm.
Chameleónovité jaštery väčšinou žijú v korunách stromov a kríkov. Pre tento spôsob života sú dobre prispôsobené. Chameleóny sa vyznačujú niektorými charakteristickými znakmi.
Telo chameleóna
Chameleóny majú bočne sploštené telo eliptického prierezu, dlhý chvost, ktorého schopnosť zvinutia sa do špirály umožňuje obkrútením okolo konára vytvoriť piaty pevný oporný bod pri držaní sa v korune stromov.
Nohy
Nohy chameleóna sú tzv. zygodaktilné – prsty sú zrastené do akýchsi klieští: na predných končatinách sú tri vonkajšie a dva vnútorné prsty a na zadných je to naopak. Na prstoch majú silné pazúry, ktoré umocňujú silu uchytenia počas lezenia po kôre stromov.
Hlava chameleóna
Chameleonóvité nemajú Jacobsonov orgán a podobne ako hadom im chýba vonkajšie aj stredné ucho. Toto nasvedčuje tomu, že by mohli byť chameleóny hluché, no dokážu medzi sebou komunikovať cez vibrácie pevných materiálov ako sú konáre stromov.
Oči

Oko chameleóna je považované za „technicky“ najdokonalejšie v celej živočíšnej ríši. Horná a dolná časť očných viečok chameleóna je zrastená kožou pokrytou jemnými farebnými šupinami tak, že je nich otvor vo veľkosti očnej zrenice. Oči tohto plaza sa môžu pohybovať samostatne a dokážu súčasne vnímať dva objekty. Zviera môže vidieť v rozsahu 360°. Chameleón vertikálne otáča okom o 90°, horizontálne ním otáča o 180°. Vo chvíli lokalizácie koristi sa dokážu chameleóny obidve oči sústrediť na tento jeden bod, čím chameleón získa ostrý, priestorový stereoskopický obraz, registrujú takto aj najmenší hmyz už do vzdialenosti 5 – 10 cm.
Chameleóny sú schopné vnímať aj ultrafialové svetlo.[3]
Jazyk
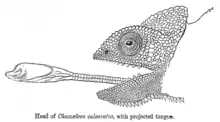
Chameleónovité jaštery majú veľmi dlhý jazyk, ktorý svojou dĺžkou môže presiahnuť dĺžku ich tela. Tento jazyk dokážu z tlamy vystreliť rýchlosťou väčšou, aká je postrehnuteľná ľudským okom.[4] Jeho koniec je silne lepkavý, s hruškovitým svalom, ktorý rýchlo udrie a aj s pomocou mierneho podtlaku uchopí korisť. Následne je korisť rýchlo vtiahnutá do jeho silnej tlamy, v ktorej ju rozdrví sa skonzumuje.
Farba chameleóna


Známa je schopnosť chameleóna meniť svoju farbu.
Chameleóny majú špeciálne kožné bunky, nazývané chromatofóry, ktoré sa nachádzajú pod ich priesvitnou vonkajšou kožou. Vo vrchných vrstvách sa nachádzajú bunky nazvané xanthofóry a erythrofóry, ktoré obsahujú žltý a červený pigment. Pod týmito bunkami sa nachádza vrstva, v ktorej sú bunky iridofóry (alebo aj guanofóry), obsahujúce transparentnú kryštalickú substanciu guanín. Cez túto substanciu sa vytvára modré sfarbenie, ktoré v kombinácii s vonkajšou žltou či červenou farbou vytvára výsledný efekt zelených či fialových odtieňov. Tmavé farbivo, melanín, ktoré reguluje tmavé farebné škály, sa nachádza pod týmito dvoma vrstvami.
Týmto spôsobom pri nervovom podráždení v jednotlivých chromatofóroch dochádza k zmenám viditeľnosti pigmentov ich posunom smerom k povrchu, alebo dovnútra kože. Princípom podobným, ako keď maliar mieša zrniečka farebných pigmentov na svojej palete, sa takto menia farebné odtiene kože chameleóna v závislosti od genetických dispozícií jednotlivých druhov. Farby chameleónov sa takto môžu meniť od zelenej, hnedej, čiernej, bielej, smotanovej, žltej až po modrú[5][6]
Základné sfarbenie chameleónov je, podobne ako u väčšiny živočíchov, kombinované tak, aby splynuli so svojím prostredím. Nie je pravda, že zmena farby chameleóna je podmienená aktuálnou adaptáciou na farbu prostredia. Na sfarbenie má vplyv aj teplota, ročné obdobie, intenzita osvetlenia, denná doba, ale aj jeho nálada, pohlavie a správanie v období pohlavnej aktivity, stres a iné vplyvy.[7] Zmena farby chameleóna je teda skôr jedným zo spôsobov komunikácie. Vyjadruje aj emócie a môže byť cestou, ktorá má jedinca počas dvorenia zatraktívniť u opačného pohlavia.[8] Rôzne druhy chameleónov používajú na zmenu svojho sfarbenia rozdielne farby a ich kombinácie.
- Rôzne sfarbenia druhu Furcifer pardalis
 Furcifer pardalis (samec)
Furcifer pardalis (samec) Furcifer pardalis v strese.
Furcifer pardalis v strese. Furcifer pardalis
Furcifer pardalis.jpg.webp) Furcifer pardalis detail.
Furcifer pardalis detail.
Pohlavný dimorfizmus

U väčšiny druhov chameleónov sa vyskytuje výrazný pohlavný dimorfizmus, samce v porovnaní so samicami sa vyznačujú väčšou pestrosťou a často u viacerých druhov chameleónov majú na sebe rôzne výraznejšie ozdoby, ako sú výrastky, goliere, rohy, alebo helma (napríklad aj chameleón jemenský, Chamaeleo calyptratus).
Reprodukcia

Chameleóny sú najčastejšie vajcorodé, niektoré sú vajcoživorodé.
Samičky vajcorodých druhov kladú vajíčka asi po 3 – 6 týždňoch gravidity. Zlezú zo stromov a v závislosti od druhu chameleóna vyhrabú 10 – 30 cm hlbokú jamu. Na dne tejto jamy sa počas kladenia otáčajú, po nakladení jamu zahrabú a toto miesto opustia. Množstvo nakladených vajec tiež závisí od druhu, malé druhy rodu Brookesia môžu naklásť iba 2 – 4 vajcia, pokým Chamaeleo calyptratus dokáže naklásť až 80 – 100 vajec, ale aj toto množstvo môže byť u tohto druhu chameleóna variabilné. Doba do vyliahnutia mláďat je tiež variabilná a môže byť od 4 do 12 mesiacov. Inkubácia môže trvať aj dlhšie, z vajec druhu Calumma parsonii sa mláďatá vyliahnu až po 24 mesiacoch.
Vajcoživorodé chameleóny (napr. Chamaeleo jacksonii) majú obdobie gravidity okolo 5 – 6 mesiacov. Novonarodené chameleóny spia obalené v priesvitnom obale, kým sa nedotknú zeme či konára, a vtedy sa po prebudení pokúšajú vyslobodiť z membrány. Samice vajcoživorodých chameleónov dokážu mať naraz 8 – 30 živých mláďat.
Potrava
Zaujímavý je spôsob lovu chameleónovitých jašterov. Hlavnou potravinovou zložkou chameleónov je rôzny hmyz, hlavne kobylky (Acrididae), modlivky (Mantodea), svrčky (Gryllidae), na ktorý si dokážu počkať v stave nehybného postoja na konári. V prípade, že sa ich korisť usadí v dosahu ich dlhého lepkavého jazyka, vymrštia ho a úctyhodnou rýchlosťou si chytený hmyz týmto jazykom vtiahnu do papule. Väčšie druhy sa môžu živiť aj menšími vtákmi, alebo inými menšími zástupcami radu jašterotvarých.
Druhy ako sú Chamaeleo calyptratus a Chamaeleo jacksonii majú ako doplnok stravy aj časti niektorých rastlín.
Chameleóny preferujú tečúcu, dažďovú vodu pred stálymi vodnými zdrojmi, nie je preto jednoduché ich naučiť, aby ju v zajatí konzumovali z misky.
Systematika
Rody z podčeľade chameleóny sa po slovensky volajú chameleón.
Podčeľaď Chamaeleoninae - chameleóny




.jpg.webp)







- Rod: Bradypodion Fitzinger, 1843
- Bradypodion caffer (Boettger, 1889)
- Bradypodion damaranum (Boulenger, 1887)
- Bradypodion dracomontanum Raw, 1976
- Bradypodion gutturale (Smith, 1849)
- Bradypodion kentanicum (Hewitt, 1935)
- Bradypodion melanocephalum (Gray, 1865)
- Bradypodion nemorale Raw, 1978
- Bradypodion occidentale (Hewitt, 1935)
- Bradypodion pumilum (Gmelin, 1789)
- Bradypodion setaroi Raw, 1976
- Bradypodion taeniabronchum (Smith, 1831)
- Bradypodion thamnobates Raw, 1976
- Bradypodion transvaalense (FitzSimons, 1930)
- Bradypodion ventrale (Gray, 1845)
- Rod: Calumma Gray, 1865
- Calumma amber Raxworthy & Nussbaum 2006
- Calumma ambreense Ramanantsoa 1974
- Calumma andringitraensis (Brygoo, Blanc & Domergue 1972)
- Calumma boettgeri (Boulenger 1888)
- Calumma brevicorne (Günther 1879)
- Calumma capuroni (Brygoo, Blanc & Domergue 1972)
- Calumma cucullatum (Gray 1831)
- Calumma crypticum Raxworthy & Nussbaum 2006
- Calumma fallax (Mocquard 1900)
- Calumma furcifer (Vaillant & Grandidier 1880) Methuen & Hewitt 1913
- Calumma gallus (Günther 1877)
- Calumma gastrotaenia (Boulenger 1888)
- Calumma glawi Böhme 1997
- Calumma globifer (Günther 1879)
- Calumma guibei (Hillenius 1959)
- Calumma guillaumeti (Brygoo, Blanc & Domergue 1974)
- Calumma hilleniusi (Brygoo, Blanc & Domergue 1973)
- Calumma hafahafa Raxworthy & Nussbaum 2006
- Calumma jejy Raxworthy & Nussbaum 2006
- Calumma malthe (Günther 1879)
- Calumma marojezense Brygoo, Blanc & Domergue 1970
- Calumma nasutum (Duméril & Bibron 1836) Mertens 1933
- Calumma oshaughnessyi Günther 1881
- Calumma parsonii (Cuvier 1824)
- Calumma peyrierasi (Brygoo, Blanc & Domergue 1974)
- Calumma peltierorum Raxworthy & Nussbaum 2006
- Calumma tigris (Kuhl 1820)
- Calumma tsaratananense (Brygoo & Domergue 1968)
- Calumma tsycorne Raxworthy & Nussbaum 2006
- Calumma vatosoa Andreone, F. Mattioli, R. Jesu & J. E. Randrianirina 2001
- Calumma vencesi Andreone, F. Mattioli, R. Jesu & J. E. Randrianirina 2001
- Rod: Chamaeleo Laurenti, 1768
- Chamaeleo africanus Laurenti, 1768
- Chamaeleo anchietae Bocage, 1872
- Chamaeleo arabicus Matschie, 1893
- Chamaeleo calcaricarens Böhme, 1985
- Chamaeleo calyptratus Duméril & Bibron, 1851
- Chamaeleo chamaeleon (Linnaeus, 1758)
- Chamaeleo dilepis Leach, 1819
- Chamaeleo gracilis Hallowell, 1842
- Chamaeleo etiennei Schmidt, 1919
- Chamaeleo laevigatus Gray, 1863
- Chamaeleo monachus Gray, 1865
- Chamaeleo namaquensis Smith, 1831
- Chamaeleo quilensis Bocage, 1866
- Chamaeleo roperi Boulenger, 1890
- Chamaeleo ruspolii Boettger, 1893
- Chamaeleo senegalensis Daudin, 1802
- Chamaeleo zeylanicus Laurenti, 1768
- Rod: Trioceros Swainson, 1839
- Trioceros affinis Rüppell, 1845
- Trioceros balebicornutus Tilbury, 1998
- Trioceros bitaeniatus Fischer, 1884
- Trioceros camerunensis Müller, 1909
- Trioceros chapini Witte, 1964
- Trioceros conirostratus Tilbury, 1998
- Trioceros cristatus Stutchbury, 1837
- Trioceros deremensis Matschie, 1892
- Trioceros eisentrauti Mertens, 1968
- Trioceros ellioti Günther, 1895
- Trioceros feae Boulenger, 1906
- Trioceros fuelleborni Tornier, 1900
- Trioceros goetzei Tornier, 1899
- Trioceros harennae Largen, 1995
- Trioceros hoehnelii Steindachner, 1891
- Trioceros incornutus Loveridge, 1932
- Trioceros ituriensis Schmidt, 1919
- Trioceros jacksonii Boulenger, 1896
- Trioceros johnstoni Boulenger, 1901
- Trioceros kinetensis Schmidt, 1943
- Trioceros laterispinis Loveridge, 1932
- Trioceros marsabitensis Tilbury, 1991
- Trioceros melleri (Gray, 1865)
- Trioceros montium Buchholz, 1874
- Trioceros narraioca Necas, Modry & Slapeta, 2003
- Trioceros pfefferi Tornier, 1900
- Trioceros quadricornis Tornier, 1899
- Trioceros rudis Boulenger, 1906
- Trioceros schoutedeni Laurent, 1952
- Trioceros schubotzi Sternfeld, 1912
- Trioceros sternfeldi Rand, 1963
- Trioceros tempeli Tornier, 1899
- Trioceros tremperi Necas, 1994
- Trioceros werneri Tornier, 1899
- Trioceros wiedersheimi Nieden, 1910
- Rod: Furcifer Fitzinger, 1843
- Furcifer angeli (Brygoo & Domergue 1968)
- Furcifer antimena (Grandidier 1872)
- Furcifer balteatus (Duméril & Bibron 1851)
- Furcifer belalandaensis (Brygoo & Domergue 1970)
- Furcifer bifidus (Brongniart 1800)
- Furcifer campani (Grandidier 1872)
- Furcifer cephalolepis (Günther 1880) (Komoren)
- Furcifer labordi (Grandidier 1872)
- Furcifer lateralis (Gray 1831))
- Furcifer minor (Günther 1879)
- Furcifer monoceras (Boettger, 1913)
- Furcifer nicosiai (Jesu, Mattioli & Schimmenti 1999)
- Furcifer oustaleti (Mocquard 1894)
- Furcifer pardalis (Curvier 1829))
- Furcifer petteri (Brygoo & Domergue 1966)
- Furcifer polleni (Peters 1873) (Komoren)
- Furcifer rhinoceratus (Gray 1843)
- Furcifer sp. "Montagne d'Ambre"
- Furcifer tuzetae (Brygoo, Bourgat & Domergue 1972)
- Furcifer verrucosus (Cuvier 1829)
- Furcifer willsii (Günther 1890)
- Rod: Kinyongia Tilbury, Tolley & Branch, 2006
- Kinyongia adolfifriderici (Sternfeld, 1912)
- Kinyongia carpenteri (Parker, 1929)
- Kinyongia excubitor (Barbour, 1911)
- Kinyongia fischeri (Reichenow, 1887)
- Kinyongia oxyrhina Klaver & Böhme, 1988
- Zweihornchamäleon (Kinyongia tavetana)
- Kinyongia tenuis (Matschie, 1892)
- Kinyongia uthmoelleri (Müller, 1938)
- Kinyongia xenorhina (Boulenger, 1901)
- Rod: Nadzikambia Tilbury, Tolley & Branch, 2006
- Nadzikambia mlanjensis (Broadley, 1965)
Podčeľaď Brookesiinae - brukesie


- Rod: Brookesia Gray, 1865
- Brookesia ambreensis (Raxworthy & Nussbaum, 1995)
- Brookesia antakarana (Raxworthy & Nussbaum, 1995)
- Brookesia bekolosy (Raxworthy & Nussbaum, 1995)
- Brookesia betschi (Brygoo, Blanc & Domergue, 1974)
- Brookesia bonsi (Ramanantsoa, 1980)
- Brookesia brygooi (Raxworthy & Nussbaum, 1995)
- Brookesia decaryi (Angel, 1939)
- Brookesia dentata (Mocquard, 1900)
- Brookesia ebenaui (Boettger, 1880)
- Brookesia exarmata (Schimmenti & Jesu, 1996)
- Brookesia griveaudi (Brygoo, Blanc & Domergue, 1974)
- Brookesia karchei (Brygoo, Blanc & Domergue, 1970)
- Brookesia lambertoni (Brygoo & Domergue, 1970)
- Brookesia lineata (Raxworthy & Nussbaum, 1995)
- Brookesia lolontany (Raxworthy & Nussbaum, 1995)
- Brookesia minima (Boettger, 1893)
- Brookesia micra (Glaw et al., 2012)
- Brookesia nasus (Boulenger, 1887)
- Brookesia perarmata (Angel, 1933)
- Brookesia peyrierasi (Brygoo & Domergue, 1974)
- Brookesia stumpffi (Boettger, 1894)
- Brookesia superciliaris (Kuhl, 1820)
- Brookesia therezieni (Brygoo & Domergue, 1970)
- Brookesia thieli (Brygoo & Domergue, 1969)
- Brookesia tuberculata (Mocquard, 1894)
- Brookesia vadoni (Brygoo & Domergue, 1968)
- Brookesia valerieae (Raxworthy, 1991)
- Rod: Rhampholeon Günther, 1874
- Podrod: Bicuspis Loveridge, 1956
- Rhampholeon marshalli Boulenger, 1906
- Rhampholeon gorongosae Broadley, 1971
- Podrod: Rhampholeon Günther, 1874
- Rhampholeon spectrum (Buchholz, 1874)
- Rhampholeon temporalis (Matschie, 1892)
- Rhampholeon viridis Mariaux & Tilbury, 2006
- Podrod: Rhinodigitum Matthee, Tilbury & Townsend, 2004
- Rhampholeon acuminatus Mariaux & Tilbury, 2006
- Rhampholeon beraduccii Mariaux & Tilbury, 2006
- Rhampholeon boulengeri Steindachner, 1911
- Rhampholeon chapmanorum Tilbury, 1992
- Rhampholeon moyeri Menegon, Salvidio & Tilbury, 2002
- Rhampholeon nchisiensis (Loveridge, 1953)
- Rhampholeon platyceps Güntherc, 1893
- Rhampholeon spinosus (Matschie, 1892)
- Rhampholeon uluguruensis Tilbury & Emmrich, 1996
- Podrod: Bicuspis Loveridge, 1956
- Rod: Rieppeleon Matthee, Tilbury & Townsend, 2004
- Rieppeleon brachyurus (Günther, 1893)
- Rieppeleon brevicaudatus (Matschie, 1892)
- Rieppeleon kerstenii (Peters, 1868)
Referencie
- Rivaling the World's Smallest Reptiles: Discovery of Miniaturized and Microendemic New Species of Leaf Chameleons (Brookesia) from Northern Madagascar
- GLAW, Frank, Vences, Miguel A Field Guide to Amphibians and Reptiles of Madagascar. 2nd edition. vyd. Köln : M. Vences & F. Glaw Verlags GbR, 1994. ISBN 3-929449-01-3.
- Chameleon News, August 2004
- A Lethal Lashing Tongue
- National Geographic. May 2007. P. 10.
- National Geographic Explorer (Student Magazine) - Featured Article
- HARRIS, Tom. How Animal Camouflage Works [online]. How Stuff Works, [cit. 2006-11-13]. Dostupné online.
Iné projekty
 Commons ponúka multimediálne súbory na tému Chameleónovité
Commons ponúka multimediálne súbory na tému Chameleónovité Wikidruhy ponúkajú informácie na tému Chameleónovité
Wikidruhy ponúkajú informácie na tému Chameleónovité
Zdroje
- Tento článok je čiastočný alebo úplný preklad článkov Chameleon na anglickej Wikipédii a na neurčenej Wikipédii (číslo revízie nebolo určené).
- Petr Nečas: Chameleoni