Auxiny
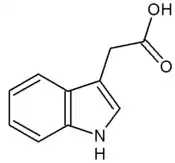
Auxiny jsou významná skupina rostlinných hormonů (neboli fytohormonů), které již v malých koncentracích ovlivňují dlouživý růst rostlinných buněk. Jejich působnost je však významnější, jsou nepostradatelné pro jakýkoli vývojový nebo tropický proces v rostlinách. Nejznámějším auxinem je kyselina indol-3-octová (IAA – z anglického indoleacetic acid).[1]
Historie
Název „auxin“ pochází z řeckého auxein což znamená dloužit se, prodlužovat. Auxin tento název získal díky své schopnosti působit zrychlování růstu ošetřených částí rostlin a zvláště prodlužování mladých částí stonků. První indicie o jeho existenci získal již Charles Darwin se svým synem Francisem okolo roku 1880. Společně prováděli série experimentů s klíčními rostlinami ovsa a chrastice a tyto výsledky zveřejnili ve své památné publikaci The Power of Movement in Plants (Pohyby v rostlinách). Zjistili, že když byla špička koleoptyle zakryta materiálem nepropustným pro světlo, tento orgán se neohýbal ke světelnému zdroji a nevykazoval tedy tzv. fototropickou reakci, tj. ohnutí ke světelnému zdroji. Identitu auxinu blíže nepopsali, ale předpověděli existenci signálu předávaného mezi pletivy, který tyto jevy ovlivňuje (klonili se spíše k mechanickému stimulu přenášenému mezi pletivy). Že se jedná patrně o malou molekulu, která těmito pletivy prochází, ukázal až roku 1913 dánský výzkumník Peter Boysen-Jensen (1883 - 1959).
Jasnější důkaz o této substanci (a také její jméno auxin) přinesl až roku 1926 holandský rostlinný fyziolog Frits Warmolt Went (1903 - 1990), který je také pokládán za oficiálního objevitele tohoto hormonu. Ten zjistil, že osvítíme-li opět špičku koleoptyle ovsa, pak ji uřízneme, položíme na agar a po nějaké době agarové bločky z takto mechanicky oddělených špiček přiložíme na jiné, neosvícené koleoptyle, jejichž špičky byly také odříznuty, budou vykazovat ohyb jako při fototropické reakci. Na základě těchto pozorování došel k závěru, že světlo vyvolává ve špičce produkci substance, která následně difunduje do agarového bločku a při přenesení na jinou koleoptyle vyvolá fototropickou reakci. Chemická struktura tohoto fytohormonu byla zjištěna v roce 1933 (z extraktů pocházejících z lidské moči) a až v roce 1946 byla IAA přímo popsána i v rostlinách.[2]
Zástupci
Za auxin můžeme označit řadu chemických látek, které mají společnou nebo obdobnou biologickou aktivitu, i když se chemicky liší. Nejvýznamnějším zástupcem je kyselina indol-3-octová, která je přírodním, tedy přirozeným auxinem, je v rostlině zřetelně nejvíce zastoupena a má nejvýznamnější účinek. Dalšími přirozenými auxiny jsou 4-chlor-indoloctová kyselina, fenyloctová kyselina (PAA, phenylacetic acid), indol-3-propionová kyselina (IPA, indole-3-propionic acid) a indol-3-máselná kyselina (IBA, indole-3-butyric acid). Poslední jmenovaná je dle všeho raději prekurzorem IAA než aktivním auxinem a někteří badatelé její přítomnost v některých modelových organizmech dokonce zpochybňují.
Vedle přirozených auxinů existují také auxiny syntetické, tedy připravované uměle. Zpravidla se jedná o chemicky odlišné látky, přesto se vyznačují účinky velmi podobným auxinům přirozeným. Významná je například kyselina 1-naftyloctová (NAA). Na rozdíl od IAA je chemicky stálejší (i levnější) a přidává se proto často jako účinný auxin do komerčních prostředků na podporu zakořeňování i jiných stimulátorů růstu. Dalším účinným v komerční sféře používaným syntetickým auxinem je 2,4-dichlorfenoxyoctová kyselina (2,4-D), která se často používá do některých desikantů a popř. herbicidů. Výhodou syntetických auxinů je, že ačkoli působí jako auxin, rostlina je neumí odbourávat a musí na ně tedy reagovat.
Syntéza a metabolizmus
Genetické důkazy získané na huseníčku (Arabidopsis thaliana) i dalších modelových organizmech ukázaly, že regulace auxinové syntézy je nepostradatelná pro utváření tělního plánu v rostlinách. Usuzuje se, že auxiny se syntetizují převážně z aminokyseliny tryptofanu (dráhy tryptofan-dependentní), neboť geny zapojené v těchto procesech jsou asociované s vážnými vývojovými defekty. Existují biosyntetické dráhy na tryptofanu nezávislé, avšak i tyto dráhy však sdílejí velkou část své syntézy s dráhou na tryptofanu závislou. Modernější důkazy podporující důležitost tryptofan-nezávislých drah však chybí.
Enzymy, které katalyzují syntézu tryptofanu, jsou lokalizovány v chloroplastech. Další kroky, vedoucí od tryptofanu k IAA, se ubírají se přes indol-3-pyruvát. Odehrávají se buď cytosolu nebo v endoplazmatickém retikulu. Právě manipulace s geny kódujícímy enzymy indol-3-pyruvátové dráhy ústí ve vývojové defekty, které se v mnoha rysech také shodují s poruchami spojovanými s defekty v genech pro syntézu tryptofanu. Rovněž byly popsány další dvě tryptofan-závislé dráhy: patrně cytoplazmatická, produkující IAA z indol-3-acetonitrilu a méně prozkoumaná z indol-3-acetamidu. Oproti indol-3-pyruvátové dráze se však nezdá, že by dvě později jmenované měly výrazný vliv na vývoj a morfogenezi u Arabidopsis. Jejich úloha snad bude v odpovědi na stimuly z okolí nebo při syntéze látek s auxinem souvisejícími jen nepřímo, ovšem i zde bude třeba některé indicie prověřit.
Auxiny se inaktivují reverzibilně (vratně) připojením aspartátu či glukózy. Spojuje se karboxyl auxinu s hydroxylem glukózy či aminem aspartátu. V případě spojení karboxylu a hydroxylu jde o esterifikaci. Inaktivace ireverzibilní (nevratná) se děje prostřednictvím oxidace uhlíku, který leží mezi dusíkem a zbytkem kyseliny octové, na karbonyl. Tento proces se patrně uskutečňuje v cytosolu a takto vzniká neaktivní oxindol-3-octová kyselina (OxIAA). Dle starších názorů dochází k degradaci také pomocí peroxidace, a to k nahrazení zbytku kyseliny octové methylenem na 3-metylenoxindol. V současném světle poznání však převládají argumenty, že by tento způsob degradace je biologicky nevýznamný, alespoň v běžných modelových organizmech.
Hormonální působení
Auxiny se účastní všech růstových regulací v rostlinném vývoji a pomáhají rostlině zprostředkovat vnější podněty a okolní svět, stejně jako pomáhají uskutečňovat zásadní kroky utváření tělního plánu. Přítomnost auxinu je zásadní pro jakýkoli vývojový proces v rostlinách a již v časných fázích určuje založení apikálního a bazálního pólu vznikajícího embrya.
Auxin není přítomen ve všech částech rostliny stejnou měrou. Dle klasických představ vzniká jeho převážná část v růstovém vrcholu prýtu. Odtud je do celého rostlinného těla transportován, a to až do kořenových špiček. Silné experimentální důkazy poukazují také na roli tzv. lokální auxinové biosyntézy, která ovlivňuje pouze buňky v místě takové syntézy nebo v jejím nejbližším okolí. Bylo zjištěno, že se auxin syntetizuje překvapivě i v kořenovém meristému a ovlivňuje tak jeho funkci. Jak jsou tyto dva principiálně odlišné procesy koordinovány, nebylo dosud uspokojivě vysvětleno.
Gradient auxinu vytvářený v růstovém vrcholu prýtu je zodpovědný za jeho apikální dominanci. Auxin při ní působí jako inhibitorrozvětvování tím, že opožďuje iniciaci laterálních pupenů (pupenů na boku stonku). Ve výsledku boční větve nepřerůstají hlavní vrchol. Při odstranění hlavního vrcholu přestane auxin proudit stonkem směrem dolů, laterální pupeny mohou vystoupit z inhibiční zóny a odejmutý vrchol přerůst. Znalost tohoto projevu je prospěšná při řezu stromů v zahradnické praxi.
Rovněž byla popsána zásadní role auxinu při založení místa formování budoucího pupenu na apikálním vrcholu (tzv. fylotaxe). Probíhá na základě pravidelného ustavování po genetické spirále podle tzv. zlatého úhlu. V prýtu auxin podporuje dlouživý růst. V kořeni jej auxin naopak inhibuje, avšak působí stimulačně na zakládání nových kořenů postranních. Z tohoto důvodu je nepostradatelnou součástí všech přípravků používaných k podpoře zakořeňování rostlinných stonkových řízků zvláště, a při vegetativním rozmnožování rostlin obecně. Auxin se tak účastní jako hlavní signální molekula komplexní regulace rozmístění i směru růstu kořenů a hlavních a postranních větví v prostoru. Tímto způsobem se rostlina snaží zefektivnit rozložení postranních orgánů, aby tím zlepšila svou pozici vůči konkurenčním rostlinám a urychlila svůj vývoj.
Část auxinu projde vodivými pletivy až do kořenové špičky, kde působí jako hlavní organizační prvek při ustavování dalšího směru růstu. Kořenová špička přerozděluje toky auxinu pomocí proteinu PIN do rostoucích částí kořene v závislosti na gravitaci, kterou je schopna vnímat díky zrnům škrobu (statolitům) umístěným ve specializovaných buňkách statocytech v oblasti kolumely kořenové čepičky. Různě přerozdělený auxin odpovídajícím způsobem ovlivňuje rychlost růstu části kořene, do které se dostal, a působí tak na určení směru růstu. Růstová reakce na gravitační podnět se nazývá pozitivní gravitropismus (tedy ve směru zemské tíže). V prýtu je auxin zodpovědný analogickým mechanismem za gravitropismus negativní (tedy ve směru od zemské tíže).
Transport
Transportují se pouze aktivní auxiny (tedy dle všeho nikoli jejich prekurzory, konjugáty nebo metabolické produkty). Na dlouhé vzdálenosti putuje auxin pasivně floémem spolu s asimiláty. Na krátké vzdálenosti se transportuje do buněk volným přechodem přes plazmatickou membránu anebo aktivně pomocí vtokových přenašečů rodiny AUX1/LAX. Jeho výtok je pak uskutečněn pomocí výtokových přenašečů z proteinových rodin PIN, či případně ABCB. Tyto přenašeče translokují disociovaný auxin přes membránu, buněčnou stěnou putuje sám difuzí. Nedisociovaný auxin dokáže procházet sám přes buněčnou stěnu i přes membránu.
AUX1 transportují disociovaný auxin symportem s jedním nebo dvěma protony a tedy bez přímé spotřeby ATP, PIN zřejmě antiportem (výměnou za proton) a ABCB na účet ATP. Protony, jejichž koncentrace v buňce se tímto neodvratně zvyšuje, jsou opět transportovány ven ATPázami na účet ATP.
Umístění zejména PIN v membráně je možné rychle přizpůsobit aktuálním podmínkám, což zajišťuje možnost rychlé změny směru a polarity toku auxinu.
Molekulární mechanismus působení auxinu
Auxin aktivuje přímo v jádře receptor TIR1, který přiměje komplex ubikvitinové ligázy E3 (tzv. SCFTIR1 komplex) k navázání ubikvitinu na AUX/IAA. Komplex AUX/IAA je inhibiční komplex transkripčních regulátorů AUX/IAA, který dosud okupoval promotor časných auxinem indukovaných genů a bránil jim v jejich transkripci. AUX/IAA s navázaným ubikvitinem je tak odsouzen k degradaci v proteasomu 26S, čímž se uvolní promotor časných auxinem indukovaných genů a tímto se spustí jejich transkripce. Takto je jsou aktivovány mj. transkripční faktory ARF, které se vážou na promotory pozdních auxinem indukovaných genů a iniciují tím jejich transkripci. Produktem genové exprese pozdních auxinem indukovaných genů jsou už proteiny, které přímo způsobují dloužení buněk, gravitropní odpověď či jiné, pro auxin typické reakce.
Byly také popsány účinky signální dráhy spouštěné proteinem ABP1 (auxin binding protein 1), objeveným již v sedmdesátých letech v kukuřici. Nověji publikovaná data ukazující zásadní důležitost tohoto proteinu v Arabidopsis však později nebyla potvrzena v současnosti procházejí revizí.
Odkazy
Reference
- Malý encyklopedický slovník. Praha: Academia, 1972, s. 74.
- PROCHÁZKA; ŠEBÁNEK, Jiří. Regulátory rostlinného růstu. Praha: Academia, 1997. 395 s. ISBN 80-200-0597-8.
Externí odkazy
 Obrázky, zvuky či videa k tématu auxiny na Wikimedia Commons
Obrázky, zvuky či videa k tématu auxiny na Wikimedia Commons - J. Friml - Auxin – univerzální vývojový signál v životě rostlin - časopis Živa 1/2007