Plastid
Plastid je eukaryotická semiautonomní organela přítomná v buňkách rostlin a některých dalších eukaryotických organismů (zejména různých řas). V typickém případě slouží k fotosyntéze a nazývají se chloroplasty. V mnohých případech však plastidy ztrácí tuto funkci a přizpůsobují se k funkcím jiným. Mohou pak sloužit jako zásobní organela nebo odpovídat za určité zabarvení buňky.
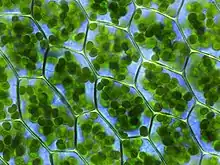
Je obalen nejméně dvěma membránami a uvnitř té vnitřní se nachází stroma s thylakoidy. Plastid je dále typický obsahem tzv. plastidové DNA (pDNA) a má tak důležitý podíl na mimojaderné dědičnosti.
Evoluce plastidů
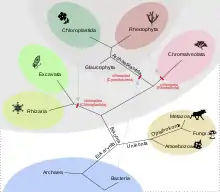
Podle tzv. endosymbiotické teorie se předpokládá, že jde o potomky bývalých endosymbiontů sinicového typu, v několika málo případech u konkrétních druhů řas však je známo, že jejich plastidy jsou potomky eukaryotických symbiontů typu řas (tzv. sekundární endosymbióza).
Vznikly pravděpodobně v evoluční historii až určitou dobu po vzniku samotné eukaryotické buňky a samotné eukaryogeneze se neúčastnily. Na rozdíl od mitochondrií se plastidy vyvinuly u několika, a to často nepříbuzných skupin eukaryot. Tzv. primární plastidy však pochází z sinice a vznikly pravděpodobně pouze jednou, a to u rostlin, které v širším pojetí zahrnují nejen zelené rostliny (Viridiplantae), ale i ruduchy (Rhodophyta) a glaukofyty (Glaucophyta).[1] Výjimkou jsou řasy rodu Paulinella, které získaly plastidy nedávno a nezávisle na rostlinách.[2]
Plastidy u jiných druhů fotosyntetizujících eukaryot vznikly především sekundární endosymbiózou, tedy pohlcením jedné ze skupin rostlin, případně endosymbiózou terciární.[3] Pohlcením ruduchy vznikly plastidy (někdy kvůli svému původu zvané rhodoplasty) např. u různých heterokont (Heterokonta) a rozsivek (Bacillariophyceae), pohlcením zelené řasy vznikly plastidy u Chlorarachniophyta, některých krásnooček (Euglenozoa) a jedné obrněnky.[4] Jako příklad prokázané terciární endosymbiózy lze uvést obrněnky rodu Kryptoperidinium, jejichž plastidy pocházejí z pohlcené rozsivky.[3]
Typy plastidů
Proplastidy
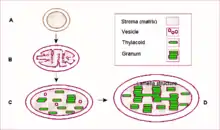

Proplastid je nezralý plastid. Lze ho nalézt v buňkách dělivých pletiv a v mladých buňkách. Během diferenciace a zrání buňky se mění v některý z dalších typů plastidů.
Chloroplasty
Fotosynteticky aktivní plastidy jsou takové, které obsahují fotosynteticky aktivní barviva a ve kterých tedy probíhá fotosyntéza, tj. zachycení sluneční energie a přeměna na organické látky.
Typickým příkladem je chloroplast, tzn. fotosynteticky aktivní zelený plastid obsahující chlorofyly, najdeme jej např. v buňkách zelených rostlin, proto mají zelenou barvu. Má 2 obalové membrány, vnitřní odškrcuje váčky tylakoidy, které tvoří třetí membránovou soustavu, jejíž části se nazývají grana (sloupcovité shluky mincovitých tylakoidních váčku) a lamely neboli stromální tylakoidy (propojující můstky).[5]
Někdy se odlišují na základě obsahu jistých fotosyntetických barviv či výskytu u určitých skupin organizmů i další typy fotosynteticky aktivních plastidů. Rodoplast (též rhodoplast) je fotosynteticky aktivní červený plastid obsahující fykoerytrin (červený) a fykocyanin (modrý). Lze jej nalézt třeba u ruduch (Rhodophyta). Feoplast je hnědý fotosynteticky aktivní plastid obsahující chlorofyl a fukoxantin, lze jej nalézt u hnědých řas (Phaeophyta).
Chromoplasty
Chromoplast je žlutý nebo červený plastid obsahující pouze pomocné fotosyntetické pigmenty (karotenoidy-karoteny, xantofyly), které nejsou schopny fotosyntetizovat. (Slouží pouze jako doprovodné sběrače fotonů pro chloroplasty.) Je to tedy bývalý chloroplast, který postrádá chlorofyl. Tvoří se především ve starších buňkách a to nejčastěji v plodech. Jeho úkolem je zbarvit povrch plodu nápadnou barvou aby přilákal konzumenty a umožnil tak šíření semen rostlin. Je také zodpovědný za podzimní barvu listí, neboť v něm je chlorofyl již rozložen.
Leukoplasty
Leukoplasty jsou plastidy, které neobsahují žádná barviva. Jejich úkolem je obvykle shromažďovat zásoby. Jsou známy tyto typy:
Genom plastidů
V plastidech je prokázána přítomnost kruhových molekul DNA, známých jako pDNA. U všech plastidových DNA se o pozůstatek prokaryotického genomu sinice, která byla kdysi v procesu eukaryogeneze pohlcena eukaryotem.[6] Navíc někdy vznikají plastidy vzniklé sekundární endosymbiózou, které však obsahují rovněž genom sinice. Zvláštností je u některých skupin tzv. nukleomorf, tedy zbytkový genom jiného eukaryota po sekundární endosymbiotické události.[7] Velikost pDNA je velice rozmanitá, redukované plastidové genomy se nachází zejména u druhů, které ztratily svou fotosyntetickou funkci.
Symbiotické chloroplasty

Za prvotní fázi nedokončeného procesu endosymbiotického vzniku druhotných plastidů lze považovat tzv. kleptoplastii, tedy zabudovávání živých plastidů získaných z potravy do buněk hostitelského, normálně heterotrofního organismu.
U některých mořských plžů živících se zelenými řasami bylo zjištěno, že do vlastních epitelových buněk zabudovávají chloroplasty z pozřených řas. Jedná se například o plže Elysia atroviridis, Elysiella pusilla, Elysia tuca, Bosellia mimetica, Oxynoe antillarum či Plakobranchus ocellatus.[8] Tyto plastidy přežívají až několik měsíců a zůstávají fotosynteticky aktivní. Nazývají se kleptoplastidy (řidčeji kleptoplasty).[9] Protože nejsou součástí zárodečného vývoje plžů, musí si je každá generace znovu získat z potravy.[pozn. 1]
V době potravního nedostatku jsou funkční kleptoplastidy důležité pro výživu plžů — ti přecházejí na mixotrofní, možná i autotrofní způsob výživy. Podíl na výživě za normálních podmínek, kdy je dostatek přirozené potravy, však prokázán nebyl.[8]
Kleptoplastidy byly zjištěny i u jiných hostitelských organismů.
U obrněnek Gymnodinium spp. a Pfisteria piscicida jsou fotosynteticky aktivní pouze několik dní, u Dinophysis spp. až 2 měsíce.[13]
Nálevník Myrionecta rubra získává kleptoplastidy z kryptomonády Geminigera cryophila.[14] U některých dírkonošců z rodů Bulimina, Elphidium, Haynesina, Nonion, Nonionella, Nonionellina, Reophax a Stainforthia, byly objeveny kleptoplastidy pocházející z obrněnek.
Tanozom
V r. 2013 byl objeven nový druh organely, odvozené od chloroplastu. Vyskytuje se v buňkách některých cévnatých rostlin. Probíhá v ní syntéza kondenzovaných taninů – polyfenolů, využívaných rostlinami k ochraně před býložravými škůdci a ultrafialovým záření. Podle toho dostala svůj název (tanin + lat. soma, tedy tělísko).[15][16]
Tanozomy se tvoří jako malé kulovité váčky (s velikostí kolem 30 nm), které se oddělují uvnitř chloroplastu z thylakoidů. Shluky tanozomů se pak dostávají ven z chloroplastu ve formě transportních váčků obalených membránou, vzniklou fúzí dvou obalových chloroplastových membrán. Transportní váčky přenášejí taniny v tanozomech do vakuoly. Celý váček se vchlípí do vakuoly a z tonoplastu tak získá další obalovou membránu. Taninové akrece ve vakuolách tak obsahují taniny uložené ve třech rozdílných membránách.[17]
Poznámky
- U plže Elysia chlorotica byl prokázán horizontální přenos více než 50 genů z chloroplastové DNA kleptoplastidů získaných z různobrvek druhu Vaucheria litorea do genomu plže a dokonce jejich translační aktivita. U vajíček a larv plže přitom přítomnost těchto genů v genomu zjištěna nebyla, což naznačuje, že genový přenos nastává u každé generace plžů až poté, co se začnou řasami živit.[10][11][12]
Reference
- Hedges S. B., Blair J. E., Venturi M. L., Shoe J. L. A molecular timescale of eukaryote evolution and the rise of complex multicellular life.. BMC Evol Biol. Jan 2004, čís. 28;4:2.. Dostupné online. PMID 15005799. (anglicky)
- NAKAYAMA, Takuro; ARCHIBALD, John M. Evolving a photosynthetic organelle. S. 1–3. BMC Biology [online]. 24. duben 2012. Svazek 10, čís. 35, s. 1–3. Dostupné online. PDF . ISSN 1741-7007. DOI 10.1186/1741-7007-10-35. PMID 22531210. (anglicky)
- ARCHIBALD, John M. The Puzzle of Plastid Evolution. Current Biology. 27. leden 2009, svazek 19, čís. 2, s. R81–R88. Dostupné v archivu pořízeném dne 2012-12-24. DOI 10.1016/j.cub.2008.11.067. (anglicky) Archivováno 24. 12. 2012 na Wayback Machine
- ČEPIČKA, Ivan; KOLÁŘ, Filip; SYNEK, Petr. Mutualismus, vzájemně prospěšná symbióza; Přípravný text – biologická olympiáda 2007–2008. Praha: NIDM ČR, 2007. S. 87.
- VOET, D.; VOETOVÁ, J. Biochemie. 1. čes. vyd. Praha: Victoria Publishing, 1995. ISBN 80-85605-44-9. S. 658–659.
- ALBERTS, Bruce , et al. The Molecular Biology of the Cell. [s.l.]: Garland Science, 2002. (4th. ed). Dostupné online. ISBN 0-8153-3218-1.
- ARCHIBALD, John M. Nucleomorph genomes: structure, function, origin and evolution. BioEssays: News and Reviews in Molecular, Cellular and Developmental Biology. 2007-04, roč. 29, čís. 4, s. 392–402. Dostupné online [cit. 2008-08-22]. ISSN 0265-9247. DOI 10.1002/bies.20551.
- MAEDA, Taro; HIROSE, Euichi; CHIKARAISHI, Yoshito, KAWATO, Masaru; TAKISHITA, Kiyotaka, et al. Algivore or Phototroph? Plakobranchus ocellatus (Gastropoda) Continuously Acquires Kleptoplasts and Nutrition from Multiple Algal Species in Nature. S. 1–12, e42024. PLoS ONE [online]. 25. červenec 2012. Svazek 7, čís. 7, s. 1–12. Dostupné online. PDF . DOI 10.1371/journal.pone.0042024. (anglicky)
- WAUGH, G. R.; CLARK, K. B. Seasonal and geographic variation in chlorophyll level of Elysia tuca (Ascoglossa: Opisthobranchia). Marine Biology. Září 1986, svazek 92, čís. 4, s. 483–487. Dostupné online. DOI 10.1007/BF00392508. (anglicky)[nedostupný zdroj]
- SCHWARTZ, Julie A.; CURTIS, Nicholas E.; PIERCE, Sidney K. FISH Labeling Reveals a Horizontally Transferred Algal (Vaucheria litorea) Nuclear Gene on a Sea Slug (Elysia chlorotica) Chromosome. S. 300–312. The Biological Bulletin [online]. 1. prosinec 2014. Svazek 227, čís. 3, s. 300–312. Dostupné online. ISSN 1939-8697. PMID 25572217. (anglicky)
- BHATTACHARYA, Debashish; PELLETREAU, Karen N.; PRICE, Dana C., SARVER, Kara E.; RUMPHO, Mary E. Genome Analysis of Elysia chlorotica Egg DNA Provides No Evidence for Horizontal Gene Transfer into the Germ Line of This Kleptoplastic Mollusc. S. 1843–1852. Molecular Biology and Evolution [online]. 2. květen 2013. Svazek 30, čís. 8, s. 1843–1852. Dostupné online. PDF . ISSN 1537-1719. DOI 10.1093/molbev/mst084. PMID 23645554. (anglicky)
- PAZDERA, Josef. „Eko-slimák“. O.S.E.L. [online]. 7. únor 2015. Dostupné online.
- MINNHAGEN, Susanna; CARVALHO, Wanderson F.; SALOMON, Paulo S., JANSON Sven. Chloroplast DNA content in Dinophysis (Dinophyceae) from different cell cycle stages is consistent with kleptoplasty. S. 2411–2417. Environmental Microbiology [online]. 2. červen 2008. Svazek 10, čís. 9, s. 2411–2417. Dostupné online. DOI 10.1111/j.1462-2920.2008.01666.x. (anglicky)
- JOHNSON, Matthew D.; OLDACH, David; DELWICHE, Charles, STOECKER Diane K. Retention of transcriptionally active cryptophyte nuclei by the ciliate Myrionecta rubra. S. 426–428. Nature [online]. 25. leden 2007. Svazek 445, čís. 7126, s. 426–428. Dostupné online. ISSN 1476-4687. DOI 10.1038/nature05496. (anglicky)
- GRENS, Kerry. New Organelle: The Tannosome. The Scientist [online]. 23. září 2013. Dostupné online. (anglicky)
- KUTÍK, Jaromír. Tanosomy. S. 115. Vesmír [online]. Únor 2014. Svazek 93, čís. 2014/2, s. 115. Dostupné online. ISSN 1214-4029.
- BRILLOUET, Jean-Marc; ROMIEU, Charles; SCHOEFS, Benoît, SOLYMOSI, Katalin; CHEYNIER, Véronique; FULCRAND, Hélène; VERDEIL, Jean-Luc; CONÉJÉRO, Geneviève. The tannosome is an organelle forming condensed tannins in the chlorophyllous organs of Tracheophyta. S. 1003–1014. Annals of Botany [online]. 11. září 2013. Svazek 112, čís. 6, s. 1003–1014. Dostupné online. pdf . ISSN 1095-8290. DOI 10.1093/aob/mct168. (anglicky)
Externí odkazy
 Obrázky, zvuky či videa k tématu plastid na Wikimedia Commons
Obrázky, zvuky či videa k tématu plastid na Wikimedia Commons  Slovníkové heslo plastid ve Wikislovníku
Slovníkové heslo plastid ve Wikislovníku