Trichobilharzia szidati
Trichobilharzia szidati je ptačí motolice z čeledi Schistosomatidae střídající v rámci svého životního cyklu 2 hostitele – definitivního hostitele (ptáci z čeledi kachnovití) a mezihostitele (plži plovatka bahenní a Stagnicola palustris).[2] Oproti příbuzným lidským krevničkám se T. szidati vyskytuje i v mírnějších evropských podmínkách, kde je společně s celým rodem Trichobilharzia označována za nejčastějšího původce kožního onemocnění tzv. cerkáriové dermatitidy u lidí.[3][4][5] Jejich cerkárie totiž dokáží pronikat i do pokožky savců (vč. člověka)[5], ve kterých však nejsou schopny dokončit svůj cyklus a umírají během penetrace pokožky či případné následné migraci organismem.[6]
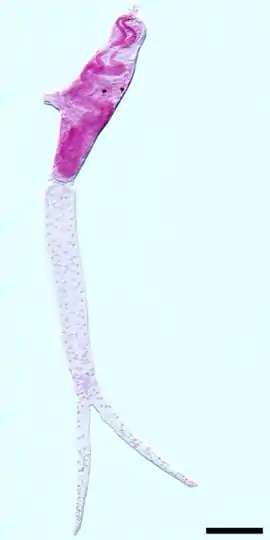 Trichobilharzia szidati, cerkárie | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | ploštěnci (Platyhelminthes) |
| Třída | motolice (Trematoda) |
| Podtřída | Digenea |
| Čeleď | Schistosomatidae |
| Rod | Trichobilharzia |
| Binomické jméno | |
| Trichobilharzia szidati Neuhaus, 1952 [1] | |
| Některá data mohou pocházet z datové položky. | |
Druhová determinace
Determinace a popis druhu Trichobilharzia szidati (resp. ocelátních furkocerkárií uvolňovaných z evropských sladkovodních plžů) je nejasný. S prvním záznamem se setkáváme v roce 1855, kdy La Valette označil cerkárie vylučované z plže Lymnaea stagnalis jako Cercaria ocellata, klasifikovanou v roce 1931 Brumptem za larvální stádium druhu Trichobilharzia ocellata. Ve 30. a 40. letech 20. století přibývalo pozorování výskytu „C. ocellata“, avšak tyto cerkárie se lišily rozměry i mezihostitelskými plži, což vyústilo v popis několika nových „druhů“ cerkárií. V roce 1952 se Neuhausovi podařilo popsat kompletní životní cyklus jednoho z nich, který pojmenoval T. szidati. I přes to bylo nadále často používáno jméno T. ocellata, i když se ukázalo, že jde o komplex, ze kterého se postupně vydělilo několik dalších druhů (T. anseri, T. franki,T. mergi, T. regenti, T. salmanticensis). Rudolfová a kol. (2005) prokázali pomocí molekulárních metod identitu evropských izolátů ocelátních furkocerkárií uvolněných z L. stagnalis chovaných v evropských parazitologických laboratořích (Praha, Amsterdam, Erlangen) i odlovených ve volné přírodě (ČR, Polsko). Na základě toho bylo navrženo označovat tyto druhy jednotně jako T. szidati a jméno T. ocellata dále nepoužívat.[7]
Životní cyklus
Dospělí jedinci jsou v definitivních hostitelích nejčastěji lokalizováni ve střevní tkáni či blízkých cévách. Při spárování dospělců jsou samičky dočasně usídleny v canalis gyneacophorus samců a jsou jimi oplozovány. Produkovaná vajíčka s již plně vyvinutými miracidii opouštějí hostitele společně s trusem.
Krátce po kontaktu vajíček s vodou dochází k uvolnění prvních larválních stádii – miracidií, která ve vodním prostředí aktivně vyhledávají své mezihostitele. Orientaci ve vodním prostředí, resp. vyhledání mezihostitelů, jim usnadňují látky, které plži vylučují (MAGs = miracidia attracting glycoproteins)[8], a schopnost pozitivní či negativní fototaxe a geotaxe. Přes tělní povrch plžů miracidia penetrují pomocí proteolytických enzymů uvolňovaných z apikální žlázy v přední části těla. Penetrace je doprovázena svlékáním povrchových ciliárních buněk a vznikem syncytiálního tegumentu (neodermis) na povrchu těla parazitů. Z miracidií se tak stávají mateřské sporocysty, z jejichž zárodečných buněk následně vznikají další generace dceřiných sporocyst.
Po uplynutí prepatentního období se uvnitř plže začnou vytvářet generace cerkárií, které opouští tělo plže zpátky do vody. Ve vodě se pohybují pomocí rozdvojeného ocásku a vyhledávají definitivního hostitele. Po přichycení k pokožce cerkárie uvolňují obsah penetračních žláz s histolytickými enzymy, díky kterým jsou schopny proniknout do kůže hostitele.[9] Při penetraci pokožky navíc cerkárie odhazují ocásky, čímž se mění na poslední larvální stádia tzv. schistosomula, která poté migrují prostřednictvím krevního řečiště do oblasti střev, kde se již jako pohlavně vyzrálé motolice rozmnožují.
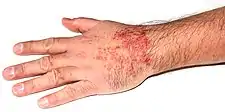
Vlivem podobného složení chemických látek na pokožce teplokrevných obratlovců (ptáci a savci) mohou cerkárie ptačích motolic penetrovat i do savčích hostitelů (vč. člověka), kteří v jejich životním cyklu představují tzv. náhodné hostitele. V nich jsou parazité imunitním systém zlikvidováni již při penetraci pokožky či případné následné migraci organismem, a tak svůj vývoj v těchto hostitelích nejsou schopny dokončit. Infekce náhodných hostitelů je charakteristická vznikem kožní vyrážky – cerkáriové dermatitidy.[10][11]
Migrace a patogenita v definitivním hostiteli
Po průniku do definitivního hostitele se cerkárie T. szidati mění na poslední larvální stádia – schistosomula. Kvůli potřebě vyhnout se imunitnímu systému hostitele je tato přeměna doprovázena shozením imunogenního povrchového glykokalyxu[12] a zdvojením cytoplazmatické membrány na povrchu těla parazita.[13] Aby mohla schistosomula vstoupit do krevního řečiště hostitele, musí nejdříve prostoupit několika vrstvami kůže, kde se orientují pomocí negativní fototaxe.[14] Kromě toho bylo u schistosomul T. szidati in vitro také popsáno pozitivně chemotaktické chování k určitým koncentracím L-argininu a D-glukóze. Má se za to, že tímto způsobem schistosomula dokáží určit svou polohu v kůži hostitele a lokalizovat blízké cévy.[15] Jakmile schistosomula opustí kůži (19 hodin po infekci), pronikají do oběhové soustavy hostitele, odkud migrují do srdce a plic.[16] Dospělí jedinci jsou nejčastěji lokalizování ve tkáních střev či blízkých cévách.[17]
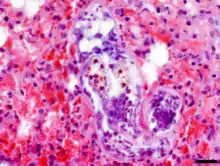
Penetrace kůže definitivního hostitele je doprovázena zarudnutím v místě průniku cerkárií či rozvojem petechií. Následná migrace a kumulace schistosomul v plicích může být doprovázena vznikem hemoragií, edému a zánětlivých uzlíků v plicní tkáni tvořených lymfocyty, heterofily, granulocyty a makrofágy.[16] U příbuzného druhu Anserobilharzia brantae bylo popsáno, že při uvolňování oplozených vajíček může dojít k jejich zanesení do tkáně jater nebo střev (podobně jako u lidských schistosom), kde mohou spustit zánětlivou imunitní reakci;[18] pro T. szidati však tato data k dispozici nejsou.
Migrace a patogenita v náhodném hostiteli
.png.webp)
Při experimentálních infekcích náhodných hostitelů jsou cerkárie schopny transformace na schistosomula, a dokonce i částečné migrace hostitelem.[19][20] Vývoj a migrace schistosomul byly prokázány u myší, křečků, morčat, králíků i makaků rhesus.[20][19] V laboratorních myších se nejvíce schistosomul koncentruje v plicích, avšak larvy obsazují i další orgány (srdce, játra, ledviny, střeva). K průniku přes pokožku a migraci hostitelem dochází především při primárních infekcích, kdy imunitní systém hostitele reaguje méně agresivně, nežli při reinfekcích. Opakované infekce u náhodných hostitelů, včetně lidí, vyvolávají mnohem silnější projevy cerkáriové dermatitidy[5] (kožní vyrážka) zapříčiněné sekrecí histaminu žírnými buňkami v kůži.[21]
Infikované myši i ostatní zvířata (křečci, králíci, makakové rhesus) mají na povrchu plic četné hemoragie.[16][20] Při histopatologickém vyšetření je pozorováno překrvení a edémy plicní tkáně a 4. den po infekci také shlukování lymfocytů v okrajových částech plicních laloků.[16] Schistosomula T. szidati se v plicích myší nalézají nejdříve 10 h po infekci[19] a nejpozději 10. den po infekci.[16] S přihlédnutím na nepoškozený tegument je za jejich úhyn zodpovědný buď neznámý imunologický faktor, nebo nedostatek esenciálních nutričních látek.[22]
Odkazy
Reference
- NEUHAUS, W. Biologie und Entwicklung yon Trichobilharzia szidati n. sp. (Trematoda, Schistosomatidae), einem Erreger von Dermatitis beim Menschen. Zeitschrift für Parasitenkunde, 1952, 15, 203–266.
- KOCK, S. Investigations of intermediate host specificity help to elucidate the taxonomic status of Trichobilharzia ocellata (Digenea: Schistosomatidae). Parasitology, 2001, 123, 67–70.
- SOLDÁNOVÁ, M., SELBACH, C., KALBE, M., KOSTADINOVA, A., SURES, B. Swimmer's itch: Etiology, impact, and risk factors in Europe. Trends in Parasitol. 2013, 29, 65–74.
- HORÁK, P., MIKEŠ, L., LICHTENBERGOVÁ, L., SKÁLA, V., SOLDÁNOVÁ, M., BRANT, S.V. Avian schistosomes and outbreaks of cercarial dermatitis. Clin. Microbiol. Rev., 2015, 28, 165–190.
- MACHÁČEK, Tomáš; TURJANICOVÁ, Libuše; BULANTOVÁ, Jana. Cercarial dermatitis: a systematic follow-up study of human cases with implications for diagnostics. Parasitology Research. 2018-12, roč. 117, čís. 12, s. 3881–3895. PMID: 30302587. Dostupné online [cit. 2019-05-14]. ISSN 1432-1955. DOI 10.1007/s00436-018-6095-0. PMID 30302587.
- HORÁK, P., KOLÁŘOVÁ, L., ADEMA, C.M. Biology of the schistosome genus Trichobilharzia. Adv. Parasitol., 2002, 52, 155–233.
- RUDOLFOVÁ, J., HAMPL, V., BAYSSADE-DUFOUR, C., LOCKYER, A.E., LITTLEWOOD, D.T., HORÁK, P. Validity reassessment of Trichobilharzia species using Lymnaea stagnalis as the intermediate host. Parasitol. Res., 2005, 95, 79–89.
- KALBE, M., HABERL, B., HAAS, W. Snail host finding by Fasciola hepatica and Trichobilharzia ocellata: compound analysis of “miracidia-attracting glycoproteins”. Exp. Parasitol., 2000, 96, 231–242.
- MIKEŠ, L., ZÍDKOVÁ, L., KAŠNÝ, M., DVOŘÁK, J., HORÁK, P. In vitro stimulation of penetration gland emptying by Trichobilharzia szidati and T. regenti (Schistosomatidae) cercariae. Quantitative collection and partial characterization of the products. Parasitol. Res., 2005, 96, 230–241.
- KOLÁŘOVÁ, L., HORÁK, P., SKÍRNISSON, K., MAREČKOVÁ, H., DOENHOFF, M. Cercarial dermatitis, a neglected allergic disease. Clinic Rev. Allerg. Immunol., 2013, 45, 63–74.
- KOLÁŘOVÁ, L. Cerkáriová dermatitida stále aktuální – spektrum patogenních agens a jejich vývoj v nespecifickém hostiteli. Remedia - Klinická mikrobiologie. 1999, 3, 260–267.
- HORÁK, P. Developmentally regulated expression of surface carbo- hydrate residues on larval stages of the avian schistosome Trichobilharzia szidati. Folia Parasitol, 1995 42, 255–265.
- HORÁK, P. KOVÁŘ, L. KOLÁŘOVÁ, L. NEBESÁŘOVÁ, J. Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and its putative immunological impact. Parasitology, 1998, 116, 139–147.
- GRABE, K., HAAS, W. Navigation within host tissues: cercariae orientate towards dark after penetration. Parasitol. Res., 2004a, 93, 111–113.
- GRABE, K., HAAS, W. Navigation within host tissues: Schistosoma mansoni and Trichobilharzia ocellata schistosomula respond to chemical gradients. Int. J. Parasitol., 2004b, 34, 927–934.
- CHANOVÁ, M., VUONG, S., HORÁK, P. Trichobilharzia szidati: the lung phase of migration within avian and mammalian hosts. Parasitol. Res., 2007, 100, 1243–1247.
- BOURNS, T., ELLIS, F.C., RAU, M.E. Migration and development of Trichobilharzia ocellata (Trematoda: Schistosomatidae) in its duck hosts. Can. J. Zool., 1973, 51, 1021–1030.
- WOJCINSKI, Z.W., BARKER, I.K., HUNTER, D.B., LUMSDEN, H. An outbreak of schistosomiasis in Atlantic brant geese, Branta bernicla hrota. J. Wild. Dis., 1987, 23, 248–255.
- HAAS, W., PIETSCH, U. Migration of Trichobilharzia ocellata schistosomula in the duck and in the abnormal murine host. Parasitol. Res., 1991, 77, 621–644.
- OLIVIER, L. Observations on the migration of avian schistosomes in mammals previously unexposed to cercariae. J. Parasitol., 1953, 39, 237–246.
- KOUŘILOVÁ, P., HOGG K.G., KOLÁŘOVÁ L., MOUNTFORD A.P. Cercarial dermatitis caused by bird schistosomes comprises both immediate and late phase cutaneous hypersensitivity reactions. J. Immunol. 2004, 172, 3766-3774.
- HORÁK, P., KOLÁŘOVÁ, L. Survival of bird schistosomes in mammalian lungs. Int. J. Parasitol., 2000, 30, 65–68.